
全缘千里光的染色体核型分析及其系统植物学特征
2020-08-12 09:00左青青肖佩林
遵义医科大学学报 2020年3期
左青青,钱 露,高 瑜,熊 婷,肖佩林,徐 榕,李 林,钱 刚
(1.遵义医科大学 细胞生物学教研室 贵州 遵义 563099;2.遵义医科大学 医学影像学院2016级本科生,贵州 遵义 563099;3.遵义医科大学 临床医学系2017级本科生,贵州 遵义 563099)
全缘千里光(SenecioscandensBuch.-Ham.ex D.Don)是具有良好开发前景的抗菌中草药[1],其功用主要表现为对风热感冒、目赤肿痛、泄泻痢疾、皮肤湿疹疮疖等感染的治疗上[2],现代药理学研究揭示,千里光在抗菌、抗钩端螺旋体、抗滴虫、抗氧化以及自由基清除活性等方面功效显著[3]。
染色体核型分析能够从物种的细胞学水平直观显示染色体数目、形态、整体结构等特征[4-5],物种之间染色体核型差异可以反映物种遗传进化过程的基本规律,从系统植物学的角度揭示物种起源与演化,并由此判断物种间的亲缘关系,阐明该物种的细胞遗传学机制。因此,核型分析成为物种分类研究的重要依据,是研究物种的分化、系统演化途径、亲缘关系、远缘杂交及遗传工程的基本手段[6],被广泛应用于植物分类学研究[7]。
我国的菊科千里光族(Senecioneae)植物种类繁多,包括23属434种[8],迄今,通过细胞遗传学手段对千里光族植物类群开展的研究极其有限,目前报道的仅涉及橐吾属、合耳属、垂头菊属等少数物种[9],这导致千里光族植物各类群间的界定关系不明确且缺乏系统性,物种起源与演化仍没有定论。在此,本文以全缘千里光为研究材料,探讨菊科千里光属近缘物种的演化及系统分类学特征,为千里光杂交育种的细胞遗传学机制提供基础试验资料。
1 材料与方法
1.1 材料 参试材料全缘千里光野生种群采自贵州省遵义地区,经遵义医科大学钱刚教授筛选、检测,选择遗传性状稳定的强抗菌千里光植株为试验材料。
1.2 方法
1.2.1 材料的扦插 选取生长势良好、无损伤、无病虫害的千里光植株,插穗为2~3节的无叶茎段,长度约10±2 cm,其生长前端的腋芽利用600 mg/L NAA处理2 h,无菌水清洗后扦插于5%的KMnO4溶液消毒后的河沙中,置于22~25 ℃、相对湿度85±5%的温室,日常管理用MS液体培养基(10 ×)浇灌以保持表层土壤湿润。30 d后取出扦插株,并用去离子水将根洗净后置于MS液体培养基(10 ×)继续培养。
1.2.2 取材与固定 待根继续延伸至2~3 cm时,剪取长度约0.5 cm根尖于冰水混合物,预处理15 h后,转入新配置的卡诺固定液固定24~48 h。
1.2.3 酶解 将固定后的试验材料置于0.075 mol/L KCL溶液前低渗处理30 min后,用去离子水冲洗3~5遍,于37 ℃恒温环境下,用2.5%果胶酶和2.5%纤维素酶混合液(pH=5.5,由PBS溶解)处理60~90 min之后,用去离子水低渗处理40~60 min,备用。
1.2.4 制片与染色 将试验材料放置于预冷的载玻片上,滴加少量双蒸水以保持根尖处于液体环境,用解剖针压平根尖(长度为2~4 mm),滴加适量卡诺氏固定液,晾干,滴加0.1 μg/mL苯酚品红染液,染色20 min以上,盖上盖玻片后用滤纸吸去多余染液,轻敲盖玻片、压片。
1.2.5 染色体检测与核型分析 染色体标本制备完毕,经显微镜观察,选择25个染色体分散良好、着丝点清晰的分裂相用于染色体形态观察和数目的统计,利用Photoshop CS 软件测量每个细胞的染色体长臂、短臂和总长度。核型分析标准参考李懋学和陈瑞阳(1985)[10]的方法,二点四区系统标准被用于确定着丝粒位置[11],不对称性系数按照Stebbins(1971)[12]方法计算。其中,核型分析相关参数如臂比(Arm ratio)、染色体相对长度(Index of relative length)、核型不对称系数(Asymmetric karyotype coefficient,AKC)和着丝粒指数(Centromere index)的计算公式如下:臂比=长臂长度/短臂长度;染色体相对长度=(染色体长度/染色体组总长)× 100%;核型不对称系数(%)=(染色体长臂总长/全组染色体总长)× 100%;着丝粒指数(%)=(短臂长度/染色体组全长)× 100%。按Levan(1964)的标准划分:臂比在1.0~1.7之间为中部着丝粒染色体;臂比在1.7~3.0之间为亚中部着丝粒染色体;臂比在3.0~7.0为亚端部着丝粒染色体;臂比大于7.0者为端部着丝粒染色体。
2 结果
2.1 染色体组分析 为了确定染色体倍数,我们选择细胞周期中既有处于有丝分裂中期,也有处于有丝分裂中后期的分裂相(见图1A)。当细胞分裂相处于有丝分裂中期时,每条染色体由两条染色单体构成(见图1B);当细胞周期进入有丝分裂中后期,染色体从着丝粒处分离,每条染色体含有1条染色单体,此时细胞并没有一分为二,染色体数目为40条(见图1C),由此确定千里光为二倍体。

A:有丝分裂中期(红色箭头)和有丝分裂中后期(黑色箭头);B:有丝分裂中期的染色体组(黑色箭头);C:有丝分裂中后期的染色体组(黑色箭头)。
2.2 染色体形态 选择细胞分裂相处于有丝分裂中期的图片25张,均未发现染色体形态和结构异常的染色体;通过观察染色体形态结构清晰的其它分裂相的细胞,包括细胞周期处于中后期或后期的分裂相,也未观察到随体和端部着丝粒染色体,染色体形态和结构均相对稳定(见图2)。
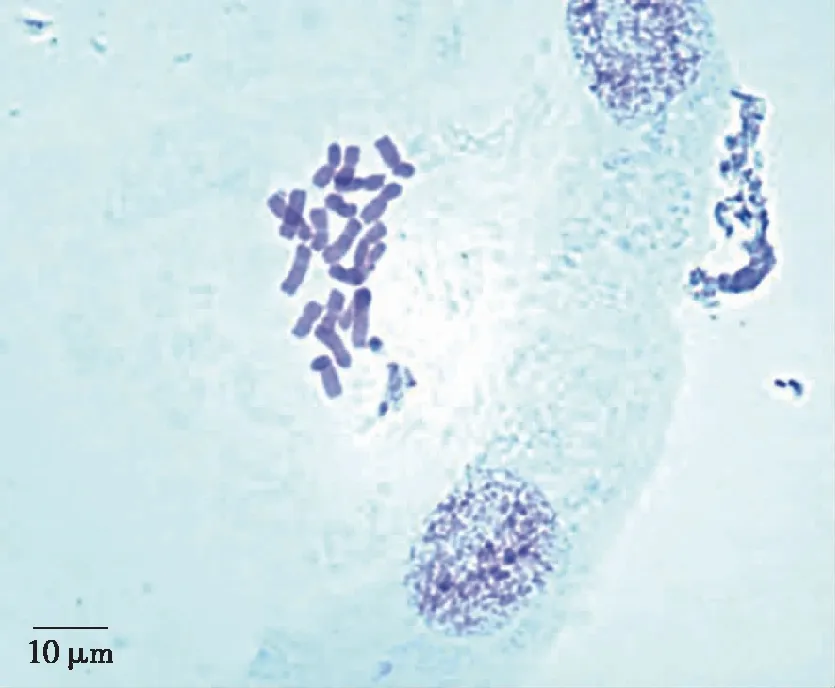
图2 全缘千里光有丝分裂中期的染色体形态
2.3 染色体数目 根据植物核型分析标准,选取染色体分散良好、形态清晰的有丝分裂中期的分裂相,此时每条染色体含有1个着丝粒,每条染色体含有2条染色单体的细胞分裂相,综合有丝分裂中期和中后期着丝粒的个数可确定,千里光的染色体数目为2n=2x=20(见图3)。

图3 全缘千里光染色体核型
本研究发现,10对千里光染色体的组成由8对中部着丝粒染色体(m)和2对亚中部着丝粒染色体(sm)组成,未发现随体,核型公式为:2n=2x=20=8m+2sm(2SAT)(见图4)。
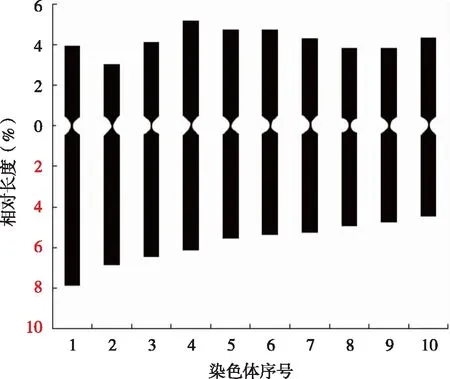
图4 全缘千里光染色体核型模式分析
2.4 染色体核型参数 基于植物核型分析标准,在测量每一组染色体的长臂和短臂之后,根据公式计算其染色体相对长度、臂比、着丝粒指数以及染色体相对长度指数等指标。结果显示,千里光染色体全长在2.168~2.984 μm,相对长度为8.607~11.847%,臂比值介于1.023~2.267之间,平均臂比值1.408,核型不对称系数 为57.78%,最长与最短染色体的比值为1.376。综合相关参数,千里光(菊科-千里光属)的核型属2A型(见表1)。
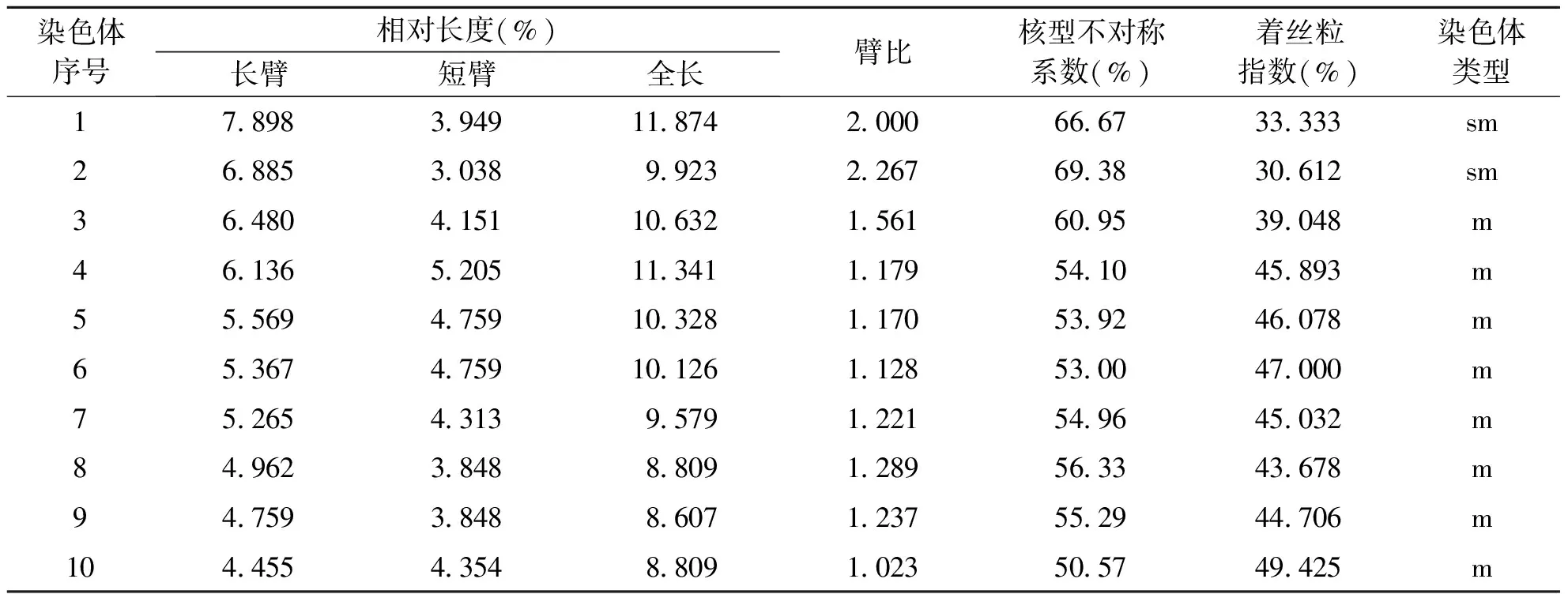
表1 全缘千里光染色体核型参数
3 讨论
关于千里光族植物系统发生与演化,目前得到普遍认同的观点是菊科千里光族早期进化过程中分化为两个相对独立的支系,即千里光支系(X=10)和款冬支系(X=30),之后千里光族近缘物种的演化沿着这两个支系进行[8,13]。目前,有研究对千里光族几个代表性物种的系统植物学研究发现,千里光属(X=10)与橐吾属(X=15)和大风草属(X=15)的遗传关系较远(Senecio);与染色体核型基数相同(X=10)的垂头菊属、合耳菊属的亲缘关系最近[8-9]。本研究结果显示,千里光属中部和亚中部着丝点染色体,未发现多倍体和非整倍体染色体组,表明千里光属植物的遗传特性稳定。此外,Stebbins[12]的研究表明,染色体核型进化的基本趋势是由对称向不对称发展,本研究发现千里光的核型不对称系数介于50.57%和69.38%之间(见表1),即该物种的核型对称性较高,表明千里光属植物是进化速度缓慢、遗传性状稳定的原始物种。
鉴于同一物种的染色体数目、形态和结构等保持相对恒定,植物染色体组的测定及组型分析是细胞遗传学研究的重要内容[14-15],因此,植物核型分析及相关参数不仅有助于揭示植物资源的系统发生、物种演化以及物种间的亲缘关系[2],还有助于解析生物的遗传变异规律和发育机制,进而对作物的遗传育种提供细胞学参考资料[16];于是,核型分析的种间亲缘关系鉴定和植物新品种的选育提供指导作用[17]。本研究中千里光参试资源的核型公式同为2n=2x=20=8m+2sm(2SAT),属于典型的对称性核型,符合2A型分类标准;然而,与已报道的菊科千里光族核型相比较[8],不对称系数、染色体相对长度和臂比值等染色体核型参数存在差异,表明千里光属的不同种或亚种之间的染色体结构存在差异。由此推知,千里光属植物的遗传育种过程中,染色体结构差别可能会对千里光的远缘杂交产生影响,其细胞遗传机制在于染色体配对障碍的生殖隔离可导致杂种不育。
迄今,就千里光属植物的不同种或亚种之间分类标准而言,仍采用传统的植物形态结构进行分类,即以叶片的形态分为掌裂千里光(SinoseneciopalmatisectusC.Jeffreyet Y.L.Chen)、羽叶千里光(SeneciojacobaceaL.)、麻叶千里光(SeneciocannabifoliusLess.)和全缘千里光(SenecioscandensBuch.-Ham.ex D.Don),缺少稳定可靠的细胞和分子水平上的分类依据。于是,有观点认为,物种的不同种或亚种间核型差异主要体现在染色体数目和形态方面的变化,尤其是一些近缘物种[1];因此,在研究植物种、亚种或不同居群之间的亲缘关系时,由于具有相同数目的染色体,染色体形态特征可能是分析物种演化趋势之关键[18]。此外,有研究证实,无论是在核型公式、核型对称性,或者随体的有无,物种不同资源产地的核型均存在细微差异[19],可能源自物种长期进化过程中,因适应环境而导致染色体结构变异以降低植物对环境的选择压力,进而表现为种内核型的多态性;本研究结果与千里光族其它成员之间染色体核型参数的差异[9]也印证了这一观点。
猜你喜欢
云南医药(2021年6期)2022-01-08
临床检验杂志(2021年10期)2021-11-24
川北医学院学报(2021年6期)2021-07-13
科学大众(中学)(2019年3期)2019-05-17
科学之谜(2019年3期)2019-03-28
汽车观察(2018年10期)2018-11-06
科学之谜(2018年8期)2018-09-29
科技知识动漫(2017年1期)2017-02-06
中学生理科应试(2016年4期)2016-11-19
恋爱婚姻家庭·养生版(2016年9期)2016-09-07
