
水文连通强度对芦苇、碱蓬生态化学计量的影响
2020-08-24 14:55刘佳凯王宇张振明张明祥
生态科学 2020年4期
刘佳凯, 王宇, 张振明, 张明祥
水文连通强度对芦苇、碱蓬生态化学计量的影响
刘佳凯, 王宇, 张振明, 张明祥*
北京林业大学, 生态与自然保护学院, 北京 100083
黄河三角洲以芦苇()和盐地碱蓬()等植物建立的湿地生态系统对维持区域生态安全有着十分重要的作用。近年来由于受到人类活动和气候变化的双重影响, 该区域湿地出现了不同程度的退化。目前黄河三角洲自然保护区正在探索引入水文连通理论指导退化湿地的恢复和重建工作。本研究以实验室模拟结合生态化学计量方法, 探究不同水文连通强度的补水修复措施对建群种芦苇和碱蓬的生长及化学计量的影响。研究结果表明较弱的水文连通强度有利于碱蓬生长, 而水文连通强度对芦苇生长影响不显著; 碱蓬体内氮磷元素含量随水文连通减弱而增加, 芦苇体内氮磷元素含量则不受水文连通强度影响。此外, 植物体内氮磷比主要取决于土壤内的氮磷元素含量, 水文连通对植物体内氮磷比的影响并不显著。
化学计量; 水文连通强度; 黄河三角洲; 芦苇; 碱蓬
0 前言
黄河三角洲位于渤海南岸, 是我国最完整且最年轻的滨海湿地, 由黄河大量的泥沙淤积于河口区域而形成[1]。黄河三角洲是许多重要鸟类迁徙路线重要的停歇地或栖息地, 同时还起着维持湿地系统正常演替和调节气候等多重重要功能[2], 因此以芦苇()和碱蓬()等植物建立的湿地生态系统对维持区域生态安全有着十分重要的作用。但近年来, 在人类活动和气候变化的双胁迫下, 黄河三角洲的湿地生态系统遭到了较为严重的破坏和不同程度的退化[3, 4], 如何有效的恢复受损湿地成为黄河三角洲保护的重要任务。
水文连通是指水或者以水为介质的物质和能量在水文循环各要素中迁移的过程[5]。量化水文连通强度有两类: 静态水文连通指数和动态水文连通指数[6]。静态水文连通指数是指水分运动过程中经过的各要素的组成结构, 最为常用的方法包括基于地形的水流路径长度计算[7]和基于土壤湿度的热熵值法或积分联通尺度法[8, 9]。动态水文连通是指连接两个要素所需要的水量[6, 10], 通常量化动态水文连通强度的参数包括水淹深度, 水淹频率等。在研究过程中, 通常以静态水文连通指数探究径流或侵蚀与下垫面结构的关系, 而动态水文连通一般用于解释水文过程和其他生态过程之间的关系。目前黄河三角洲自然保护区正在探索引入水文连通理论指导退化湿地的恢复和重建工作, 但是目前仍然有很多问题尚不明确, 其中不同的水文连通强度对主要植物群落的影响就是其中最重要的理论问题之一。
目前判断湿地植物群落是否稳定的方法有很多, 最经典的方法是进行野外固定样地监测[11], 但该方法需要的时间较长, 且一旦发现植物群落受到影响时, 生态过程可能已经遭受了不可逆的破坏[12], 此时再实施修复措施可能为时已晚。除此以外, 生态化学计量方法是近年来新型的方法和研究热点[13-15]。生态化学计量方法是通过研究植物或环境内的主要元素含量和各元素比例关系的变化来研究生态系统是否稳定[13], 其基础理论是生物在变化的环境中具有保持其自身化学组成的能力, 形成一定的内稳定机制[16],因此当植物体内氮磷含量或者氮磷比发生显著的变化时, 说明该植物群落或生态系统会发生相应的变化。前人研究指出, 纬度、年均温度会对植物叶片中氮磷比产生影响[17], 而内稳定性越强的植物具有较高的优势度[18, 19]。
基于此, 本研究以实验室模拟结合生态化学计量方法, 探究黄河三角洲不同水文连通强度的补水修复措施对当地建群种芦苇和碱蓬的生长及化学计量的影响, 以对黄河三角洲的湿地恢复工作提供理论基础。
1 材料与方法
1.1 实验材料
本研究的总体思路是结合野外采样与室内实验, 实验用土壤采自北京林业大学八家苗圃基地。实验材料芦苇和盐地碱蓬于2017年5月采自山东省黄河三角洲自然保护区, 每种植物480株。芦苇和盐地碱蓬的采集皆使用挖掘法并尽量保存植株地上地下部分的完整性, 采集当天套袋带回, 隔天将实验植物移栽至实验用的水槽中培养14 d至植物稳定。实验所用营养液按照黄河三角洲恢复区内的水质配比而成, 其中TN, TP和盐度分别为4.86 mg·L-1, 1.67 mg·L-1和3‰, pH值为8.0。
1.2 实验设置
本研究中以水淹深度和水淹周期量化水文连通强度; 以有无水淹以及水淹周期的短长表征水文连通的强弱。已有研究表明, 当淹水深度为10 cm时芦苇具有最高的生长量[20], 且目前黄河三角洲恢复区补水周期分别设置为3周和5周, 基于此, 本研究设置实验共8组(表1), 每组6个重复, 分别探讨水淹深度和水淹周期两个动态水文连通指数对芦苇及碱蓬生物化学计量的影响。实验植物芦苇和盐地碱蓬种植于长1 m, 宽0.2 m, 高0.25 m的水槽(图1)中, 种植密度与恢复区芦苇和碱蓬的平均生长密度保持一致[21, 22]。实验开始时, 种植的所有芦苇和碱蓬株高为10 cm, 在PD1、PD2、SD1和SD2组各个重复中加入20 L营养液, 之后再分别在PD2和SD2组各个重复中加入10 L蒸馏水保证其淹没植物10 cm。之后每3 d对各个重复用蒸馏水进行补水, 保证预设的水淹深度并记录每次的补水量(L)。在PP1、PP2、SP1和SP2各个重复中加入30 L营养液, 保证其淹没植物10 cm, 并按照淹水周期定期放水, 在放水结束后再重新加入30 L营养液。
实验从6月开始为期70 d, 在实验开始和结束阶段分别测量每个重复中的植株高度、茎叶中的氮磷含量、水体中TN和TP含量及生物量, 此外实验中每10 d将对植物的株高和水体中的TN及TP含量进行一次测量。最后记录各组植株高度增长(cm), 净生长量(kg·m-2), 植物茎和叶及整株的氮磷含量(mg·g-1)以及茎叶及植株的氮磷比(N/T)。

表1 实验设置情况表

图1 实验装置布设图
Figure 1 The diagram of experiment groove
1.3 数据分析
本研究以PD1和PD2组测得数据进行置信度为95%的单因素方差分析, 研究水淹深度对芦苇生长及化学计量的影响; 同样的方法对SD1和SD2各数据进行分析研究水淹深度对碱蓬生长及化学计量的影响。对PP1和PP2以及SP1以及SP1和SP2的各数据进行分析以得出两种不同的水淹周期分别对芦苇和碱蓬碱的生长及化学计量的影响。
2 结果与分析
2.1 水文连通强度对植物生长的影响
图2显示了不同水淹强度下芦苇(图A)和碱蓬(图B)植株高度增长量和净生长量。在为期70 d的实验过程中, 在没有水淹的条件下, 芦苇由10 cm增长至(39.58 ± 0.50) cm; 在水淹10 cm条件下, 高度由10 cm增长至(40.19 ± 0.41) cm。水淹与否两种条件对芦苇的高度影响没有明显差异(p = 0.94)。无水淹时芦苇生物量为(1.68 ± 0.16) kg·m-2, 水淹条件下芦苇的净生长量为(1.74 ± 0.09) kg·m-2, 两者差异不显著(p = 0.11)。对于碱蓬而言, 在没有水淹的情况下, 植株高度由初始状态的10 cm增加至(16.86 ± 0.58) cm; 在水淹条件下, 植株高度由10 cm增长至(16.24 ± 0.30) cm, 两者没有显著差异(p = 0.51)。无水淹时碱蓬净生长量为(0.43 ± 0.032) kg·m-2, 显著高于水淹10 cm条件下碱蓬的生物量(0.32 ± 0.03 kg·m-2) (p = 0.02)。
图3显示了不同水淹周期下芦苇(图A)和碱蓬(图B)的植株高度生长量和净生长量。对于芦苇而言,当水淹周期为2周时, 芦苇高度由最初的10 cm增长至(39.76 ± 0.69) cm; 当灌溉时间为4周时, 芦苇高度由10 cm增长至(40.01 ± 0.34) cm。不同水淹周期对芦苇高度增长影响差异不显著(p = 0.99)。灌溉周期为2周的芦苇净生长量为(1.71 ± 0.16) kg·m-2, 水淹4周的生物增长量为(1.71 ± 0.09) kg·m-2, 二者差异不显著(p = 0.08)。对于碱蓬而言, 当水淹周期为2周时, 植株高度由初始株高10 cm增长至(16.74 ± 0.49) cm; 当灌溉时间为4周时, 植株高度由最初的10 cm增长至(16.35 ± 0.56) cm, 植株高度增长量显著小于2周水淹周期(p = 0.01)。

图2 不同水淹深度下芦苇(A)碱蓬(B)生长状况
Figure 2 Growth status of(A) and(B) under different water depth

图3 不同水淹周期下芦苇(A)碱蓬(B)生长状况
Figure 3 Growth status of(A) and(B) under different water logging period
综上, 在无水淹条件下, 碱蓬的生长量高于水淹条件, 而不同的水淹周期下, 碱蓬的生长量没有显著差异, 但在较短水淹周期下植株高度统计学上显著高于较长的水淹周期, 但绝对值相差不到植株平均高度的6%。而有无水淹及水淹周期对芦苇的生长没有显著影响。既较弱的水文连通强度一定程度上有利于碱蓬生长, 而水文连通强度对芦苇生长影响较弱。
2.2 水文连通强度对植物体内N, P含量的影响
图4显示了在不同水淹强度下芦苇(图A)和碱蓬(图B)茎叶中的氮磷含量的变化量。无水淹和水淹条件下, 芦苇茎的氮含量增长量分别为(2.13 ± 0.16) mg·g-1和(2.23 ± 0.19) mg·g-1, 差异不显著(p = 0.73); 叶的氮含量增长量分别为(1.49 ± 0.07) mg·g-1和(1.53 ± 0.07) mg·g-1, 差异不显著(p = 0.87); 茎磷含量增长量分别为(0.49 ± 0.05) mg·g-1和(0.51 ± 0.05) mg·g-1, 差异不显著(p = 0.92); 叶磷含量增长量分别为(0.49 ± 0.05) mg·g-1和(0.53 ± 0.04) mg·g-1, 差异不显著(p = 0.88)。
对于碱蓬而言, 在不同水淹条件下, 无水淹和有水淹时茎的氮含量增长量分别为(0.64 ± 0.06) mg·g-1和(0.52 ± 0.11) mg·g-1, 差异不显著(p = 0.46); 叶的氮含量增长分别为(0.46 ± 0.14) mg·g-1和(0.29 ± 0.03) mg·g-1, 水淹条件显著低于无水淹条件(p = 0.023); 茎磷含量增长分别为(0.18 ± 0.02) mg·g-1和(0.10 ± 0.02) mg·g-1, 水淹条件显著低于无水淹条件(p = 0.033); 叶磷含量增长分别为(0.18 ± 0.03) mg·g-1和(0.11 ± 0.02) mg·g-1, 无显著差异(p = 0.47)。
图5显示了在不同水淹周期下芦苇(图A)和碱蓬(图B)茎叶中的氮磷含量的变化量。水淹周期为2周和4周时芦苇茎的氮含量增长量分别为(2.19 ± 0.03) mg·g-1和(2.18 ± 0.02) mg·g-1, 差异不显著(p = 0.97); 叶的氮含量增长量分别为(1.51 ± 0.09) mg·g-1和(1.52 ± 0.10) mg·g-1, 差异不显著(p = 0.87); 茎的磷含量增长量分别为(0.49 ± 0.07) mg·g-1和(0.51 ± 0.05) mg·g-1, 差异不显著(p = 0.91);叶的磷含量增长量分别为(0.53 ± 0.04) mg·g-1和(0.55 ± 0.04) mg·g-1, 差异不显著(p = 0.91)。
对碱蓬而言, 水淹周期为2周和4周时茎的氮含量增长量分别为(0.62 ± 0.11) mg·g-1和(0.55 ± 0.09) mg·g-1, 差异不显著(p = 0.65); 叶的氮含量增长量分别为(0.47 ± 0.02) mg·g-1和(0.29 ± 0.01) mg·g-1, 水淹两周条件下的增长量显著高于水淹四周(p = 0.022); 茎的磷含量增长量分别为(0.17 ± 0.05) mg·g-1和(0.11 ± 0.02) mg·g-1, 差异不显著(p = 0.52); 叶的磷含量增长量分别为(0.18 ± 0.01) mg·g-1和(0.11 ± 0.03) mg·g-1,水淹两周条件下的增长量显著高于水淹四周(p = 0.03)。
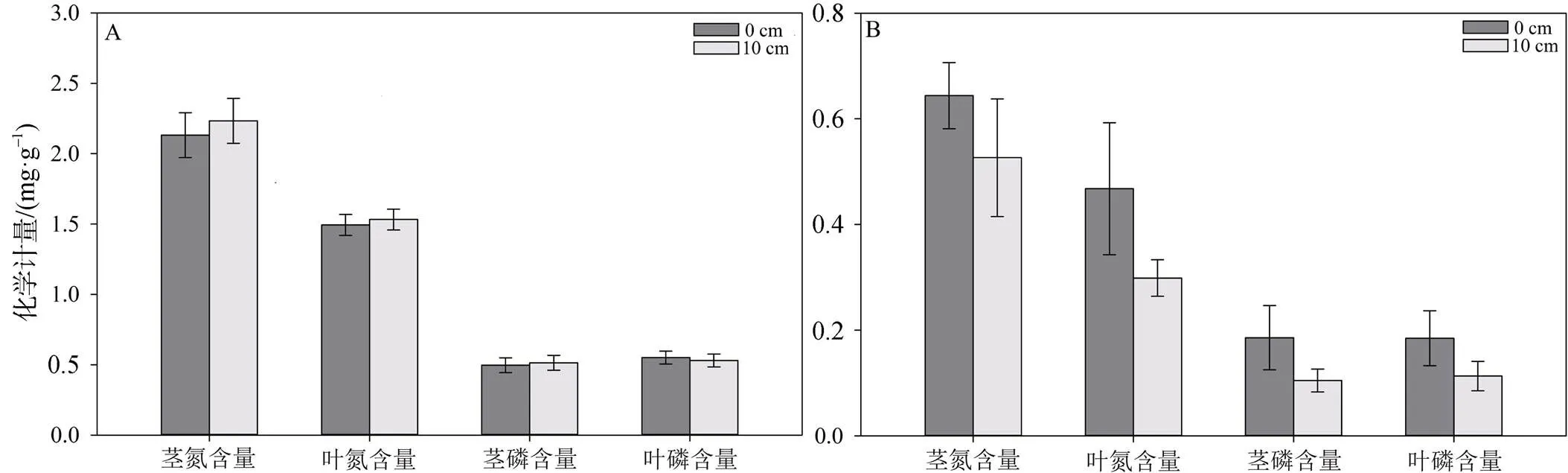
图4 不同水淹深度下芦苇(A)碱蓬(B)植物体内氮、磷含量变化
Figure 4 Changes of nitrogen and phosphorus content of(A) and(B) under different water depth
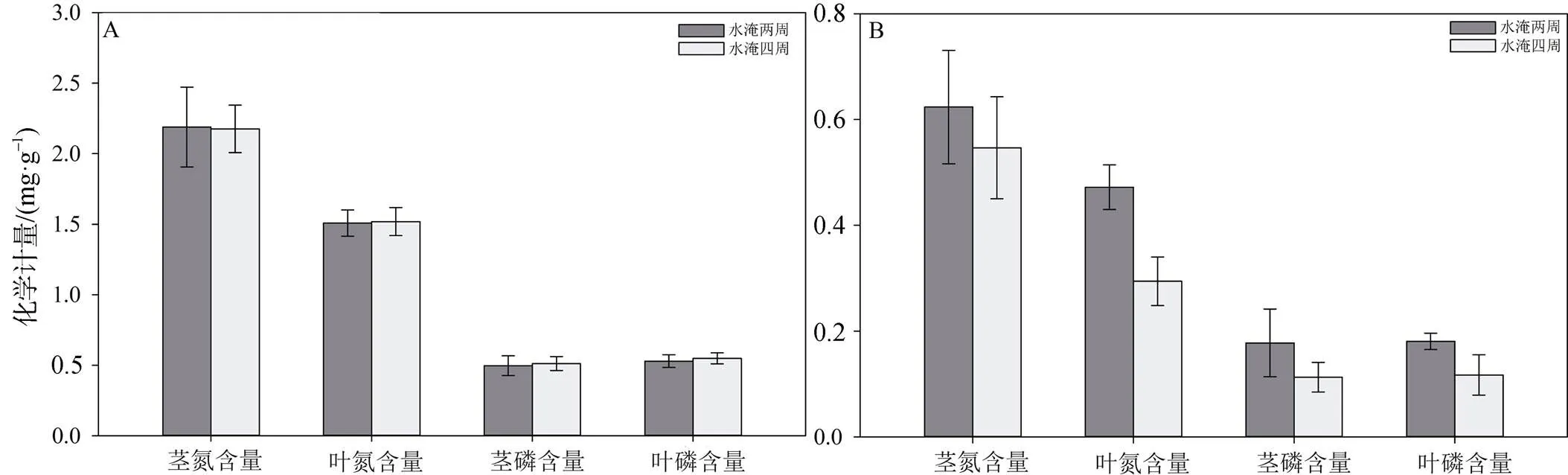
图5 不同水淹周期下芦苇(A)碱蓬(B)植物体内氮、磷含量变化
Figure 5 Changes of nitrogen and phosphorus content of(A) and(B) under different water logging period
综上, 水淹强度和水淹周期对芦苇茎叶吸收氮磷元素基本没有影响, 而有水淹条件下碱蓬叶的氮含量和茎的磷含量增量显著低于无水淹条件, 并且较短的水淹周期下碱蓬叶的氮磷含量增长量显著高于较长水淹周期。即水文连通强度对芦苇植物体内氮磷元素累积无显著影响影而较强的水文连通则会降低碱蓬对氮磷元素的累积。
2.3 水文连通强度对植物N/P的影响
不同水淹深度和水淹周期下芦苇和碱蓬在实验开始和结束时茎叶内氮磷比如表2所示, 在不同水淹条件下, 芦苇和碱蓬茎中的氮磷比都较为稳定, 在24左右, 而叶中的氮磷比则皆由一个较大值急剧减小到平均值为23.31和12.74。这是由于在实验开始时种植的芦苇和碱蓬叶内磷含量十分低, 而在生长过程中叶中的磷逐渐累积[1]。在生长期结束后, 不同的水文连通强度并没有对芦苇或碱蓬茎、叶内的氮磷比产生显著影响, 各组之间氮磷比差异不显著。既水文连通强度对植物体内氮磷比影响不显著。
3 讨论
本研究在进行实验种植时参考了黄河三角洲恢复区芦苇群落和碱蓬群落的生长密度[21-23], 在生长期结束后, 黄河三角洲恢复区内芦苇和碱蓬的生物量为2.52 kg·m-2和0.65 kg·m-2, 在本研究中芦苇和碱蓬的平均生物量为 2.41 kg·m-2和0.72 kg·m-2。说明实验室条件下和野外条件下植物生长情况相似, 模拟实验的结果可以反应真实野外条件下的情况。
植物体内的氮元素对氨基酸、蛋白质等物质的合成以及提高植物光合作用能力有着重要作用, 而磷元素则是核酸和酶的重要组成部分[24], N/P的稳定对植物的生长和种群的建立及稳定都有着至关重要的作用[25]。在本研究中, 不同的水文连通强度对芦苇和碱蓬的N/P值没有显著影响, 其在生长结束时平均值分别为(23.82 ± 1.24)和(13.17 ± 1.01)。其余相关研究结果如表3所示。一般认为当植物中N/P比大于16时, 植物生长主要受到磷含量的限制, 而当N/P小于14时则主要受到氮元素的限制[26]。结合本研究的实验室模拟研究结果和屈凡柱等在黄河三角洲的野外调查研究结果[24]可得知, 恢复区芦苇生长主要受到磷元素的限制, 而在实验室条件下, 芦苇生长主要受到氮元素缺乏的限制, 实验室所用的基质土壤来自黄河三角洲恢复区, 淹水实验用水水质参照黄河淡水水质配比。限制条件的偏差来自于定期的海水入侵: 恢复区在每个月大潮期间会接受一次海水入侵, 而海水含有大量氮元素, 却缺乏磷元素, 这就导致了恢复区内氮元素过剩而芦苇生长主要受限于磷元素缺乏。而无论在恢复区还是实验室条件下, 碱蓬的生长皆主要受到氮元素的限制。

表2 不同水淹深度和水淹周期下植物茎叶内氮磷比变化

表3 不同研究中芦苇、碱蓬化学计量对比
在黄河三角洲内, 不同区域限制植物生长的因子也各不相同, 在靠近海岸线一侧, 分布于黄河三角洲低潮滩中的碱蓬主要受氮元素缺乏的限制,[1]而分布在潮上带的芦苇生长都主要受磷元素的限制; 分布于这一区域的芦苇植物体内平均氮磷含量分别为2.4 mg·g-1和0.19 mg·g-1; 碱蓬体内平均氮磷含量分别为1.4 mg·g-1g和0.11 mg·g-1。而在磷元素供应充足的条件下, 芦苇植物体内的磷含量会达到该数值的2~3倍, 而氮含量则会达到3~4倍[27]; 此外, 对于碱蓬而言, 当氮供应充足时, 碱蓬叶片氮磷含量可以分别达到30.02 mg·g-1和1.81 mg·g-1, 皆可达到黄河三角洲碱蓬体内氮磷含量的10倍以上N/P则可达到16.58以上[28, 29]。由此可见, 当背景环境参数如土壤氮、磷含量, 土壤氮磷比[1]等发生变化时, 芦苇和碱蓬体内氮磷元素的变化范围较大, 其内稳定性较弱[30-33]。在本研究中, 水文连通强度的改变对芦苇和碱蓬的生长量及化学计量的影响不大, 因此目前黄河三角洲恢复区所实行的不同补水方案对植物生长及群落特征影响较小。
不同的水淹条件下, 植物对水体中TP和TN的吸收存在一定的差异[36-38]。在水深为2 cm和15 cm水淹条件下, 芦苇对水中TN(初始浓度5 mg·L-1)的平均去除率分别为74.8%和38.1%, 对TP(初始浓度0.2 mg·L-1)的平均去除率分别为74.8%和38.1%[34]。此外, 当水中TN和TP的初始浓度为4.6 mg·L-1和2.0 mg·L-1的条件下, 种植于浮岛的碱蓬对二者的去除率分别为68.4%和56.7[35]。其余若干研究也指出, 湿地植物对废水中氮磷等营养元素的去除有着至关重要的作用[36-38], 却少数有研究量化过植物的贡献率。然而本研究在无水淹和水淹10 cm两个实验组中发现, 水中TN和TP的平均去除率分别为58.3%和47.2%, 但是由碱蓬吸收的TN和TP仅占去除量的1.07%和0.89%; 芦苇对TN和TP的吸收仅占去除量的11.46%和6.18%, 其余的氮磷则累积在土壤中或是直接进入大气; 植物对水体中营养物质去除的贡献率较小。然而本研究并没有对元素进行同位素示踪, 因此还需要进一步实验才能得出更加明确的结论。
4 结论
(1)强水文连通补水策略会降低碱蓬体内氮磷元素的累积量, 并影响碱蓬生长; 而芦苇体内氮磷元素累积量和生长量皆不受水文连通强度的影响。
(2)水文连通强度的改变不会影响芦苇和碱蓬体内氮磷比。而相比水文连通强度, 背景环境如土壤和水的氮磷含量对芦苇和碱蓬的化学计量都会产生更为显著的影响
(3)综合上述研究结果, 建议黄河三角洲采取较弱强度的水文连通补水策略, 在保证恢复区内碱蓬的正常生长的同时尽量减少淡水资源的使用以保证足够的黄河水水量。
[1] 孙志高, 宋红丽, 胡星云. 黄河口潮滩碱蓬种子萌发与幼苗生长对土壤中氮磷比的响应[J]. 湿地科学, 2017, 15(1): 10–19.
[2] ZHANG X L, CHEN D J, XU Z J, et al. Service value of ecosystem of coastal wetlands in the Yellow River Delta.[J]. Science & Technology Review, 2009, 27(10): 37–42.
[3] QI S, LIU H. Natural and anthropogenic hazards in the Yellow River Delta, China[J]. Natural Hazards, 2017, 85(3): 1907–1911.
[4] ZHANG X, WANG L, FU X, et al. Ecological vulnerability assessment based on PSSR in Yellow River Delta[J]. Journal of Cleaner Production, 2017, 167: 1106–1111.
[5] Freeman M C, Pringle C M, Jackson C R. Hydrologic Connectivity and the Contribution of Stream Headwaters to Ecological Integrity at Regional Scales[J]. Jawra Journal of the American Water Resources Association, 2010, 43(1): 5–14.
[6] BRACKEN L J, WAINWRIGHT J, ALI G A, et al. Concepts of hydrological connectivity: Research approaches, pathways and future agendas[J]. Earth-Science Reviews, 2013, 119: 17–34.
[7] LANE S N, REANEY S M, HEATHWAITE A L. Representation of landscape hydrological connectivity using a topographically driven surface flow index[J]. Water Resources Research, 2009, 45(8).
[8] MCGUIRE K J, MCDONNELL J J. Hydrological connectivity of hillslopes and streams: Characteristic time scales and nonlinearities[J]. Water Resources Research, 2010, 46(10).
[9] ALI G A, ROY A G. Shopping for hydrologically representative connectivity metrics in a humid temperate forested catchment[J]. Water Resources Research, 2010, 46(12): 65–74.
[10] BACHMAIR S, WEILER M. Hillslope characteristics as controls of subsurface flow variability[J]. Hydrology and Earth System Sciences, 2012, 16(10): 3699–3715.
[11] 李满良. 北京湿地主要植物群落对水因子的响应[D]. 北京: 首都师范大学, 2006.
[12] 何池全, 赵魁义, 余国营, 等. 湿地生态过程研究进展[J]. 地球科学进展, 2000, 15(2): 164–171.
[13] 贺金生, 韩兴国. 生态化学计量学: 探索从个体到生态系统的统一化理论[J]. 植物生态学报, 2010, 34(1): 2–6.
[14] 程滨, 赵永军, 张文广, 等. 生态化学计量学研究进展[J]. 生态学报, 2010, 30(6): 1628–1637.
[15] ELSER J J, STERNER R W, GOROKHOVA E, et al. Biological stoichiometry from genes to ecosystems[J]. Ecology Letters, 2010, 3(6): 540–550.
[16] STERNER R W, ELSER J J. Ecological Stoichiometry: The Biology of Elements From Molecules to The Biosphere[M]. Princeton, N.J.: Princeton University Press, 2002: 272–281.
[17] 任书杰, 于贵瑞, 陶波, 等. 中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究[J]. 环境科学, 2007, 28(12): 2665–2673.
[18] HE J S, FANG J, WANG Z, et al. Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China[J]. Oecologia, 2006, 149(1): 115–122.
[19] YU Q, CHEN Q, ELSER J J, et al. Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability[J]. Ecology Letters, 2010, 13(11): 1390–1399.
[20] WANG X, ZHANG D, GUAN B, et al. Optimum water supplement strategy to restore reed wetland in the Yellow River Delta[J]. PLOS ONE, 2017, 12(5): e177692.
[21] 李敏, 刘帅帅, 徐飞. 黄河三角洲芦苇、盐地碱蓬单生与混生群落生长差异分析[J]. 山东农业科学, 2017, (03): 52–57.
[22] 李敏, 刘帅帅, 徐飞. 黄河三角洲芦苇—碱蓬群落结构和功能对3种人为干扰的响应[J]. 湿地科学, 2017, (01): 92–98.
[23] 李敏. 不同处理对黄河三角洲芦苇—碱蓬群落结构和功能的影响[D]. 济南: 山东师范大学, 2017: 45-87.
[24] 屈凡柱, 孟灵, 付战勇, 等. 不同生境条件下滨海芦苇湿地C、N、P化学计量特征[J]. 生态学报, 2018, 38(5): 1731–1738.
[25] 王维奇, 徐玲琳, 曾从盛, 等. 河口湿地植物活体-枯落物-土壤的碳氮磷生态化学计量特征[J]. 生态学报, 2011, 31(23): 7119–7124.
[26] KOERSELMAN W. The Vegetation N: P Ratio: a New Tool to Detect the Nature of Nutrient Limitation[J]. Journal of Applied Ecology, 1996, 33(6): 1441–1450.
[27] 朱秋丽. 闽浙沪滨海湿地草本植物碳氮磷生态化学计量特征[D]. 福州: 福建师范大学, 2017.
[28] 冉越男, 王光美, 于君宝, 等. 土壤氮磷供应变化条件下盐地碱蓬叶片碳、氮和磷的化学计量特征[J]. 湿地科学, 2017, 15(1): 148–156.
[29] 冉越男. 氮磷供应条件对盐地碱蓬生长及生态化学计量特征的影响[D]. 烟台: 中国科学院烟台海岸带研究所, 2017: 25-48.
[30] 蒋利玲, 何诗, 吴丽凤, 等. 闽江河口湿地3种植物化学计量内稳性特征[J]. 湿地科学, 2014, (3): 293–298.
[31] 刘长发, 陶韦, 武嘉文, 等. 翅碱蓬氮、磷内稳性与生长对模拟氮、磷沉降的响应[C]//中国海洋湖沼学会.“全球变化下的海洋与湖沼生态安全”学术交流会论文摘要集.北京: 中国海洋湖沼学会, 2014:113.
[32] QU F, YU J, DU S, et al. Influences of anthropogenic cultivation on C, N and P stoichiometry of reed-dominated coastal wetlands in the Yellow River Delta[J]. Geoderma, 2014, 235-236: 227–232.
[33] 刘兴华, 陈为峰, 段存国, 等. 黄河三角洲未利用地开发对植物与土壤碳、氮、磷化学计量特征的影响[J]. 水土保持学报, 2013, 27(2): 204–208.
[34]张葆华, 吴德意, 叶春, 等. 不同水深条件下芦苇湿地对氮磷的去除研究[J]. 环境科学与技术, 2007, 30(7): 4–6.
[35] 潘军标, 王栋, 王趁义, 等. 碱蓬对富营养化海水养殖水体中氮磷的去除研究[J]. 环境保护科学, 2018(2), 44(02): 37–41.
[36] LIU J L, LIU J K, ANDERSON J T, et al. Potential of aquatic macrophytes and artificial floating island for removing contaminants[J]. Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology, 2016, 150(4): 702–709.
[37] LIU J, LIU J, ZHANG R, et al. Impacts of aquatic macrophytes configuration modes on water quality[J]. Water Science and Technology, 2014, 69(2): 253–261.
[38] 张振明, 刘佳凯, 刘晶岚, 等. 不同挺水植物对水体净化效果及生长特性研究[J]. 灌溉排水学报, 2013, (01): 139–141.
The influence of hydrological connectivity on stoichiometry ofand
LIU Jiakai, WANG Yu, ZHANG Zhenming, ZHANG Mingxiang*
Beijing Forestry University, School of Ecology and Nature Conservation, Beijing 100083, China
Yellow river delta, dominated byand, has many important ecological service function such as habitat supporting and is crucial important for regional ecological safety. While in recent decades, this coastal wetland has been suffering serious degradation due to anthropological activity and climate change. Local government and managers in Yellow River Delta National Conservation now are seeking for efficient measurement to restore the degraded ecosystem based on the concept of hydrological connectivity (HC). Thus, the current study aims to find the influence of HC on local species in simulation experiments combined with stoichiometry thesis. The results show low intensive HC is conducive to the growth ofand high intensive HC is also positively correlated to the accumulation of TP in the leaves of. While different HC conditions have no significant influence on the growth ofor the accumulation of TN and TP in this species. Besides, HC has insignificant influence on the N/P inor. Compared with previous studies, the TN and TP concentration or N/P in soils and other environment conditions will influence the stoichiometry ofandmore dramatically.
stoichiometry; hydrological connectivity; Yellow River Delta;;
10.14108/j.cnki.1008-8873.2020.04.011
刘佳凯, 王宇, 张振明, 等. 水文连通强度对芦苇、碱蓬生态化学计量的影响[J]. 生态科学, 2020, 39(4): 81–88.
LIU Jiakai, WANG Yu, ZHANG Zhenming, et al. The influence of hydrological connectivity on stoichiometry ofand[J]. Ecological Science, 2020, 39(4): 81–88.
Q948
A
1008-8873(2020)04-081-08
2019-03-20;
2019-05-15
国家重点研发计划(2017YFC0505903)
刘佳凯(1991—), 男, 重庆人, 博士研究生, 从事湿地学研究, E-mail: timberfield1991@163.com
张明祥(1972—), 男, 教授, 从事湿地生态学研究, E-mail: zhangmingxiang@bjfu.edu.cn
猜你喜欢
今日农业(2022年14期)2022-11-10
汽车实用技术(2022年16期)2022-09-03
现代园艺(2020年9期)2020-05-24
现代园艺(2019年1期)2019-02-18
南方农业学报(2018年12期)2018-09-10
小哥白尼(军事科学)(2018年3期)2018-06-15
安徽农学通报(2017年12期)2017-07-13
中国烟草学报(2012年1期)2012-04-09
天然气勘探与开发(2012年2期)2012-03-25
现代农业科技(2009年19期)2009-03-20
