
萎锈灵包衣对低温胁迫下棉种萌发特性的影响*
2020-08-26 08:31周小云刘梦丽张军高杜鹏程翟梦华甘恩祥高文伟
中国农业气象 2020年8期
周小云,刘梦丽,李 进,张军高,梁 晶,杜鹏程,王 莉,翟梦华,甘恩祥,高文伟,雷 斌**
萎锈灵包衣对低温胁迫下棉种萌发特性的影响*
周小云1,刘梦丽2,李 进1,张军高1,梁 晶1,杜鹏程2,王 莉2,翟梦华2,甘恩祥2,高文伟2,雷 斌1**
(1.新疆农业科学院核技术生物技术研究所/农业部荒漠绿洲作物生理生态与耕作重点实验室,乌鲁木齐 830091;2.新疆农业大学农学院,乌鲁木齐 830052)
为减轻低温胁迫对棉花种子萌发能力的影响,探索萎锈灵包衣提高棉种萌发期耐低温能力的功效。以对温度敏感的“新陆早50”棉种为实验材料,设置25℃、18℃、15℃和12℃共4个温度处理,将萎锈灵按照不同含量制成悬浮种衣剂即0(种衣剂中仅含惰性成分)、4.2%、5.7%、7.2%、8.7%和10.2%共6个梯度对棉种进行包衣处理,以光籽不包衣处理为对照,采用砂培法,观测各处理条件下的发芽势、发芽率、发芽指数以及活力指数等种子萌发活性指标和干物质质量、种子干物质转化效率以及呼吸消耗量等干物质转移指标,计算低温胁迫与正常温度下相比的抗逆系数,比较不同含量萎锈灵包衣对低温胁迫下棉花种子萌发的影响。结果表明:在不包衣的情况下,正常温度下(25℃时)棉种的萌发指标最高,发芽率可达80.5%,随着温度降低棉种萌发指标均呈下降趋势,说明低温胁迫严重影响棉种的萌发;而在萎锈灵包衣的情况下,正常温度下(25℃时)棉种的萌发指标最高,发芽率可达96.0%,随着温度降低各处理棉种萌发指标均呈缓慢下降趋势,说明萎锈灵能够明显缓解低温胁迫对种子萌发的伤害。在相同温度条件下,随着种衣剂中萎锈灵含量的增加,除种子的呼吸消耗量呈先下降后上升的趋势外,种子发芽率等萌发指标以及鲜重、干物质质量、干物质转化效率指标均呈先上升后下降的趋势;萎锈灵处理较不含萎锈灵处理能使棉种萌发进程提早2~4d;低温胁迫下不含萎锈灵处理的种子活力低,棉花幼苗生长也受到抑制,幼苗鲜重显著减少,导致幼苗干物质积累也显著减少;未包衣和不含萎锈灵处理的种子平均抗逆系数仅分别为0.68和0.70,而含萎锈灵包衣的种子平均抗逆系数为0.73~0.91。研究表明萎锈灵能增强棉花种子活力,进而提早萌发进程,提高发芽率,促进幼苗生长,从而增强棉花萌发期的耐低温能力。
萎锈灵;发芽率;发芽指数;活力指数;抗逆系数;干物质转化效率;棉花;种子萌发;低温胁迫
棉花是新疆的支柱产业,随着棉花生产的高速发展,2018年新疆棉花产量分别占全国和全球棉花总产的83.8%和19.7%[1]。新疆作为全国最大棉区,栽培中经常遭遇“倒春寒”等低温气象灾害,导致每年有不同程度的烂种、烂芽、死苗现象发生,造成“出苗难、保苗难”,影响植株生长发育和产量、品质形成。1961−2016年新疆棉区4月下旬−5月上旬发生低温冷害16次,此期正值棉花出苗和幼苗生长关键期,低温冷害会造成棉苗死亡或延迟生长[2],成为制约棉花产量和品质提高的重要因子。随着种子包衣技术的发展,种衣剂包衣棉种措施有效解决了正常年份棉田上述“出苗难、保苗难”问题。低温胁迫下,5mmol·L−1水杨酸包衣处理能缓解低温胁迫对幼苗生长的抑制,提高棉花幼苗的抗寒性[3];18.6%拌·福·乙种衣剂能提高棉种发芽率和出苗率,促进棉花幼苗茎秆增粗、根系伸长、生物量积累[4];具有抗寒功能的种衣剂能促棉种萌发,提高发芽率,保护棉苗细胞超微结构稳定,维持正常生长,增强幼苗抵御低温的能力[5]。但是,在低温危害重的年份仍然难以确保田间较高保苗率,迫切需要研究提高棉花苗期耐低温能力的新技术和新产品。
萎锈灵,化学名称为5,6−二氢−2−甲基−N−苯基−1,4−氧硫杂环己烯−3−甲酰胺,具有氧硫杂环己二烯酰胺类结构,是琥珀酸脱氢酶抑制剂类(Succinate dehydrogenase inhibitors,SDHIs)杀菌剂,在植株体内具有内吸传导作用,其作用机制主要是抑制病原菌琥珀酸脱氢酶活性,从而干扰其呼吸作用,阻碍能量代谢,抑制病原菌的生长,导致其死亡,从而达到防治病害的目的[6]。已有研究表明,萎锈灵包衣处理对麦类锈病、棉花立枯病等有较好防效[7−9],而且能促进棉花生长并具有抗氧化防御机制[10],还能增强田间棉花幼苗耐低温冷害胁迫的能力[11]。但是,关于萎锈灵对棉种萌发的影响不清楚,尤其是萎锈灵对低温胁迫下棉种萌发特性的影响尚待进一步探讨。本研究将萎锈灵按照不同含量制成悬浮种衣剂包衣棉种,以对温度敏感的棉花品种新陆早50[12]为研究对象,比较不同含量萎锈灵对低温环境下棉种萌发和干物质转移相关指标的影响,以期筛选出萎锈灵提高棉苗耐低温的适宜剂量,为在生产中利用萎锈灵提高棉花幼苗耐寒性提供理论依据。
1 材料与方法
1.1 实验材料
棉花品种:新陆早50。2018年收获,种子含水率等基础性状符合国家标准GB4407.1−2008(经济作物种子第1部分:纤维类),由新疆农业科学院经济作物研究所提供。
药剂:(1)99%萎锈灵原药。(2)基础种衣剂料浆:只包含成膜剂、表面活性剂等种衣剂惰性成分(非活性助剂成分),不包含杀菌剂农药活性成分。(3)系列含量萎锈灵:在基础种衣剂基础上添加4.2%、5.7%、7.2%、8.7%和10.2%含量的萎锈灵。(2)和(3)均由新疆农业科学院核技术生物技术研究所自行研制。
1.2 实验设计
实验于2019年在新疆农业科学院核技术生物技术研究所化控室进行,采用双因素随机区组设计方法。选择籽粒饱满的种子经1%次氯酸钠消毒10min,灭菌蒸馏水冲洗干净并晾干备用。以光籽不包衣处理为对照,萎锈灵处理设置0(种衣剂中仅含惰性成分)、4.2%、5.7%、7.2%、8.7%和10.2%共6个梯度含量的萎锈灵按药种重量比1:100进行包衣,依次记为CK1(未包衣)、CK2(不含萎锈灵包衣)、S1(4.2%萎锈灵包衣)、S2(5.7%萎锈灵包衣)、S3(7.2%萎锈灵包衣)、S4(8.7%萎锈灵包衣)、S5(10.2%萎锈灵包衣);温度处理参照高利英等[13]方法,以25℃为常温对照,设置3个胁迫温度:18℃、15℃、12℃,昼夜恒温,光周期为12h/12h(光/暗)。实验共设置28个处理,每处理3次重复。
1.3 种子萌发
采用砂培法进行种子萌发实验。经水洗净的细砂干热灭菌后回温到室温下,添加13%的灭菌水混匀,装入发芽盒(规格20cm×16cm×12cm)中构建发芽床。每个发芽盒沙床上等距离摆放不同处理的50粒种子;每个处理重复摆放4盒,共计200粒种子;置于特定温度的室内人工智能气候箱进行种子萌发。实验过程中,从第4天开始每48h观察记录种子萌发情况,25℃和18℃温度下萌发实验中,苗高达4cm及以上视为正常发芽[14],15℃和12℃低温萌发实验中,以芽长超过0.5cm,颜色为白色、无腐烂的芽为正常发芽[15]。
1.4 项目观测
从种子萌发实验开始,14d后种子结束萌发。25℃处理下,棉花种子经过5~7d就完成发芽,后续快速生长,发芽盒盖将被顶开,可采用更高的盒子罩住,以防止水分蒸发。从各处理中均随机选出10株棉苗,洗净后,经105℃杀青30min,80℃烘干至质量恒定,用万分之一电子分析天平称量发芽前种子干重、发芽后的残留物(种皮和种壳)干重以及幼苗鲜/干质量。
1.5 指标计算
参照高荣岐等[14]的方法计算发芽势、发芽率、发芽指数以及活力指数等种子萌发活性指标;参照施成晓等[16]的方法计算全苗干物质质量、种子干物质转化效率以及呼吸消耗量等干物质转移指标。
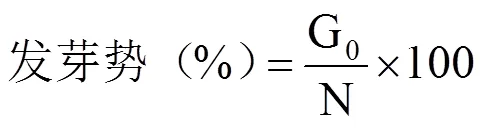

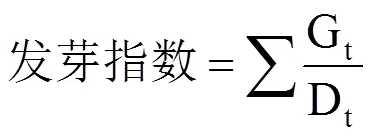

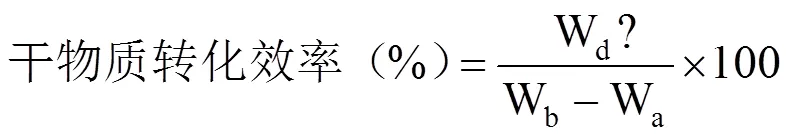
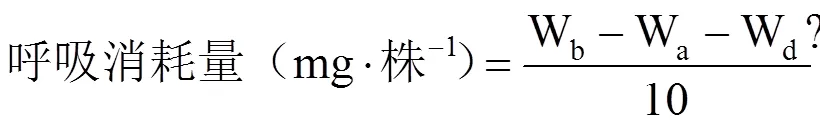
式中,G0为25℃第4日或18℃~12℃第6日统计的发芽种子数,G为正常发芽种子数,N为种子总数,Gt为第t日统计的发芽数,Dt为Gt相对应的种子出苗经历的时间(d);Wb为发芽前随机选的10粒各处理种子干重(g),Wa为发芽12d后随机选取的10株幼苗的种子残留物干重(g),Wd为12d后随机选取的10株幼苗的干物质质量(g)。
采用仪小梅等[17]方法计算各处理种子的发芽势、发芽率等8个指标的抗逆系数(Adversity resistance coefficient,AC)和种子的平均抗逆系数(即一个处理下8个指标抗逆系数的平均值),对7个不同处理进行抗逆分析,判定其耐低温冷害的能力。抗逆系数是评价抗逆境胁迫能力的指标,平均抗逆系数越大,说明种子耐低温冷害能力越强,反之则越弱。
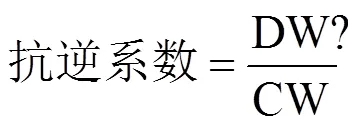

式中,DW为低温胁迫下各处理种子萌发活性和干物质转移指标值,CW为25℃条件下种子萌发活性和干物质转移指标值,N为种子发芽势等8个指标数目即等于8。
1.6 数据处理
采用Microsoft Excel 2010进行数据分析,用GraphPad Prism v5.0进行双因素方差分析和作图,基于发芽势、发芽率等8个指标采用IBM SPSS 20.0软件将7个处理进行聚类分析。
2 结果与分析
2.1 萎锈灵包衣对低温胁迫下棉种萌发进程的影响
图1a显示,正常温度条件下(25℃),实验第4天观测时,各处理种子均已萌发,且发芽率均超过60.0%,其中以CK1(未包衣)和CK2(不含萎锈灵包衣)的发芽率最低,均为64.0%,不同含量萎锈灵包衣的各处理即S1(4.2%萎锈灵包衣)、S2(5.7%萎锈灵包衣)、S3(7.2%萎锈灵包衣)、S4(8.7%萎锈灵包衣)、S5(10.2%萎锈灵包衣)的发芽率均超过73.0%,且除S5外萎锈灵含量越高棉种发芽率越高。从整个14d种子萌发进程看,与CK1和CK2比较,以萎锈灵包衣的各处理棉种的萌发进程提早了1~2d,发芽率提高了5.0~16.3个百分点。可见,在正常温度条件下,萎锈灵包衣可促进种子萌发,加快萌发进程并提高发芽率。
图1b显示,在18℃温度条件下,CK1(未包衣)和CK2(不含萎锈灵包衣)处理中,棉种均在第6天开始萌发,比正常温度下(25℃)推迟了2d,发芽高峰期也推迟2d,最高发芽率降低5.0个百分点。与CK1和CK2相比,不同含量萎锈灵包衣的各处理即S1、S2、S3、S4、S5的发芽率均超过了60.0%,S3和S4处理棉种的发芽高峰期到来最早,均比CK1提早2d,最大发芽率提高了10.0~14.2个百分点。
图1c、d显示,当温度继续降至15℃和12℃时,未包衣(CK1)和不含萎锈灵包衣(CK2)处理中,棉种在第6~8天以后才开始萌发,比正常温度下(25℃)推迟4~6d,发芽高峰期也推迟2~4d到达,发芽率均比正常温度下降低15.5~65.2个百分点。与CK1和CK2相比,15℃和12℃时不同含量萎锈灵包衣的各处理即S1、S2、S3、S4、S5的最高发芽率分别为63.5%~76.3%和7.5%~13.5%,萎锈灵包衣的棉种发芽高峰期也未提早。
由此表明,随着温度降低,未含萎锈灵包衣的棉种发芽率均随之降低,发芽时间延长2~6d,含萎锈灵包衣的棉种发芽率亦均随温度降低而降低,发芽时间延长1~6d;低温条件下,萎锈灵包衣能提高种子活性,促进种子萌发,加快种子的萌发进程。
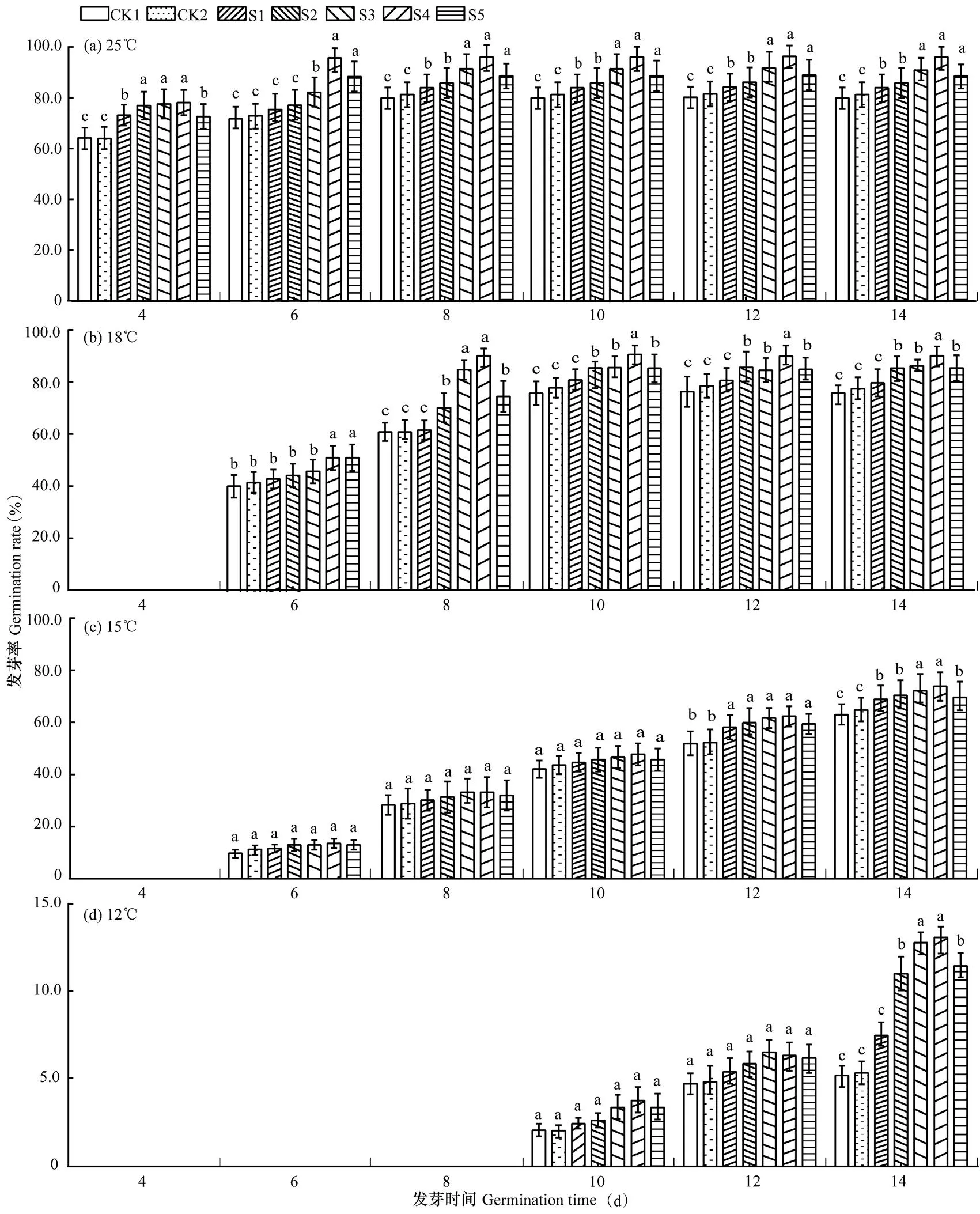
图1 不同温度条件下各包衣处理棉种发芽进程的比较
注:短线表示标准差,小写字母表示各处理在0.05水平上的差异显著性。CK1:未包衣,CK2:不含萎锈灵包衣,S1:4.2%萎锈灵包衣,S2:5.7%萎锈灵包衣:S3:7.2%萎锈灵包衣,S4:8.7%萎锈灵包衣,S5:10.2%萎锈灵包衣;温度处理以25℃为常温对照,设置3个胁迫温度:18℃、15℃、12℃。下同。
Note:The bar is mean standard deviation. Lowercase letters indicate the significant difference among treatments at 0.05 level. CK1:uncoated.CK2:coated with no carboxin. S1:coated with 4.2% carboxin. S2: coated with 5.7% carboxin. S3:coated with 7.2% carboxin. S4:coated with 8.7% carboxin. S5:coated with 10.2% carboxin. 25℃ as normal temperature control, three stress temperatures: 18℃, 15℃, 12℃. The same as below.
2.2 萎锈灵包衣对低温胁迫下棉种萌发活性的影响
由图2可见,在不同低温胁迫条件下,随着温度降低,各处理棉种萌发活性指标均呈下降趋势,25℃时各处理棉种萌发活性最高,18℃和15℃其次,12℃下最低,在相同温度条件下,CK1(未包衣)和CK2(不含萎锈灵包衣)处理棉种的萌发活性各指标数值均最小,随着萎锈灵含量的增加,种子萌发活性各指标值均增加。图2b显示,S4处理(8.7%萎锈灵包衣)棉种发芽率受影响最大,25℃、18℃、15℃和12℃下发芽率分别为96.0%、90.0%、76.0%和13.5%,分别较CK1处理增加16.3、14.5、12.1和8.2个百分点;S3处理(7.2%萎锈灵包衣)棉种发芽指数受影响最大,25℃、18℃、15℃和12℃下发芽指数分别为11.4、6.8、4.5和0.5,分别较CK1处理增加17.5%、12.5%、7.1%和5.5%(图2c);S4处理(8.7%萎锈灵)棉种活力指数受影响最大,25℃、18℃、15℃和12℃下活力指数分别为790.8、579.5、529.5和420.0,分别较CK1处理增加37.6%、44.0%、46.7%和55.0%(图2d)。由此表明,随着温度降低,不含萎锈灵包衣的棉种萌发活性指标均随之降低,含萎锈灵包衣的棉种萌发活性指标也均随之降低,但是经8.7%萎锈灵包衣棉种活力指数较CK1处理增加37.6%~55.0%,相应的发芽率也提高6.3~13.1个百分点。说明低温条件下,萎锈灵包衣后能提高种子活性,从而提高棉种发芽率。
棉种萌发活性相关指标的双因素方差分析结果表明(表1),萎锈灵包衣和温度对棉花种子的发芽势、发芽率、发芽指数和活力指数的贡献度不同,从萎锈灵包衣和温度所占总平方和的百分比来看,温度对发芽率、发芽势、发芽指数和活力指数的贡献度较高,分别为96.0%、98.0%、95.5%和94.6%;萎锈灵包衣对发芽率、发芽势、发芽指数和活力指数的贡献度为其次,分别为1.5%、0.9%、0.6%和2.9%。由此表明,温度是影响棉种萌发的主要因素,萎锈灵包衣是次要因素。
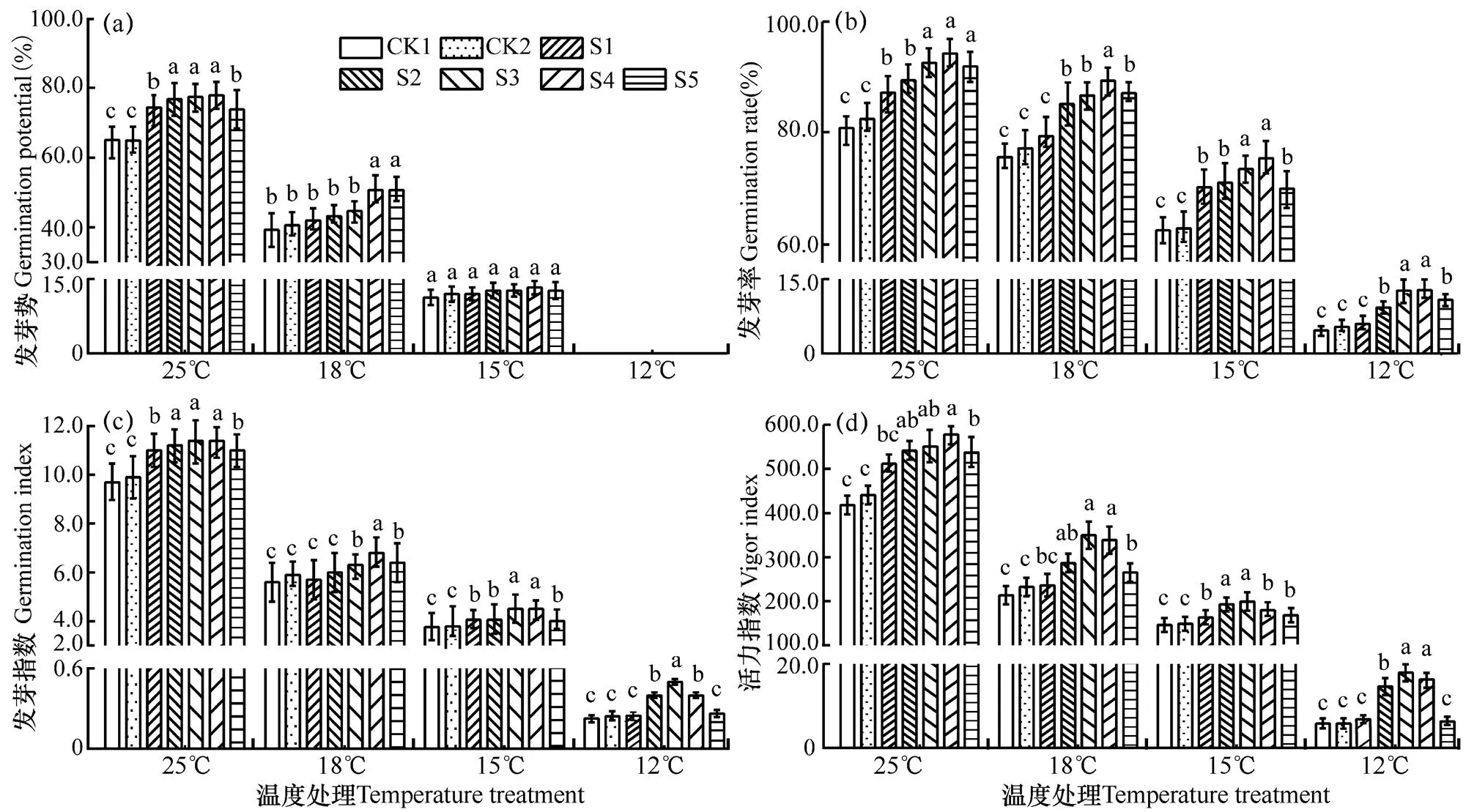
图2 同一温度条件下各处理棉种萌发活性相关指标的比较
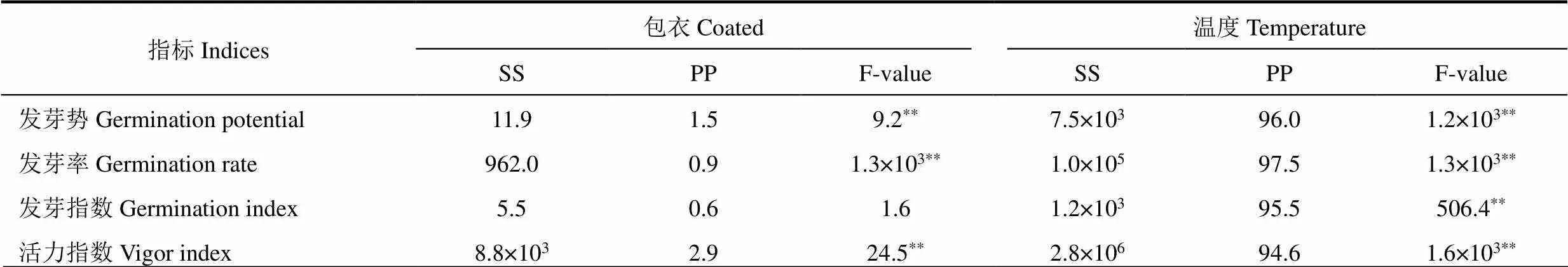
表1 棉种萌发活性相关指标的双因素方差分析
注:SS表示平方和,PP表示占处理变异平方和的百分比,*和**分别表示相关系数通过0.05、0.01水平的显著性检验。下同。
Note:SS indicates sum of squares. PP indicates percentage of the value to the total sum of squares.*is P< 0.05,**is P<0.01.The same as below.
2.3 萎锈灵包衣对低温胁迫下棉种干物质转移的影响
各包衣处理对低温胁迫下棉种干物质转移的变化如图3。由图可见,随着温度降低,各处理的棉种干物质转移各个指标均呈下降趋势,在温度相同的条件下,CK1(未包衣)和CK2(不含萎锈灵包衣)处理棉种的萌发活性各指标中除呼吸消耗最大外,其它指标均最小,随着萎锈灵含量的增加,除种子的呼吸消耗量呈先下降后上升的趋势外,干物质转移其它相关指标均呈先上升后下降的趋势(图2)。其中,S4处理(8.7%萎锈灵包衣)棉花幼苗鲜重受影响最大,25℃、18℃、15℃和12℃下分别较CK1增加19.9%、21.6%、23.2%和31.0%(图3a);图3b显示, S3处理(7.2%萎锈灵包衣)棉幼苗干重受影响最大,各温度下分别较CK1增加22.0%、30.3%、35.3%和39.9%;图3c显示,S4处理棉种干物质转化率受影响最大,各温度下分别较CK1增加22.4、30.5、31.3和40.0个百分点;图3d表明,S4处理棉种呼吸消耗量受影响最大,各温度下分别较CK1减少74.0%、73.8%、57.0%和42.5%。由此表明,除呼吸消耗量呈上升趋势外,含萎锈灵包衣和不含萎锈灵包衣的棉种其余干物质转移指标均随温度降低而降低,但是含萎锈灵包衣的种子干物质转化率较CK1最高时提高了40.0个百分点,相应的幼苗干物质积累较CK1也增加了25.0%。说明低温条件下,萎锈灵包衣能增强种子活性,提高种子干物质转化效率,增加棉花幼苗干物质积累,更有利于幼苗生长发育和适应低温环境。
棉种干物质转移相关指标的双因素方差分析结果表明(表2),萎锈灵包衣和温度对棉花种子的鲜苗质量、干苗质量、干物质转化效率和呼吸消耗量的贡献度不同,从萎锈灵包衣和温度所占总平方和的百分比来看,温度对鲜苗质量、干苗质量、干物质转化效率和呼吸消耗量的贡献度较高,分别为90.9%、44.0%、45.9%和60.1%;萎锈灵包衣对鲜苗质量、干苗质量、干物质转化效率和呼吸消耗量的贡献度分别为6.4%、23.1%、24.4%和31.5%。由此表明,温度是影响棉种干物质转移的主要因素,萎锈灵包衣为次要因素。

图3 同一温度条件下各处理棉种干物质转移相关指标的比较
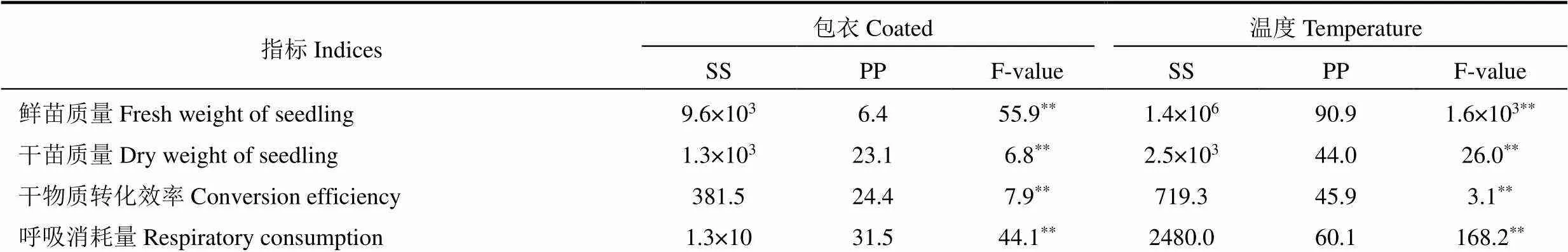
表2 棉种干物质转移相关指标的双因素方差分析

2.4 萎锈灵包衣对低温胁迫下棉种抗逆性的影响
各包衣处理棉种萌发活性和干物质转移各指标的抗逆系数如表3。由表可见,随着温度的降低,CK1(未包衣)和CK2(不含萎锈灵包衣)处理棉种萌发活性和干物质转移各指标的抗逆系数均呈下降趋势,18℃时棉种各个指标抗逆系数最大,15℃其次,12℃最小。相同温度条件下,CK1(未包衣)和CK2(不含萎锈灵包衣)处理棉种各指标中除呼吸消耗量抗逆系数差异显著外,其它指标的抗逆系数差异均不显著;与CK1和CK2相比,随着萎锈灵含量的增加,各指标抗逆系数均呈先上升后下降的趋势。其中,CK1和CK2的平均抗逆系数分别为0.68和0.70,分别位居第七和第六,表现出耐低温能力相对较弱;与CK1(未包衣)和CK2(不含萎锈灵包衣)相比,随着萎锈灵含量的增加,萎锈灵包衣棉种的平均抗逆系数呈先上升后降低的趋势,表现出一定的耐低温能力;S3(7.2%萎锈灵包衣)处理的平均抗逆系数值最高,为0.91,位居第一,其次是S4(8.7%萎锈灵包衣)处理的平均抗逆系数值,为0.88,位居第二,S1(4.2%萎锈灵包衣)处理的平均抗逆系数值最低,为0.73,位居第五。由此表明,在低温胁迫条件下,不含萎锈灵包衣的棉种其发芽势等8个指标的平均抗逆系数相对较低,表现出耐低温能力相对较弱;而含萎锈灵包衣的棉种其发芽势等8个指标的平均抗逆系数相对高,表现出一定的耐低温能力。

表3 各低温条件下处理间各项指标抗逆系数(与25℃条件下相比)的比较(平均值±标准差)
注:抗逆系数指各低温条件下处理间各项指标与25℃条件下处理间对应指标的比值。
Note: The adversity resistance coefficient refers to the ratio of each index at low temperature and the corresponding index at 25℃.
为了综合评价不同处理的棉种耐低温萌发的抗逆能力,将各包衣处理棉种的发芽势等8个指标的抗逆系数进行聚类分析,结果如图4。由图可见,各处理大致可分为3类,即CK1(未包衣)、CK2(未含萎锈灵包衣)和S1(4.2%萎锈灵包衣)聚为一类,种子萌发过程中耐低温能力相对较弱;S3(7.2%萎锈灵包衣)和S4(8.7%萎锈灵包衣)聚为一类,种子萌发时具有相对较强的抗低温能力;S2(5.7%萎锈灵包衣)和S5(10.2%萎锈灵包衣)为一类,种子萌发时具有一定的抗低温能力。由此表明,在低温胁迫条件下,未包衣和不含萎锈灵包衣的棉种活性相对低,耐低温能力相对较弱;而含萎锈灵包衣的棉种活性相对较高,受低温胁迫抑制较小,表现出一定的耐低温能力,其中S3(7.2%萎锈灵包衣)处理提高棉花耐低温胁迫的能力最佳,S4(8.7%萎锈灵包衣)处理次之。
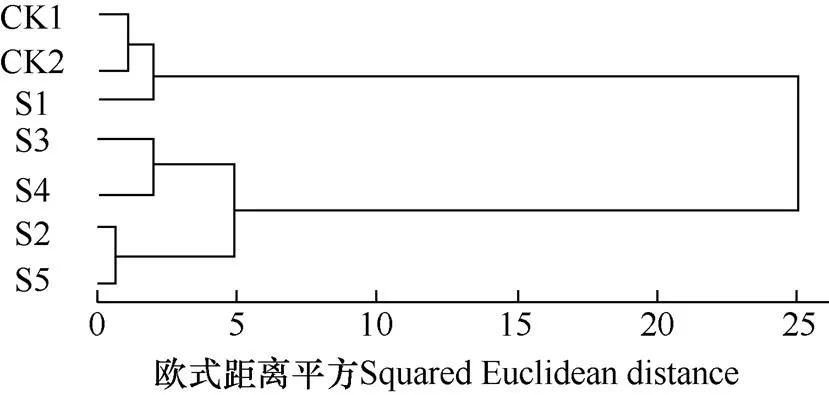
图4 棉种抗逆系数聚类分析
3 讨论与结论
3.1 讨论
温度是影响植物种子萌发的主要因素之一。已有研究表明,棉花种子萌发的最适温度为28℃左右,最低温度为12~10.5℃,而在10℃以下,棉花种子不萌发,25℃是棉花种子萌发最适温度的下限,在大多耐低温冷害的研究报道中以此温度作为对照[13]。本研究表明,在25℃条件下,不同处理的棉种发芽率虽有差异,但均在80%以上;随着温度降低,各个包衣处理的棉种发芽进程呈下降趋势,下降幅度存在显著差异,与未包衣和不含萎锈灵处理比较,萎锈灵处理明显提高了棉种活力,缩短种子发芽进程2~4d。由此表明,萎锈灵处理明显提高了棉种活力,加快萌发进程并提高萌发率。
种子萌发期的发芽势、发芽率、发芽指数和活力指数指标可反映种子萌发期耐低温胁迫的强弱,对种子萌发起着关键作用[12−13]。已有研究表明,低温胁迫下棉花种子的萌发和幼苗生长受到抑制,水杨酸以及外源调节剂复配包衣处理均能显著提高低温胁迫下棉花种子的发芽势、发芽率、发芽指数和活力指数,其中发芽指数和活力指数增幅分别为41.2%~44.4%和51.2%~63.9%[18]。本研究表明,在低温胁迫下,含萎锈灵包衣棉种的发芽率、发芽势、发芽指数和活力指数等萌发指标均高于不含萎锈灵处理,说明不含萎锈灵处理的棉花种子的萌发活性已受到抑制,而含萎锈灵包衣增强了种子活力,提高了棉种的发芽率、发芽势、发芽指数和活力指数等萌发指标。由此表明,萎锈灵处理可缓解低温胁迫对棉种萌发造成的伤害,提高棉芽的耐冷性。
种子在萌发过程中,需要转化和利用自身贮备的营养物质来提供生长所需的物质和能量,对种子萌发期耐低温胁迫起重要作用[16,19]。杨小环等研究表明,在低温胁迫下外源水杨酸处理玉米种子的幼苗干重和鲜重较对照分别提高15.5%和28.5%[20]。本研究表明,与未包衣和不含萎锈灵处理相比较,低温胁迫下萎锈灵处理后,棉花幼苗鲜苗质量、干苗质量、干物质转化效率显著增强而呼吸消耗量显著降低。由此表明,低温胁迫条件下萎锈灵处理能提高棉种活性,促进种子干物质的转移和棉花幼苗干物质积累,有利于幼苗生长发育和适应低温环境,进而提高棉种耐冷性。
种子萌发期低温耐受性是多种抗逆性状的综合表现,用单一指标评价棉花的耐低温胁迫能力具有片面性,不能全面反映其抗逆能力,采用多指标抗逆系数的平均值作为耐低温萌发能力的评价值,可消除处理间基础性状不同的影响,该数值越大,表明该处理对提高棉花耐低温萌发能力越强[21]。本研究表明,与未包衣和不含萎锈灵处理相比较,含萎锈灵包衣后的平均抗逆系数值均增加,其中,含萎锈灵包衣的平均抗逆系数介于0.73~0.91,而未包衣和不含萎锈灵处理分别为0.68和0.70,说明含萎锈灵包衣的耐低温冷害能力大于不含萎锈灵处理,未包衣和不含萎锈灵处理的棉种更容易受到低温胁迫的影响。聚类分析表明,未包衣、不含萎锈灵处理和4.2%萎锈灵处理聚为一类,种子萌发过程中耐低温能力相对较弱;7.2%和8.7%萎锈灵处理聚为一类,种子萌发时具有相对较强的抗低温能力;5.7%和10.2%萎锈灵处理聚为一类,种子萌发时具有一定的抗低温能力。这进一步表明,萎锈灵处理提高了种子活力,种子萌发期低温耐受性也得到提高。
3.2 结论
在低温胁迫下,与未包衣和不含萎锈灵处理比较,萎锈灵处理可增强棉花种子的活力,提高棉花萌发期的耐低温能力,进而提早萌发进程,提高发芽率,促进幼苗生长,但是不同含量的萎锈灵处理的结果也存在差异,7.2%萎锈灵处理效果最好,8.7%萎锈灵处理其次。
[1]Gu S,Peng H,Zheng P Y,et al.Climate change favours a destructive agricultural pest in temperate regions:late spring cold matters[J].Journal of Pest Science,2018,91(4):1191-1199.
[2] 王森,王雪蛟,吉春容,等.Cotton XL模型模拟研究延迟型低温冷害对棉花纤维品质的影响[J].农业工程学报,2019,35 (20):171-177.
Wang S,Wang X J,Ji C R,et al.Impact of delayed-type chilling damage on cotton fiber quality based on Cotton XL[J]. Transactions of the CSAE,2019,35(20):171-177.(in Chinese)
[3] 李防洲,辛慧慧,周广威,等.水杨酸包衣剂包衣棉种对棉花幼苗抗寒性的影响[J].棉花学报,2015,27(6):589-594. Li F Z,Xin H H,Zhou G W,et al.Effects of the seedlings seed film coating salicylic acid on the chilling tolerance of cotton [J].Cotton Science,2015,27(6):589-594.(in Chinese)
[4] 雷斌,李进,胡鹏,等.18.6%拌·福·乙种衣剂提高棉花耐低温胁迫能力验证[J].农药,2017,56(4):288-292. Lei B,Li J,Hu P,et al.Ability of amicarthiazol·thiram·acephate 18.6% seed coating agent to enhance the tolerance of cotton to low temperature stress[J].Agrochemicals,2017,56(4):288- 292.(in Chinese)
[5] 雷斌,李进,段留生,等.种衣剂对低温处理下棉花胚根及幼苗外部形态和超微结构的影响[[J]中国农业气象,2017,38 (4):248-256. Lei B,Li J,Duan L S,et al.Effect of seed coating agents on external morphology and ultrastructure of cotton radicles and seedlings under low temperature treatments[J].Chinese Journal of Agrometeorology,2017,38(4):248-256.(in Chinese)
[6] Ragsdale N N,Sisler H D.Metabolic effects related to fungitoxicity of carboxin[J].Phytopathology,1970,60(10): 1422-1429.
[7] Lisker N,Meiri A.Control of Rhizoctonia solani damping-off in cotton by seed treatment with fungicides[J].Crop Protection, 1992,11(2):155-159.
[8]师勇强,冯自力,李志芳,等.400g·L-1福美双·萎锈灵悬浮种衣剂对棉花苗期立枯病的防治效果[J].中国棉花,2014,41(9):28-30. Shi Y Q,Feng Z L,Li Z F,et al.Control of cotton seedlingdisease by 400g·L-1thiram·carboxin suspending formulations for seed coating[J].Journal of China Cotton,2014,41(9):28-30.(in Chinese)
[9]黄青春,周明国,叶钟音.拌种灵、叶枯唑和萎锈灵对病菌的毒力机制比较[J].农药学学报,2000,2(4):72-75. Huang Q C,Zhou M G,Ye Z Y.Analogy of seedvax,saikuzuo and carboxin in virulence mechanism[J].Chinese Journal of Pesticide Science,2000,2(4):72-75.(in Chinese)
[10] Dêmily A L,Caroline M,Alan C C,et al.Changes in antioxidants potential,secondary metabolites and plant hormones induced by different fungicides treatment in cotton plants[J].Pesticide Biochemistry and Physiology, 142:117-122.
[11] 周小云,李进,张军高,等.萎锈灵杀菌剂对提高棉花耐低温冷害胁迫能力的影响[J].新疆农业科学,2020,57(4): 705-712.Zhou X Y,Li J,Zhang J G,et al.Effects of the carbox in fungicideon the chilling stress of cotton seedlings[J]. Xinjiang Agricultural Sciences,2020,57(4):705-712.(in Chinese)
[12] 徐建伟,张晨,曾晓燕,等.近十年新疆北疆主栽棉花种子低温萌发能力差异评价[J].新疆农业科学,2017,54(9):1569- 1578.Xu J W,Zhang C,Zeng X Y,et al.Evaluation of seed germination of main-cultivated cotton under low temperature in northern Xinjiang in recent ten years[J]. Xinjiang Agricultural Sciences,2017,54(9):1569-1578.(in Chinese)
[13] 高利英,邓永胜,韩宗福,等.黄淮棉区棉花品种种子萌发期低温耐受性评价[J].棉花学报,2018,30(6):455-463. Gao L Y,Deng Y S,Han Z F,et al.Evaluation of the low-temperature region during seed germination tolerance of cotton varieties in the Huang-Huai[J].Cotton Science,2018, 30(6):455-463.(in Chinese)
[14] 高荣岐,张春庆.种子生物学[M].北京:中国农业出版社,2009. Gao R Q,Zhang C Q.Seed biology[M].Beijing:China Agriculture Press,2009.(in Chinese)
[15] 王钰静,谢磊,李志博,等.低温胁迫对北疆棉花种子萌发的影响及其耐冷性差异评价[J].种子,2014,33(5):74-77. Wang Y J,Xie L,Li Z B,et al.Effects of low temperature stress to germination of cotton seeds and evaluation of their cold resistance in Northern Xinjiang[J].Seed,2014,33(5): 74-77.(in Chinese)
[16]施成晓,陈婷,王昌江,等.干旱胁迫对不同抗旱性小麦种子萌发及幼苗根芽生物量分配的影响[J].麦类作物学报,2016,36(4):483-490. Shi C X,Chen T,Wang C J,et al.Effect of drought stress on seed germination and biomass allocation of root and shoot of different drought resistant wheat cultivars[J].Journal of Triticeae Crops,2016,36(4):483-490.(in Chinese)
[17]仪小梅,孙爱清,韩晓玉,等.黄淮麦区小麦主推品种(系)干热风抗性鉴定[J].麦类作物学报,2015,35(2):274-284. Yi X M,Sun A Q,Han X Y,et al.Identification of dry-hot wind resistance of major wheat cultivars(strains) in Huanghuai wheat region[J].Journal of Triticeae Crops,2015,35(2):274-284.(in Chinese)
[18] 李防洲,冶军,侯振安.外源调节剂包衣对低温胁迫下棉花种子萌发及幼苗耐寒性的影响[J].干旱地区农业研究,2017,35(1):192-197. Li F Z,Ye J,Hou Z A.Effects of seed film coating with exogenous regulating substances on cotton germination rate and cold tolerance of seedlings[J].Agricultural Research in the Arid Areas,2017,35(1):192-197.(in Chinese)
[19]王志恒,邹芳,杨秀柳,等.PEG-6000模拟干旱对春小麦种子萌发的影响[J].种子,2019,38(7):12-17. Wang Z H,Zou F,Yang X L,et al.Effects of PEG-6000 on seed uermination of spring wheat under simulated drought conditions[J].Seed,2019,38(7):12-17.(in Chinese)
[20] 杨小环,赵维峰,孙娜娜,等.外源水杨酸缓解低温胁迫对玉米种子萌发和早期幼苗生长伤害的生理机制[J].核农学报,2017,31(9):1811-1817. Yang X H,Zhao W F,Sun N N,et al.Physiological mechanisms of exogenous salicylic acid-mediated low temperature tolerance in seed germination and early seedling growth of maize[J].Joumal of Nuclear Agricultural Sciences, 2017,31(9):1811-1817.(in Chinese)
[21] 胡树平,苏治军,于晓芳,等.玉米自交系抗旱相关性状的主成分分析与模糊聚类[J].干旱地区农业研究,2016,34(6): 81-88. Hu S P,Su Z J,Yu X F,et al.Principal component analysis and fuzzy clustering on drought tolerance related traits of maize inbred lines[J].Agricultural Research in the Arid Arereas, 2016,34(6):81-88.(in Chinese)
Effects of the Carboxin from Seed Coating Formulation on the Cotton Seed Germination Characteristics under Low Temperature Stress
ZHOU Xiao-yun1,LIU Meng-li2,LI Jin1,ZHANG Jun-gao1,LIANG Jing1,DU Peng-cheng2,WANG Li2,ZAI Meng-hua2,GAN En-xiang2,GAO Wen-wei2,LEI Bin1
(1.Research Institute of Nuclear Technology and Biotechnology, Xinjiang Academy of Agricultural Sciences/Key Laboratory of Crop Ecophysiology and Farming System in Desert Oasis Ministry of Agriculture,Urumqi 830091,China; 2.College of Agronomy,Xinjiang Agricultural University,Urumqi 830052)
Xinjiang, as the largest cotton producing area in China, often suffer from low temperature meteorological disasters such as “late spring cold” in cultivation, which leads to different degrees of rotten seeds, rotten buds and dead seedlings every year, resulting in “difficult emergence and seedling protection”, affecting cotton growth and development and yield and quality formation. With the development of seed coating technology, the above problems of “difficult emergence and seedling protection” in cotton field in normal years have been effectively solved by coating cotton seeds with seed coating formulation. However, it is still difficult to ensure a high rate of seedling conservation in the field in the year with serious low temperature damage, so it is urgent to study new technologies and products to improve the low temperature tolerance of cotton seedling. In addition, carboxin is a succinate dehydrogenase inhibitor fungicide, which has the function of internal absorption and conduction in the plant. Its mechanism is mainly to inhibit the activity of succinate dehydrogenase of pathogenic bacteria, result to interfere with its respiration, hinder energy metabolism, inhibit the growth of pathogenic bacteria, and cause its death, so as to achieve the purpose of disease control. Recent studies have shown that the treatment with carboxin coating plays a major role in control on cotton seedling damping-off disease, and can promote the growth of cotton, and can also enhance the ability of the field cotton seedlings to tolerante cold stress. But, it is not clear about the effect of carboxin on the germination of cotton seeds, especially the effect of carboxin on the germination characteristics of cotton seeds under low temperature stress. Therefore, in order to reduce the influence of low temperature stress on the germination ability of cotton seeds, the effects of the carboxin from seed coating formulation on the cotton seed germination characteristics under low temperature stress was explored.
In this study, ‘Xinluzao 50’ cotton seeds coated with six gradients of carboxin, which were 0 (inert component only), 4.2%, 5.7%, 7.2%, 8.7% and 10.2%, were sown in plastic boxes with sand and were exposed to four temperature treatments at 25℃, 18℃, 15℃ and 12℃.The uncoated cotton seeds exposed to the same temperature treatments served as the control group (CK). Sand culture was adopted to study the effect of carboxin on the germination and dry matter transfer of cotton seeds under low temperature stress to explore the effect of carboxin on improving the cold resistance of cotton. The results showed that the germination index of cotton seed uncoated was the highest under normal temperature (25℃), and the germination rate was 80.5%. The germination index of cotton seed uncoated decreased with the decrease of temperature, which indicated that the germination index of cotton seed was seriously affected by low temperature stress, while the germination index of cotton seed coated with carboxin was the highest under normal temperature (25℃), and the germination rate was 96.0%. With the decrease of temperature, the germination indices of all treatments with coating with carboxin decreased slowly, which indicated that carboxin could obviously alleviate the damage of low temperature stress on seed germination.Under the same temperature, with the increase of the content of carboxin in seed coating agent, the other indices increased first and then decreased, except the respiratory consumption of seeds decreased first and then increased. The germination process of cotton seeds coating with carboxin was 2−4 days earlier than that in the treatment of seed uncoated and seed coating without carboxin; the seed vigor and growth of the treatment without carboxin was lower, and the fresh weight was significantly reduced, resulting in the dry matter accumulation was also significantly reduced; the average adversity resistance coefficient of the treatment uncoated cotton seeds and without carboxin coated cotton seeds was only 0.68 and 0.70, respectively, while that of the treatment with carboxin was 0.73−0.91. Our results showed that carboxin could enhance the seed vigor, advance the germination process, increase the germination rate and promote the growth of seedlings, and enhance the ability of seedlings to resist damage from the low-temperature climate during the germination period of cotton.
Carboxin; Germination rate; Germination index; Vigor index; Adversity resistance coefficient; Dry matter conversion efficiency; Cotton; Seed germination; Low temperature stress.
10.3969/j.issn.1000-6362.2020.08.003
周小云,刘梦丽,李进,等.萎锈灵包衣对低温胁迫下棉种萌发特性的影响[J].中国农业气象,2020,41(8):495-505
2020−02−20
雷斌,E-mail:leib668@xaas.ac.cn
国家自然科学基金“萎锈灵杀菌剂对提高新疆棉花苗期耐低温冷害能力的机制研究”(31760073)
联系方式:周小云,E-mail:xiaoyunzhou77@126.com
猜你喜欢
今日农业(2021年10期)2021-11-27
军事文摘(2021年16期)2021-11-05
诗潮(2021年1期)2021-03-15
今日农业(2020年23期)2020-12-15
上海农业科技(2020年2期)2020-04-02
山西农业科学(2020年1期)2020-02-27
劳动保护(2018年8期)2018-09-12
农民致富之友(2018年9期)2018-06-27
山东工业技术(2016年15期)2016-12-01
中国火炬(2015年12期)2015-07-31
