
落叶型大丽轮枝菌脱落酸合成相关基因功能研究
2020-08-27 03:55王娟菲李海源商文静胡小平
西北农业学报 2020年8期
王娟菲,李海源,秦 君,商文静,胡小平
(农业部西北黄土高原作物有害生物综合治理重点实验室,西北农林科技大学 植物保护学院,陕西杨凌 712100)
棉花黄萎病是典型的土传性真菌病害,病原菌从棉花根部入侵,越过皮层到达维管束后,随水分的输送向上传播[1],继而引起叶片的黄化萎蔫和脱落,严重时导致植株死亡。1966年Schnathorst从抗病性显著下降的抗黄萎病棉株中分离出两株黄萎病致病菌系,按其对3个鉴别品种 ‘海岛棉’‘岱字棉’和‘爱字棉’上的反应型划分为两个类型:非落叶型SS-4菌系和落叶型T9菌系,T9菌系的毒力比SS-4的强10倍[2]。与非落叶型菌株相比,落叶型菌株具有引致棉花发病重、发病快、损失严重等特点[3-7]。
ABA(Absicisic acid)是调节植物生长发育以及非生物和生物胁迫反应的关键植物激素[8]。在植物响应外界环境压力、调节气孔开放、调控叶片脱落、种子萌发与休眠等非生物胁迫中起重要的调节作用[9-12]。同时,ABA也在病原菌侵染和寄主免疫过程发挥作用,当ABA含量增加两倍,番茄灰霉病的发病率可降低50%[13]。在稻瘟菌与ABA互作中,ABA在免疫反应中起负向调控作用,与SA信号通路相拮抗参与植物免疫过程[14]。研究表明,病原菌可以通过内源产生低水平ABA参与调控植物的免疫反应,如蔷薇色尾孢霉菌Cercosporarosicola,菜豆尾孢霉菌Cercosporacruenta和灰葡萄孢霉菌Botrytiscinerea[15-16]。bcaba1是灰葡萄孢霉菌上第一个被发现的ABA合成相关基因[17],后续研究发现bcaba2、bcaba3和bcaba4基因也参与灰葡萄孢霉菌ABA的合成[18]。目前,多数研究主要集中在干旱、高盐、低温等非生物胁迫下ABA的产生以及对植物生理生长等的影响[12,19-22]。但尚未见到关于ABA与大丽轮枝菌落叶型菌株引起棉花落叶关系的研究报道。
本文通过NCBI比对灰葡萄孢霉菌ABA合成相关基因,得到落叶型大丽轮枝菌XJ592中ABA合成相关基因Vdaba1、Vdaba2(1)、Vdaba2(2)和Vdaba4,对4个基因进行克隆和敲除,比较分析其产孢量、菌落形态、压力响应、致病性、落叶情况等,了解这些基因对大丽轮枝菌的生物学特性及致病性的影响。
1 材料与方法
1.1 菌株和质粒
野生型大丽轮枝菌XJ592菌株(落叶型菌株)和根癌农杆菌菌株EHA105由西北农林科技大学土传病害实验室保存。大肠杆菌DH5α感受态细胞购自康为世纪公司。敲除质粒pOSCAR和带有潮霉素筛选标记的质粒pA-Hyg-OSCAR均由Fungal Genetics Stock Center(www.fgsc.net)提供。
1.2 ABA合成基因生物信息学分析
在NCBI(National Centre of BiotechnologyInformation)网站搜索灰葡萄孢霉菌ABA合成基因bcaba1、bcaba2、bcaba3和bcaba4的蛋白质序列信息,使用Blastp程序进行大丽轮枝菌中同源蛋白的搜索和比对分析。
1.3 敲除载体构建及敲除突变体的获得
参照Pazet的报道,设计同源臂扩增引物用于扩增目的基因Vdaba1、Vdaba2(1)、Vdaba2(2)和Vdaba4的侧翼序列,构建基因敲除载体[23]。通过PCR和酶切验证后,将阳性敲除载体导入农杆菌菌株EHA105感受态细胞中,进行农杆菌介导的遗传转化。根据转化子能扩增出潮霉素抗性基因Hyg但不能扩增出目的基因的原则,得到候选敲除转化子。将具有潮霉素B(50 μg/mL)抗性的转化子进行单孢纯化。采用PCR检测筛选阳性转化子,用于后续试验。
1.4 产孢量测定
配制浓度为1×106个/mL的敲除突变体菌株孢子悬浮液。吸取100 μL孢子悬浮液,接种至装有50 mL Czapek液体培养基的三角瓶中,于25 ℃、120 r/min条件下黑暗摇培7 d后,在显微镜下用血球计数板测量孢子悬浮液浓度,以野生型XJ592菌株为对照,重复3次。
1.5 压力敏感性测定
在PDA上培养7 d的野生型、敲除突变体菌落边缘打直径为5 mm的菌饼,分别接种到CM培养基、含有0.03% H2O2(氧化压力)、0.01% SDS(细胞壁压力)、1.2 mol/L山梨醇(渗透压力)、0.7 mol/L NaCl(盐胁迫)、0.2 g/L刚果红(细胞壁胁迫)的CM平板上,25 ℃黑暗培养15 d后观察菌落特征,用“十字”交叉法测量菌落直径。每个处理接种3个培养皿,重复3次。
1.6 致病力测定
接种方法参照棉花孢子悬浮液浸根接种法[24]。待棉花幼苗长到2片真叶平展时,将棉苗小心取出,用清水清洗干净,浸在孢子浓度为106个/mL的悬浮液中30 min,以无菌水作为空白对照。接种21 d后,统计发病率,计算病情指数。接菌后30 d统计棉花的落叶数量和落叶率。病情指数=[∑(各级病株数×相应病级)/(调查总株数×4)]×100[25]。
1.7 数据分析的方法
产孢量采用SAS v8.01软件的TTEST过程进行t测验(P=0.05)分析,野生型菌株和敲除突变体菌株的菌落直径、压力筛选及病情指数等的方差分析采用ANOVA过程进行。
2 结果与分析
2.1 ABA合成基因的生物信息学分析
基于灰葡萄孢霉菌的ABA合成相关基因bcaba1(AJ609392)、bcaba2(AJ851088)、bcaba3(AM237449)和bcaba4(AM237450)[11]蛋白序列,与大丽轮枝菌基因组序列经Blastp比对共得到4个同源基因,分别命名为Vdaba1、Vdaba2(1)、Vdaba2(2)、Vdaba4(表1)。

表1 基因基本信息Table 1 Basic information of genes
2.2 敲除载体构建及突变体筛选
用CTAB法提取大丽轮枝菌XJ592基因组DNA,使用Vdaba1、Vdaba2(1)、Vdaba2(2)和Vdaba4基因的上下游引物分别扩增其侧翼序列,采用OSCAR一步法构建基因敲除载体[15],转化大肠杆菌,用潮霉素抗性基因检测引物Hyg-F和Hyg-R检测阳性菌落并送擎科生物公司测序,将测序正确的菌落接种于LB培养基摇培,提取质粒。采用农杆菌介导的遗传转化方法,将敲除质粒转入野生型菌株XJ592中,经过50 mg/L潮霉素抗性筛选,并使用潮霉素基因检测引物、目的基因检测引物对敲除转化子菌株进行PCR验证,共得到8个突变体菌株(图1)。选取△Vdaba1-20、△Vdaba1-21、△Vdaba2(1)-48、△Vdaba2(1)-36、△Vdaba2(2)-20、△Vdaba2(2)-11、△Vdaba4-12、△Vdaba4-11等8个代表性突变体进行下一步研究。
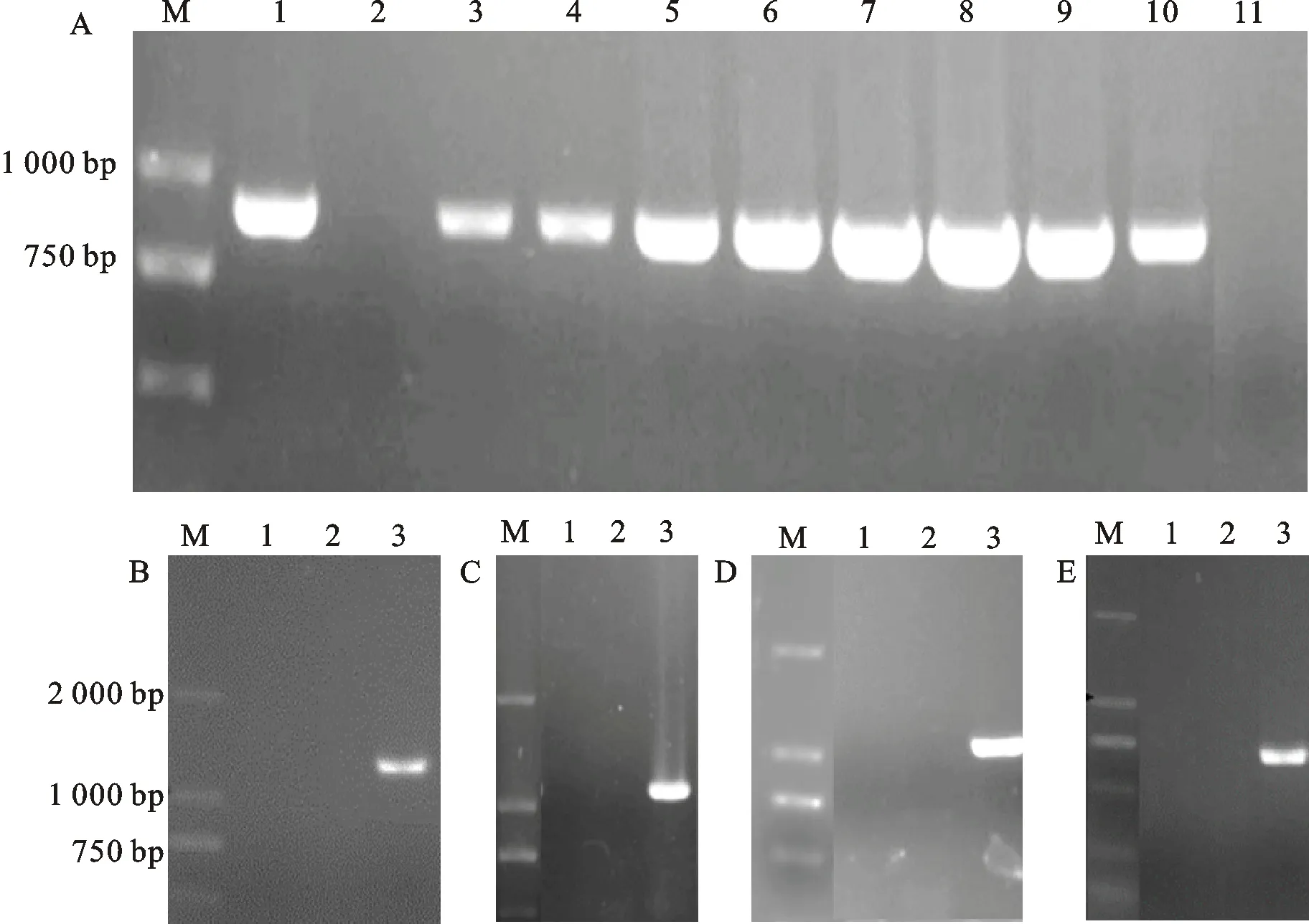
A:潮霉素引物PCR电泳图。M:DL2000Maker;泳道1:pA-Hyg-OSCAR质粒;泳道2:野生型XJ592;泳道3-10:敲除突变体 △Vdaba1-20、 △Vdaba1-21、 △Vdaba2(1)-48、 △Vdaba2(1)-36、 △Vdaba2(2)-20、 △Vdaba2(2)-11、 △Vdaba4-12、 △Vdaba4-11;泳道11:阴性对照。B、C、D、E:目的基因检测结果电泳图。M:DL2000Maker;B图,泳道1-2:敲除突变体 △Vdaba1-20、 △Vdaba1-21;泳道3:野生型XJ592;C图,泳道1-2:敲除突变体 △Vdaba2(1)-36、 △Vdaba2(1)-48;泳道3:野生型XJ592;D图,泳道1-2:敲除突变体 △Vdaba2(2)-11、 △Vdaba2(2)-20;泳道3:野生型XJ592;E图, 泳道1-2:敲除突变体 △Vdaba4-12、 △Vdaba4-11;泳道3:野生型XJ592。
2.3 产孢量测定
将野生型菌株和突变体菌株接种于液体查氏培养基中,120 r/min室温摇培,7d后测孢子浓度。结果表明,突变体△Vdaba2(1)-48、△Vdaba2(1)-36、△Vdaba2(2)-20、△Vdaba2(2)-11、△Vdaba4-12和△Vdaba4-11产孢量与野生型的无显著性差异,而突变体△Vdaba1-20和△Vdaba1-21产孢量显著少于野生型的(图2),说明Vdaba1参与大丽轮枝菌孢子的形成,Vdaba2(1)、Vdaba2(2)和Vdaba4对大丽轮枝菌的产孢过程没有显著作用。
2.4 菌落形态观察及压力筛选
将野生型XJ592和4个基因共8个突变体接种于CM培养基上,25 ℃黑暗培养15 d,运用十字交叉法测量菌落直径,结果发现,突变体△Vdaba2(1)-48、△Vdaba2(1)-36、△Vdaba2(2)-20和△Vdaba2(2)-11菌落直径显著小于野生型的,突变体△Vdaba4-12、△Vdaba4-11、△Vdaba1-20和△Vdaba1-21菌落直径与野生型无显著差异。在含有20 μg/mL SDS培养基上黑暗培养15 d后,突变体△Vdaba2(1)-48、△Vdaba2(1)-36、△Vdaba4-12、和△Vdaba4-11菌落直径显著高于野生型XJ592的(图3)。在 0.03% H2O2压力胁迫下,突变体△Vdaba4-12和△Vdaba4-11菌落直径显著受到抑制,△Vdaba1-20、△Vdaba1-21、△Vdaba2(1)-48和△Vdaba2(1)-36突变体菌落直径显著增大。可见Vdaba4基因的存在可以使大丽轮枝菌更能适应氧化环境。在1.2 mol/L山梨醇压力下,所有突变体菌落直径均显著大于野生型的,说明ABA合成相关基因可负向调节大丽轮枝菌对渗透压的敏感性。在含有0.7 mol/L NaCl的培养基上,只有△Vdaba2(2)突变体与野生型直径无显著差异,其余突变体菌落直径均显著大于野生型的,说明Vdaba1、Vdaba2(1)、Vdaba4的存在使大丽轮枝菌对盐胁迫更敏感。在0.2 g/L刚果红压力胁迫中,只有△Vdaba2(1)菌落直径显著减小。
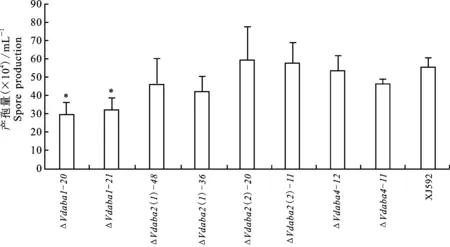
“*”表示P=0.05下具有显著性差异 “*”Significant difference at P=0.05

*表示经单因素方差分析P=0.05水平具有显著性 *Significant difference at P=0.05
2.5 致病力测定
野生型XJ592和基因突变体菌株接种‘冀棉11’,21 d后,接种野生型和突变体的棉花植株都表现出典型的黄萎病症状(图4)。按照棉花黄萎病苗期病害调查分级标准统计发病情况,突变体△Vdaba1-20、△Vdaba1-21、△Vdaba4-12和△Vdaba4-11菌株接菌棉花植株的病情指数低于野生型菌株的,而突变体△Vdaba2(1)-48、△Vdaba2(1)-36、△Vdaba2(2)-20和△Vdaba2(2)-11的病情指数与野生型无显著差异(图5)。接种30 d后,所有突变体都可以引起整株棉花叶片全部脱落,与野生型XJ592菌株接菌植株无差异(数据略)。

图4 野生型及突变体菌株接种棉花21 d后发病情况Fig.4 Disease symptoms of cotton at 21 days post inoculation of XJ592 and mutants

“*”表示经单因素方差分析P=0.05具有显著性差异 “*”Significant difference at P=0.05
3 讨 论
通过对大丽轮枝菌侵染寄主引发寄主落叶或叶片萎蔫而不脱落的表型可将大丽轮枝菌分为落叶型和非落叶型[2],落叶性状是大丽轮枝菌种群演化的重要类型。在中国,随着陆家云首次报道落叶型大丽轮枝菌之后,在全国主产棉区也陆续开展了落叶型大丽轮枝菌的相关研究。吴献忠等[26]对来自山东的16株大丽轮枝菌的致病力研究将其分为强致病力(与落叶型相似)、中等致病力和弱致病力,并发现山东省有落叶型菌系存在。落叶型大丽轮枝菌因其发病快、致病力强近年来逐渐成为棉花黄萎病的优势种群[27-30]。自从其被发现以来,科研工作者一直致力于探讨落叶型大丽轮枝菌引起寄主落叶的分子机理。通过随机多态性扩增(RAPD)确定落叶型菌株有一段长为 0.55 kb的特异序列,这为分子标记在黄萎病流行病学方面提供了新思路[31]。同时,对大丽轮枝菌致病基因挖掘与功能鉴定。基因组功能分析等方面开展相关研究,进一步推动了大丽轮枝菌病源学、流行病学、病原与寄主互作机制研究的发展[32-34]。Zhang等[35]通过比较基因组学鉴定出高致病力的落叶型菌株有一段LSR(lineage-specific regions)区编码的7个VdDfs基因,该区域在低致病非落叶型菌株基因组中基本完全缺失。通过对这7个基因结构域分析表明 VdDfs基因簇为一个典型的次生代谢物合成基因簇,该基因簇产生的次级代谢物N-酰基乙醇胺(NAEs)[36]。研究表明(NAEs)可以与ABA相互作用发挥生物学功能[37]。植物落叶是一个自然而复杂的生理过程,涉及生理、生化和分子网络等多个调控系统[38-41]。有研究表明,在水压力胁迫和一些不良环境胁迫下,ABA的产生会显著上调,从而可以调节叶片脱落[42-43]。一直以来认为一些低等病原物不能合成ABA,直到最近研究发现病原菌侵染后植物叶片ABA水平会上升,主要原因是病原菌自身合成ABA的水平提高所致。虽然病原菌只合成少量的ABA,但可以触发植物对外界环境的生理反应[44]。当灰葡萄孢霉菌侵染番茄后,番茄叶片的ABA是未接菌处理的10倍多。研究者将ABA的合成分为4步:侵染时寄主诱导病原菌ABA合成,接着病原菌释放ABA或者ABA前体物,此时植物体内的ABA产量逐渐积累,同时病原菌自身有抑制ABA代谢。相比只接菌,ABA和病原菌孢子共同处理后番茄叶片会更快的出现坏死现象[45]。因此植物的抗病程度与ABA合成相关。在拟南芥中,ABA合成相关基因aba2-1突变体对镰刀菌的抗性显著增强[46]。而aba-1突变体对疫霉菌和丁香假单胞菌的抗性明显减弱,植物叶片枯黄,脱落[47]。然而目前有关ABA合成与落叶型大丽轮枝菌引起棉花落叶之间关系的研究尚未见报道。本研究中,大丽轮枝菌ABA合成相关基因Vdaba1、Vdaba2(1)、Vdaba2(2)和Vdaba4不影响大丽轮枝菌XJ592引起的棉花落叶,但影响大丽轮枝菌的产孢能力、菌落生长及致病力等,说明ABA合成相关基因在病原菌引起的落叶过程中并不发挥功能,而对大丽轮枝菌的生理生长与致病过程起到至关重要的作用。可能ABA在生物因素引起的落叶过程和非生物胁迫引起的落叶过程中发挥不同的生物功能。
大丽轮枝菌ABA合成基因Vdaba1的敲除对大丽轮枝菌的产孢量有显著的抑制作用,说明Vdaba1参与大丽轮枝菌的产孢过程,Vdaba1敲除突变体菌株的病情指数下降,可能是由于孢子产量减少导致的。本研究△Vdaba4突变体接菌棉花后,致病力减弱,说明Vdaba4也参与大丽轮枝菌致病过程。同时发现△Vdaba4突变体菌株对H2O2压力更敏感,说明Vdaba4基因在大丽轮枝菌抵抗氧化压力中起促进作用。这可能是△Vdaba4突变体导致大丽轮枝菌致病性减弱的原因之一。基因Vdaba2(1)和Vdaba2(2)对大丽轮枝菌的致病性没有影响,却对大丽轮枝菌的营养生长起正向调控作用。因此大丽轮枝菌ABA合成基因Vdaba1、Vdaba2(1)、Vdaba2(2)和Vdaba4在调节大丽轮枝菌的生长和致病性方面行使不同的功能。前人研究表明,病原菌侵染植物寄主时,可以产生许多次级代谢物,而且真菌通常是以基因簇的形式参与次级代谢物的形成[48]。病原菌ABA的产出通过远距离的运输与转移,参与病原菌致病侵染过程[12]。
ABA在响应各种不良环境中发挥重要作用,如在各种非生物胁迫干旱、水等不适条件中,ABA的产生可以帮助植物度过不良环境[38-39]。但在大丽轮枝菌引起的棉花落叶方面,ABA合成相关基因敲除没有发挥决定作用。一方面本研究中通过NCBI比对灰葡萄孢霉菌ABA合成相关基因,并不是落叶型大丽轮枝菌ABA合成所特有的基因,不同真菌ABA合成过程可能存在差异。另一方面在这个研究中只做了单基因敲除,大丽轮枝菌ABA合成过程可能存在基因冗余的情况,致使突变体接菌棉花也表现出落叶表型,需要进一步开展双敲除、三敲除甚至四敲除突变体研究,测试他们对棉花落叶性状的影响。
猜你喜欢
亚热带农业研究(2022年1期)2022-08-08
现代食品(2022年6期)2022-04-19
复旦学报(医学版)(2021年4期)2021-08-05
农业科技通讯(2021年1期)2021-03-06
农业工程学报(2020年20期)2020-12-25
数学大王·低年级(2020年8期)2020-08-14
中国农业科技导报(2020年3期)2020-03-15
山西农业科学(2020年2期)2020-02-29
小雪花·初中高分作文(2016年9期)2016-05-14
中国质量万里行(2015年1期)2015-01-27
