
转录组解析外源ABA对玉米脱水速率的影响
2020-08-29 04:10张美微张盼盼刘京宝乔江方
华北农学报 2020年4期
李 川,黄 璐,张美微,张盼盼,刘京宝,牛 军,乔江方
(河南省农业科学院 粮食作物研究所,河南 郑州 450002)
玉米是我国重要的粮食、饲料、经济三元作物。2018年我国玉米播种面积4 200万hm2,总产量2.573亿t。随着农村劳动力转移及我国农业机械化水平的提高[1-2],生产上对适宜机械化收获的玉米品种需求愈来愈高,尤其是具备籽粒脱水速率快、早熟高产、抗倒性好等特性的机械粒收品种[3-5]。目前,玉米脱水速率相关文献多从植株性状、生理特征方面进行研究。有文献表明,玉米苞叶层数、厚度及松紧程度影响籽粒脱水速率[6-8]。玉米生理成熟时籽粒含水量与散粉时间、轴粗、百粒质量、穗位高、根倒伏呈显著正相关[9],而果柄短、籽粒小有助于籽粒脱水。总之,田间自然脱水速率与植株性状相关,低株高、高穗位、开花期较多绿叶数、高茎秆含水率、低叶片含水率、穗粗直径小、穗行数少、籽粒宽度大、苞叶薄、百粒质量小有利于籽粒脱水[10-12]。玉米籽粒含水量与籽粒破碎率、杂质率、落籽率呈显著正相关[13]。不同玉米品种脱水速率不同。白色胚乳晚熟玉米脱水速率较慢,不同熟期玉米杂交种籽粒脱水速率存在显著差异[14]。
植物激素在作物生长发育过程起着重要的调控作用,脱落酸(ABA)尤为重要[15-16]。灌浆早期喷施外源ABA降低灌浆速率,并与ABA浓度呈正相关[17];灌浆中期高浓度喷施外源ABA降低灌浆速率[18-19]。灌浆速率是影响玉米粒质量的主要限制因素,进而影响百粒质量和产量[20-21]。高浓度ABA破坏胚乳细胞结构,抑制胚乳细胞分裂从而减少胚乳细胞及淀粉粒数目[22-24]。玉米胚中ABA含量变化与脱水耐性相关[25]。外源ABA加快种子胚脱水速率[26]。但外源ABA影响玉米脱水速率的分子机制尚未明确。鉴于此,在灌浆早期分别对脱水速率较快的迪卡517和脱水速率较慢的郑单1002穗部喷施外源ABA,然后利用高通量转录组测序技术挖掘相关差异表达基因并注释其功能,分析其代谢途径和转录因子,以期找到ABA影响玉米脱水速率的重要响应基因及调控途径。
1 材料和方法
1.1 试验材料
供试玉米材料为迪卡517(DK517)、郑单1002(ZD1002)。其中,DK517由中种国际种子有限公司选育,并于2017年通过国家农作物品种审定委员会审定,审定编号:国审玉20170005,该品种生理成熟后籽粒脱水速度快叶片干枯,有利于籽粒直接收获;ZD1002由河南省农业科学院粮食作物研究所选育,于2015年通过国家农作物品种审定委员会审定,审定编号:国审玉2015017,该品种高产优质、抗病性强,适合黄淮海地区夏播,该品种籽粒脱水速率一般,在籽粒乳线消失之后适时收获可以增加产量。
1.2 试验设计
试验材料于2018年6月18日播种于河南省农业科学院研究与示范试验基地(原阳),该地属暖温带大陆性季风气候。供试土壤为潮土,地势平坦,肥力均匀,排灌方便,试验田前茬作物为小麦,常规田间管理。0~30 cm耕层土壤有机质含量17.25 g/kg、全氮含量0.96 g/kg、速效氮含量 79.35 mg/kg、有效磷含量10.22 mg/kg、速效钾含量94.56 mg/kg。
DK517和ZD1002的种植密度均为67 500株/hm2,60 cm等行距种植,每个处理3个重复,共12个小区。8月3-4日抽雄,在吐丝之前,每小区标记生长一致、无病虫害的代表性植株并进行雌穗套袋,8月5-6日授粉,8月9日算作灌浆启动。用0.08‰外源ABA(ABA80)(0.5%(V/V)Tween-20作为展开剂)进行喷施,分别于8月6,14,16日(均为晴天)16:00-17:00对穗位叶和穗部进行喷施;对照 (CK)套袋授粉后正常生长。
1.3 取样材料
灌浆14 d后(8月22日)开始取样,每个样本由5株长势较为一致的玉米穗位叶混合组成,生物学重复3次,命名为:DK518-ABA80-1、DK518-ABA80-2、DK518-ABA80-3;DK518-CK-1、DK518-CK-2、DK518-CK-3;ZD1002-ABA80-1、ZD1002-ABA80-2、ZD1002-ABA80-3;ZD1002-CK-1、ZD1002-CK-2、ZD1002-CK-3。取样后立即放于液氮中,然后置于-80 ℃超低温冰箱保存。
1.4 转录组测序
按照RNeasy plant mini kit(Qiagen,Germany) 试剂盒操作步骤提取上述样本总RNA。用1%琼脂糖凝胶电泳检验总RNA的完整性,用Agilent Bioanalyzer 2100 system(Aglilent Technologies,USA)生物芯片分析检测仪检测所提RNA的质量,并用Nanodrop ND-1000 UV-Vis Spectrophotometer (Nanodrop Technologie,USA) 定量检测所提RNA的浓度。将检测合格的RNA 样本送至北京百迈客生物科技有限公司构建cDNA文库,然后用Illumina HiSeqTM2500测序仪进行高通量测序,结果序列为pair-end序列。
1.5 基因功能注释
通过去除接头序列、低质量序列、多N序列及长度过短序列,对测序原始序列进行质量控制得到干净序列,并计算测序碱基错误率及GC碱基含量。利用Hisat 2序列比对软件,采用BWT算法将所得干净序列与参考基因组B73序列(https://www.maizegdb.org/)进行比对。利用RSeQC-2.6.3软件对比对所得基因进行测序饱和度、基因覆盖度及冗余序列分析后,参照蛋白质序列数据库Nr (NCBI non-redundant protein sequences)、核酸序列数据库Nt (NCBI non-redundant nucleotide sequence)、蛋白质家族集合数据库Pfam (Protein family)、真核生物蛋白相邻类聚簇数据库KOG/COG (Clusters of orthologous groups of proteins)、蛋白质序列详细信息数据库Swiss-Prot (A manually annotated and reviewed protein sequence database)、京都基因组数据库KO (KEGG orthology database)、基因本体论联合会建立的基因相关信息数据库GO(Gene ontology)等公共数据库进行基因功能注释。
1.6 筛选差异表达基因
在转录组测序分析中用R(F)PKM值(每1 000 000条序列中每个基因以1 000个碱基为单位,比对上的序列数量)衡量基因表达水平。依据基因数量并用edgeR软件计算差异表达基因(P<0.05)。显著差异表达基因筛选标准为基因错误发现率FDR<0.05 &基因表达量差异倍数的对数值|log2FC|≥2。之后参照GO数据库,将差异基因按照生物学过程(Biological process)、分子功能(Molecular function)、细胞组分(Cellular component)三大类对基因功能进行进一步详细的分类。参照KEGG ( Kyoto encyclopedia of genes and genomes) 数据库分析差异表达基因参与的代谢途经。
2 结果与分析
2.1 转录组测序结果统计
12个样本全转录组高通量测序共获得90.71 Gb干净序列,各样品干净序列均达到6.09 Gb,碱基质量值(Q30)在91.41%及以上,分别将各样品的干净序列与参考基因组B73序列进行比对,比对效率为84.69%~90.06%。基于序列比对结果,进行可变剪接预测分析、基因结构优化分析以及新基因的发掘,发掘新基因7 998个,其中6 444个得到功能注释。
表1详细统计了12个样品的测序结果。DK517-ABA80-1原始测序经过质量剪切过滤后获得了49 659 930 条干净序列,GC含量为55.02%,没有AT/GC分离现象,Q30值达91.75%;利用Hisat2序列比对软件与参照B73基因组比对,88.42%的序列配对成功,获得43 908 042条配对序列。
DK517-CK-1原始测序经过质量剪切过滤后获得了49 337 744条干净序列,GC含量为55.95%,没有AT/GC分离现象,Q30值达92.66%;利用Hisat2序列比对软件与参照B73基因组比对,88.68%的序列配对成功,获得43 754 859条配对序列。其余样本的测序结果详见表1。由表1可见,测序结果比较可靠。

表1 12个外源ABA处理穗位叶样本转录组测序质量统计表Tab.1 Statistics of transcriptome data of 12 ear-samples treated by ABA
2.2 外源ABA处理相关差异表达基因统计
外源ABA处理后,将不同样本中表达水平存在显著差异的基因命名为差异表达基因(DEG)。DEG可以划分为上调基因(Up-regulated gene)和下调基因(Down-regulated gene)。用R(F) PKM衡量比对到的基因表达量。12个测序样本分成3个对比组进行分析,用A VS B表示,由图1可见,DK517外源ABA处理材料与正常生长条件材料对比组(DK517-ABA80 VS DK517-CK,下同)差异表达基因共检测到1 732个DEG,其中1 251个为上调表达基因,481个为下调表达基因;DK517中上调表达基因个数占总差异表达基因个数的72.23% 。另外,检测到36 490个表达差异不显著的基因。ZD1002-ABA80 VS ZD1002-CK差异表达基因检测到52个DEG,其中24个为上调表达基因,28个为下调表达基因;ZD1002中上调表达基因占总差异表达基因的46.15%。另外,检测到38 917个表达差异不显著的基因。DK517-ABA80 VS ZD1002-ABA80差异表达基因共检测到3 867个DEG,其中1 946个为上调表达基因,1 921个为下调表达基因;另外,检测到36 265个表达差异不显著的基因。
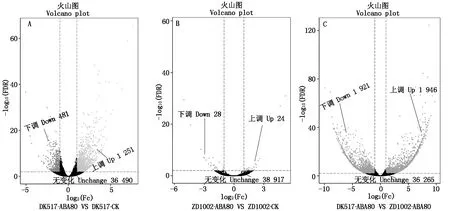
每个圆点表示1个基因,横坐标表示某个基因在两样品中表达量差异倍数的对数值;横坐标绝对值越大,说明表达量在两样品间的表达量倍数差异越大;纵坐标表示基因表达量变化的统计学显著性的负对数值。纵坐标值越大,表明差异表达越显著,筛选得到的差异表达基因越可靠。
2.3 外源ABA处理相关差异表达基因类别分析
参照GO数据库结构化的标准生物学注释系统,按照生物学过程、分子功能、细胞组分3个主要分支统计3个测序样本组中DEG的GO分类情况。DK517-ABA80 VS DK517-CK差异表达基因主要富集于细胞凋零、生物节奏过程、单有机体过程、多有机体过程、繁殖过程、复制、多细胞有机体过程、发育过程、细胞成分组织或生物合成、生物调控、刺激反应、细胞过程、代谢过程、定位、信号、免疫系统过程、生长、生物阶段、脱毒19个不同生物学过程分类;细胞成分、细胞、细胞器、细胞膜、细胞器成分、大分子复合体、细胞外区成分、细胞膜成分、细胞连接、超大分子复合体、细胞外区、膜封闭内腔12个不同细胞组分分类;结合、催化活性、运输活性、核酸结合转录因子活性、结构分子活性、电子载体活性、分子功能调节因子、信号传递活性、抗氧化活性、分子传递活性、营养储备活性、转录因子活性及蛋白质结合12个不同分子功能分类。ZD1002-ABA80 VS ZD1002-CK差异表达基因主要富集于单有机体过程、多细胞有机体过程、繁殖过程、复制、多有机体过程、发育过程、细胞成分组织或生物合成、生物调控、刺激反应、细胞过程、代谢过程、定位、生长13个不同生物学过程分类;细胞成分、细胞、细胞器、细胞膜、细胞器成分、大分子复合体、细胞外区成分、细胞膜成分、细胞连接、膜封闭内腔10个不同细胞组分分类;结合、催化活性、电子载体活性、营养储备活性4个不同分子功能分类。DK517-ABA80 VS ZD1002-ABA80差异表达基因主要富集于单有机体过程、运动、多有机体过程、繁殖过程、复制、多细胞有机体过程、发育过程、细胞成分组织或生物合成、生物调控、刺激反应、细胞过程、代谢过程、定位、信号、免疫系统过程、生长、生物阶段、脱毒18个不同生物学过程分类;细胞成分、细胞、细胞器、细胞膜、细胞器成分、大分子复合体、细胞外区成分、细胞膜成分、细胞连接、超大分子复合体、细胞外区、膜封闭内腔12个不同细胞组分分类;结合、催化活性、运输活性、核酸结合转录因子活性、结构分子活性、电子载体活性、分子功能调节因子、信号传递活性、抗氧化活性、分子传递活性、营养储备活性、转录因子活性及蛋白质结合12个不同分子功能分类。
参照COG数据库对检测到的差异表达基因产物进行直系同源分类。DK517-ABA80 VS DK517-CK差异表达基因主要分布于染色体结构及动态变化过程、能量产生及储存、细胞循环控制细胞分裂及染色体分区、氨基酸转运及代谢、核苷酸转运及代谢、碳水化合物转运及代谢、辅酶转运及代谢、脂类转运及代谢、翻译核糖体结构及生物合成、转录、复制修复及重组、细胞壁细胞膜及被膜生物合成、细胞运动、翻译后修饰蛋白质中转及分子伴侣、无机离子运输及代谢、次级代谢物生物合成运输及分解代谢、通用功能预测、未知功能、信号传导机制、胞内运输分泌及膜泡运输、防卫机制、细胞骨架、细胞外结构23个不同COG数据库分类。ZD1002-ABA80 VS ZD1002-CK差异表达基因主要分布于细胞循环控制细胞分裂及染色体分区、氨基酸转运及代谢、翻译核糖体结构及生物合成、细胞壁细胞膜及被膜生物合成、翻译后修饰蛋白质中转及分子伴侣、次级代谢物生物合成运输及分解代谢、通用功能预测、防卫机制8个不同COG数据库分类。DK517-ABA80 VS ZD1002-ABA差异表达基因的COG数据库分类与DK517-ABA80 VS DK517-CK相似,仅少了1个染色体结构及动态COG数据库分类,但同一COG数据库分类中富集到的差异表达基因数目不同。
生物体内不同基因产物往往相互协调而行使生物学功能。因此,对差异表达基因的代谢途径(Pathway)进行注释分析可进一步了解基因功能。KEGG数据库是系统分析基因功能及基因组信息的数据库,把基因和表达信息作为一个整体网络进行研究。KEGG数据库包括碳水化合物、核苷酸、氨基酸等代谢及有机物的生物降解。图2-4展示了3个对比组差异表达基因KEGG注释结果通路类型分类。DK517-ABA80 VS DK517-CK差异表达基因注释到100个不同代谢通路中,其中富集到的差异表达基因≥10的代谢通路主要有植物激素信号传导、内质网中蛋白质加工过程、DNA复制、同源重组、不匹配修复、核苷酸切除修复、核糖体、真核细胞中核糖体合成、天冬氨酸及谷氨酸代谢、半胱氨酸及蛋氨酸代谢、缬氨酸亮氨酸及异亮氨酸降解、果糖及甘露糖代谢、糖酵解/糖异生、丙酮酸代谢、淀粉及蔗糖代谢、光合器官中碳固定、光合作用、氨基酸生物合成、碳水化合物代谢、谷胱甘肽代谢、嘌呤代谢、嘧啶代谢(图2)。其中,尤其值得注意的是植物激素信号传导系统通路,共富集22个差异表达基因,如表2所示。

纵坐标为KEGG代谢途径名称;横坐标为注释到该通路下的基因个数及其个数占被注释上的基因总数的比例。图3-4同。The ordinate is KEGG metabolic pathway;The abscissa is gene numbers annotated to the pathway and the proportion of gene numbers annotated to the total number of genes annotated.The same as Fig.3-4.
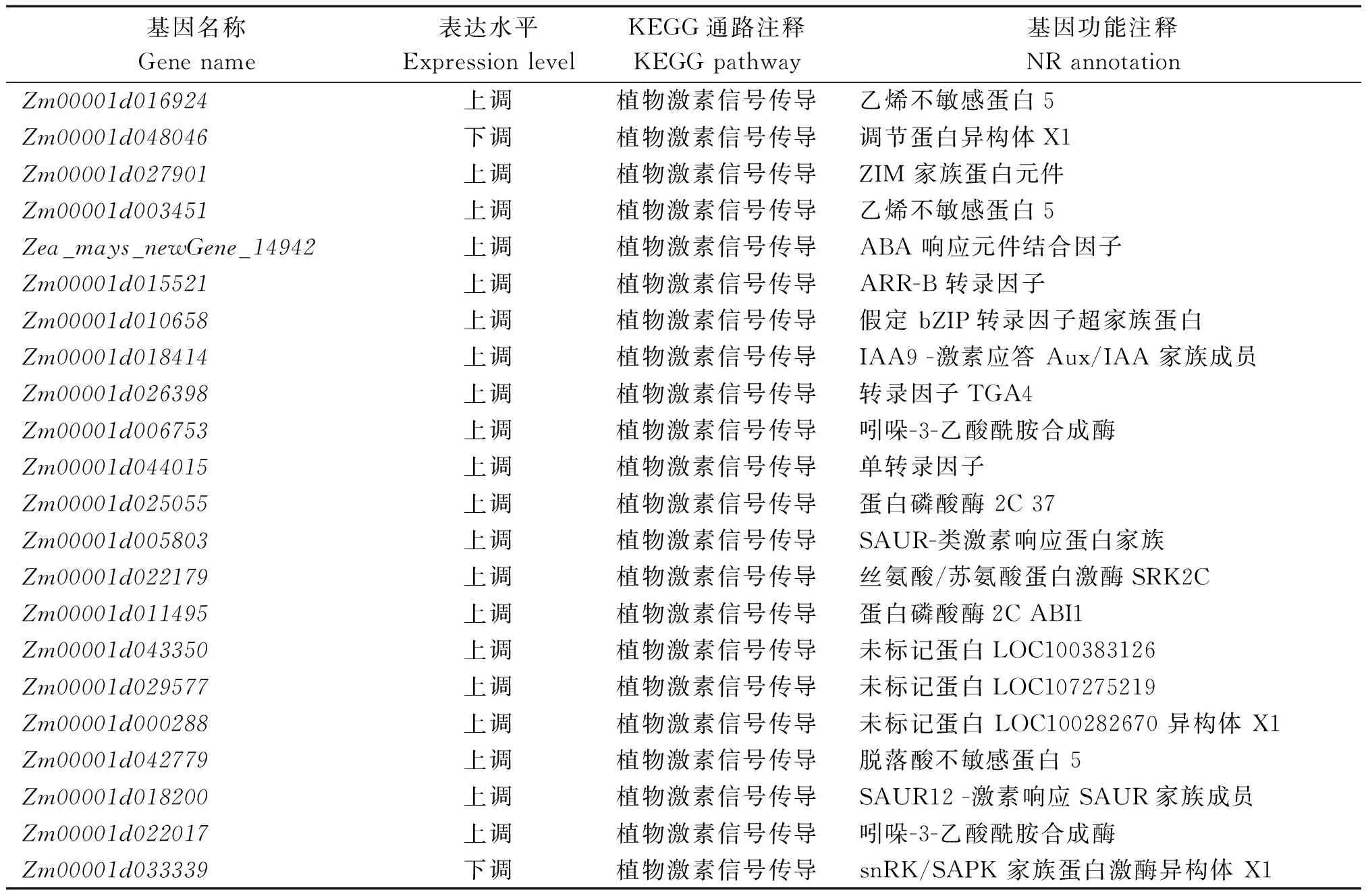
表2 DK517-ABA80 VS DK517-CK植物激素信号传导通路中共同差异表达基因Tab.2 List of DEGs in plant hormone signal transduction pathway in DK517-ABA80 VS DK517-CK group
由图3可以看出,ZD1002-ABA80 VS ZD1002-CK差异表达基因注释到12个不同代谢通路中,每个代谢通路只有1个差异表达基因。12个不同的代谢通路分别为蛋白酶体、RNA降解、核苷酸切除修复、基础转录因子、丙氨酸天冬氨酸及谷氨酸代谢、精氨酸生物合成、氨基酸及核苷酸代谢、果糖及甘露糖代谢、光合器官中碳固定、氨基酸生物合成、碳代谢、2-氧化羧酸代谢通路。

图3 外源ABA处理后ZD1002-ABA80 VS ZD1002-CK对比组差异表达基因KEGG分类图Fig.3 DEGs KEGG taxonomic map of ZD1002-ABA80 VS ZD1002-CK group treated by ABA
由图4可以看出,DK517-ABA80 VS ZD1002-ABA80差异表达基因注释到107个不同代谢通路中,其中富集到的差异表达基因≥10的代谢通路主要有内吞作用、过氧化物酶体、激酶信号系统、植物激素信号传导、内质网中蛋白质加工过程、RNA降解、泛素介导的蛋白质水解、DAN复制、同源重组、不匹配修复、核苷酸修复、接合体、RNA转运蛋白、核糖体、真核细胞中核糖体合成、mRNA监测途径、半胱氨酸及蛋氨酸代谢、苯丙醇生物合成、氨基酸及核苷酸代谢、柠檬酸循环、果糖及甘露糖代谢、糖酵解/糖异生、乙醛酸及二羧酸酯代谢、淀粉及蔗糖代谢、光合器官中碳固定、氨基酸生物合成、碳水化合物代谢、脂肪酸代谢、甘油脂类代谢、谷胱甘肽代谢、嘌呤代谢、嘧啶代谢、植物病原体相互作用通路。植物激素信号传导系统通路中共富集24个差异表达基因(表3)。
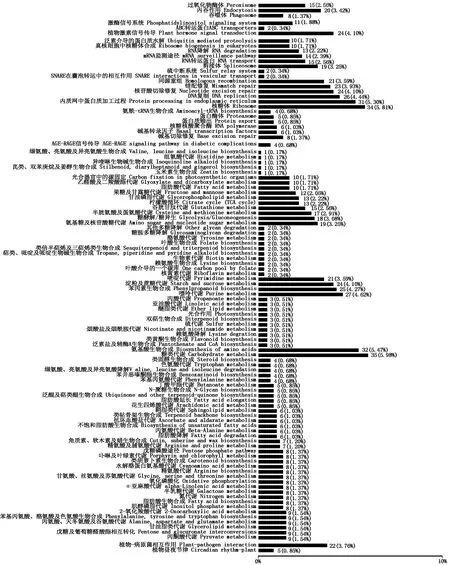
图4 外源ABA处理后DK517-ABA80 VS ZD1002-ABA80对比组差异表达基因KEGG分类图Fig.4 DEGs KEGG taxonomic map of DK517-ABA80 VS ZD1002-ABA80 group treated by ABA

表3 DK517-ABA80 VS ZD1002-ABA80植物激素信号传导通路中共同差异表达基因Tab.3 List of DEGs in plant hormone signal transduction pathway in DK517-ABA80 VS ZD1002-ABA80 group
2.4 外源ABA处理相关转录因子分析
DK517及ZD1002穗位叶喷施外源ABA后,转录组测序后进行转录因子分析,获得多个重要的转录因子家族,结果如图5所示,主要有AGC蛋白激酶家族、AP2/ERF转录因子家族、AUX/IAA转录因子家族、B3转录因子家族、ARID转录因子、Alfin-like转录因子、Aur转录因子家族、BBR-BPC转录因子家族、BES1转录因子、BUB转录因子、C2C2-CO-like转录因子。在本研究中检测到51个差异表达基因含有AGC类转录因子家族。检测到193个差异表达基因含有AP2/ERF-ERF转录因子,24个差异表达基因含有AP2/ERF-AP2转录因子,5个差异表达基因含有AP2/ERF-RAV转录因子。检测到53个差异表达基因含有AUX/IAA类转录因子家族。检测到91个差异表达基因含有B3类转录因子家族。
2.5 外源ABA对不同品玉米种脱水速率相关基因的影响
图6 对比分析了外源ABA对不同玉米品种脱水速率相关基因的影响。DK517-ABA80 VS DK517-CK(G1对比组)、ZD1002-ABA80 VS ZD1002-CK(G3对比组)共检测到了10个共同差异表达基因,分别为:Zm00001d035000,在G1对比组中上调表达而在G3对比组中下调表达,在玉米中表达未标记蛋白LOC100279096;Zm00001d042063,在G1、G3对比组中均呈上调表达,玉米植株中表达假设蛋白 ZEAMMB73;Zm00001d045314,在G1对比组中上调表达而在G3对比组中下调表达,在玉米中表达S-腺苷甲硫氨酸依赖性甲基转移酶;Zm00001d003938,在G1对比组上调表达而在G3对比组下调表达,在玉米中表达甾醇皂苷;Zm00001d028647,在G1对比组中呈上调表达而在G3对比组中呈下调表达,在玉米中表达未标记蛋白LOC100280456;Zm00001d029778,在G1对比组中呈上调表达而在G3对比组中呈下调表达,在玉米植株中表达EC蛋白同系物;Zm00001d032186,在G1、G3对比组中均呈上调表达,在玉米植株中表达未标记蛋白LOC100192110前体;Zm00001d033447,在G1对比组中呈上调表达而在G3对比组中呈下调表达,功能未知;Zm00001d037988,在G1对比组中呈上调表达而在G3对比组中呈下调表达,在玉米植株中表达EMB564蛋白;Zm00001d046065,在G1对比组中呈上调表达而在G3对比组中呈下调表达,在玉米植株中表达组蛋白乙酰转移酶B催化亚体。G1对比组中存在1 722个特异的差异表达基因;G3对比组中存在42个特异的差异表达基因。

图5 外源ABA处理不同玉米材料中转录因子分析结果Fig.5 Transcription factor analysis map of different material groups treated with exogenous ABA

G1.DK517-ABA80 VS DK517-CK;G2.DK517-ABA80 VS ZD1002-ABA80;G3.ZD1002-ABA80 VS ZD1002-CK。
G1对比组与DK517-ABA80 VS ZD1002-ABA80(G2对比组)共检测到了268个共同差异表达基因;G1对比组中存在1 464个特异的差异表达基因,G2对比组中存在3 599个特异的差异表达基因。
G2对比组与G3对比组中共检测到了15个共同差异表达基因,分别为:Zm00001d035000,在G1对比组中上调表达而在G3对比组中下调表达,在玉米中表达未标记蛋白LOC100279096;Zm00001d042063,在G1对比组中下调表达而在G3对比组中上调表达,在玉米中表达假设蛋白 ZEAMMB73;Zm00001d045314,在G1对比组中上调表达而在G3对比组中下调表达,在玉米中表达S-腺苷甲硫氨酸依赖性甲基转移酶;Zea_mays_newGene_10770,在G2对比组中上调表达而在G3对比组中下调表达,在玉米中表达假设蛋白E3泛酸蛋白连接酶SIS3;Zea_mays_newGene_18254、Zea_mays_newGene_20460均在G2对比组中上调表达而在G3对比组中下调表达,功能未知;Zea_mays_newGene_21290,在G2对比组中上调表达而在G3对比组中下调表达,玉米植株中表达未标记蛋白LOC109940605;Zm00001d003872在G2对比组中下调表达而在G3对比组中上调表达,玉米植株中表达未标记蛋白LOC100280055 异构体 X1;Zm00001d009259,在G2对比组中上调表达而在G3对比组中下调表达,在玉米植株中表达丝氨酸/苏氨酸蛋白激酶RUNKEL;Zm00001d011419,在G2对比组中下调表达而在G3对比组中上调表达,在玉米植株中表达细胞色素P450 72A15;Zm00001d017019,在G2对比组中下调表达而在G3对比组中上调表达,在玉米植株中表达ATFP4蛋白;Zm00001d024522,在G2对比组中下调表达而在G3对比组中上调表达,在玉米植株表达bHLH转录因子;Zm00001d028374,在G2对比组中下调表达而在G3对比组中上调表达,在玉米植株中表达早期真菌诱导蛋白CMPG1;Zm00001d032658,在G2对比组中下调表达而在G3对比组中上调表达,在玉米植株中表达假设蛋白ZEAMMB73_Zm00001d032658;Zm00001d034413,在G2对比组中上调表达而在G3对比组中下调表达,在玉米植株中表达球蛋白2前体。G2对比组中存在3 852个特异的差异表达基因;G3对比组中存在37个特异的差异表达基因。
ABA影响玉米籽粒灌浆速率及脱水速率。对检测到的差异表达基因GO分类分析后获得73个DEG与水分代谢相关,推测这些基因正是作为外源ABA调控的下游基因起作用,具体见表4。
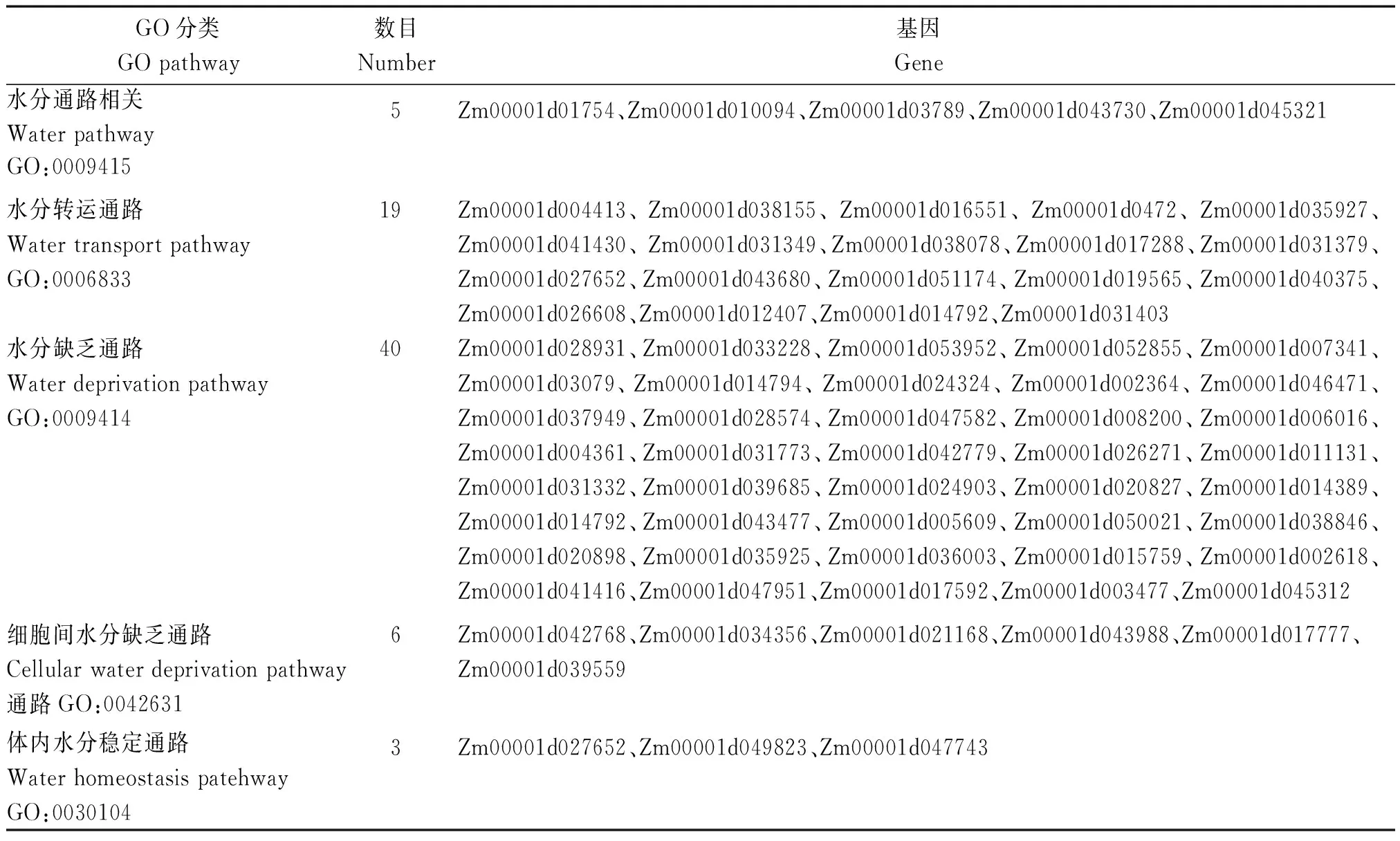
表4 DK517和ZD1002叶片中与水分代谢通路相关的差异表达基因筛选Tab.4 List of DEGs which is related with water metabolism pathway in DK517 and ZD1002 leaf samples
3 结论与讨论
在花粒期对籽粒不同脱水速率玉米品种DK517、ZD1002喷施外源ABA,对12个穗位叶样本进行转录组测序分析,发现脱水速率较快的品种DK517在喷施外源ABA后有1 732个基因的表达水平达到显著差异水平,其中1 251个上调表达,481个下调表达。脱水速率较慢的品种ZD1002在喷施外源ABA后只有52个基因的表达水平达到显著差异水平,其中24个上调表达,28个下调表达。DK517中有更多参与ABA调控籽粒脱水过程的基因在表达水平上发生了显著变化,推测正是这些基因共同作用加快了籽粒脱水速率,使得DK517适宜机收。DK517中上调表达基因个数占总差异表达基因个数的72.23%,ZD1002中上调表达基因占总差异表达基因的46.15%,远低于DK517中上调表达基因所占比例。
DK517差异表达基因GO分析结果主要富集于19个不同生物学过程分类、12个不同细胞组分分类、12个不同分子功能分类;主要分布于染色体结构及动态变化过程等23个COG 数据库分支;注释到100个不同代谢通路中。ZD1002差异表达基因GO分析结果主要富集于13个不同生物学过程分类、10个不同细胞组分分类、4个不同分子功能分类;主要分布于细胞循环控制细胞分裂及染色体分区等8个COG 数据库分支;注释到12个不同代谢通路中。由此推测,不同玉米籽粒脱水速率品种中外源ABA调控的内在分子机制不同。
转录因子是真核生物中广泛存在的能与DNA特异性结合的蛋白质因子,其通过与顺式作用元件特异性结合调控下游基因表达,参与植物生长发育过程中众多的生物学功能。AGC蛋白激酶家族由一系列在结构和功能上相关的激酶组成,包含蛋白激酶A、蛋白激酶G、蛋白激酶C,共有63个家族成员,与细胞内许多信号通路相关[27]。本研究中,共检测到51个差异表达基因含有AGC类转录因子家族。AP2/ERF转录因子家族广泛存在于植物中,含有保守的AP2/ERF结构域。根据该结构域内序列的相似性和AP2/ERF结构域的数量,可以将植物AP2/ERF类转录因子分成DREB (Dehydration-responsive element binding protein)、ERF (Ethylene-responsive element binding factors)、 AP2 (APETALA 2)、RAV (Related to ABI3/VP1)和其他类别等5个亚家族[28]。AP2/ERF转录因子参与多种生物学过程,包括干旱、高盐、低温等逆境胁迫响应等。在植物受到逆境刺激后,脱落酸、乙烯等信号相应被激活,并激活AP2/ERF转录因子的表达,从而调控功能基因的表达。本研究中,共检测到193个差异表达基因含有AP2/ERF-ERF转录因子,24个差异表达基因含有AP2/ERF-AP2转录因子,5个差异表达基因含有AP2/ERF-RAV转录因子。AUX/IAA为生长素调节类转录因子。生长素水平和生长素信号通路调控系统对植物防御网络有重要作用,生长素也可以直接作为抗菌活性的防御分子参与植物免疫反应过程的调节。生长素信号传导主要受到AUX/IAA和生长素响应因子(ARFs)2类转录因子的调控。ARFs具有能够与DNA结合的区域,可以与生长素响应基因启动子中的生长素响应元件结合,激活生长素响应基因的表达。在不含有生长素或生长素浓度很低的环境下,AUX/IAA蛋白能与ARFs结合,使其不能激活下游生长素响应基因的表达;当生长素浓度升高时,生长素能够促进AUX/IAA蛋白的泛素化降解,解除其对ARFs的抑制作用,从而使下游生长素响应基因的表达被成功激活[29]。本研究中,共检测到53个差异表达基因含有AUX/IAA类转录因子家族。B3转录因子家族在被子植物中广泛存在,数目众多,具有高度保守的B3-DNA结合结构域,分为ARF (Auxin responsefactor)、ABI3/VP1、HSI (High-level expression of sugar-inducible gene)、RAV (Related to ABI3/VP1)、REM(Reproductive meristem) 5个家族。其中,B3-ARF转录因子亚家族属于生长素响应因子,主要参与生长素响应有关的植物生理功能。B3转录因子家族在植物激素的合成以及信号转导、种子发育、开花调控、生物及非生物胁迫应答中发挥着重要作用[30]。本研究共检测到91个差异表达基因含有B3类转录因子家族。
猜你喜欢
中国人兽共患病学报(2022年9期)2022-10-19
舰船科学技术(2022年11期)2022-07-15
中国农学通报(2022年13期)2022-05-31
现代畜牧科技(2021年4期)2021-12-05
中国土壤与肥料(2021年5期)2021-12-02
科学导报(2021年29期)2021-06-03
中国生殖健康(2020年4期)2021-01-18
山东建筑大学学报(2020年5期)2020-09-16
科海故事博览·下旬刊(2019年6期)2019-04-16
科学种养(2017年6期)2017-06-13
