
不同温度处理对疏序黄荆种子萌发的影响
2020-10-22 13:22李昭华郭超凡龙左卿周绍齐苏文华
种子 2020年9期
李昭华,郭超凡,刘 婕,殷 丽,龙左卿,沙 月,周绍齐,苏文华,周 睿
(云南大学生态学与地植物学研究所,昆明 650500)
干热河谷是对又干又热的河谷地区的统称[1],是一类主要分布于中国西南地区(云南、四川)的特殊气候地貌类型。在云南境内,干热河谷主要分布于怒江、金沙江、澜沧江、红河等流域[1,2]。其中比较典型的如元谋干热河谷、元江干热河谷等[3]。干热河谷地区典型的气候特征是全年干湿季分明,热量高、降水少,水热严重失衡,蒸发作用强烈[2,4-7],全年蒸发量是降水量的3~6倍,年均温大于20 ℃,而年积温大于7 000 ℃[8]。这种特殊的气候条件使得干热河谷地区呈现出以稀树灌草丛占优势的植被类型,由于与非洲稀树草原的植被类型相似,因此被称为“河谷型”萨瓦那植被类型(Savanna of valley type)[9]。
由于气候干热,导致干热河谷地区成为全球最容易发生火灾的区域之一[10,11]。火灾是几乎跟陆地植物同时出现在地质记录中的一种世界性自然现象,影响着全球生态系统的分布和生态过程,如植物的分布、结构,物质循环如碳循环、氮循环等[12]。不同强度和频率的火灾,往往会造成不同程度的生态影响[12]。Alves等[13]通过遥感监测数据发现,一定强度的周期性燃烧对物质循环、植物更新有着重要的意义。而过高的燃烧强度或频率则不利于维持生态系统的平衡[14,15]。
在火的长期干扰下,植物也由此进化出了一系列适应机制。与易火生境有关的植物特性称为火相关性状(fire-related traits)[16,17]。此外,Lamont等[17],Zuloaga-Aguilar等[18],He等[19]发现,促使火相关性状产生的因素不仅是火,还有燃烧过程中产生的烟雾、热量、灰分等都能导致植物产生一系列相关性状。这些适应性状包括厚树皮、萌生、枝下自疏、高温或烟雾刺激种子萌发、硬质种皮、火后开花等[16,17,20-23]。
在易火生境中,种子的萌发常常与火密切相关[21]。火可以减少土壤和凋落物中的抑制物质、木炭中所含物质的化学刺激、破坏硬质种皮等[24]。通常将经过40 ℃以上温度处理后种子萌发率或萌发速度提高的现象称为热冲击效应[25-27]。研究表明,火燃烧时产生的高温对土壤种子库等具有热冲击效应[28,29]。热冲击可以打破种子休眠,改变种皮结构,从而促进种子吸水与萌发[20,30,31]。
结合干热河谷地区易火的特性及植物对火干扰的适应,对干热河谷地区的主要植物群落物种进行热冲击等火适应相关性实验,对当地的植被恢复、生态建设具有重要意义。金振洲等早期对干热河谷地区植物区系的调查研究表明,疏序黄荆(VitexnegundoL.f.laxipaniculataPei)、坡柳(DodonaeaviscosaL.Jacq)、黄茅(Heteropogoncontortus)、余甘子(PhyllanthusemblicaLinn.)等均为当地较为常见的植物群落成分[3]。
目前,对坡柳、黄茅的热冲击研究已有报道,霍冬芳等[32]、余娇娥等[33]先后发现坡柳、黄茅种子经过高温处理后萌发率较常温对照组有明显提高,表现出较强的热冲击效应。而对疏序黄荆的研究目前主要集中在药用价值方面。因此,探究疏序黄荆这一干热河谷地区植物群落的常见物种是否具有热冲击效应很有必要,这对“河谷型”萨瓦那植被的保护与恢复具有重要意义。
1 材料与方法
1.1 研究地点概况
元谋干热河谷位于云南省楚雄彝族自治州北部(25°23′~26°06′N,101°35′~102°06′E)。区域内热量、光照充足,蒸发量大,降水少[6],年均降雨量约613.7 mm,年均蒸发量约3 426.3 mm,年均相对湿度54%,降水主要集中于5—10月[34],属于亚热带干热季风气候。
1.2 实验材料
疏序黄荆(V.negundoL.f.laxipaniculataPei)是马鞭草科(Verbenaceae)牡荆属(Vitex)黄荆的一个变型,小乔木;小枝被灰褐色绒毛,小叶片椭圆形,全缘,两面有绒毛;圆锥花序大而开展,侧枝明显地分出小枝,第一回侧枝长可达21 cm;果实顶端有毛,花果期5—8月。产四川、云南,生于海拔450~1 400 m的河边密林或山坡灌丛中[35]。
于2019年1月在元谋县城西南方向法那禾村附近的阳坡面(25°42′N,101°51′E)随机选取生长良好的植株,采集植株≥30株,每株≥30粒种子。所有种子采集后进行混合并风干保存,筛选大小均匀、颗粒饱满、色泽一致的种子进行实验。
1.3 实验方法
1.3.1材料处理
为了让实验效果具有可比较性,对种子进行萌发实验前的热冲击处理,主要参照Ribeiro等[27]的方法进行。从筛选后的疏序黄荆种子中随机选择,分别采用60 ℃(10,20,40 min)、80 ℃(5,10,20 min)、100 ℃(2.5,5,10 min)共9种方式在烘箱中进行干热处理。每个处理10个重复,每个重复包含随机选出的30粒种子。另设置10个重复进行常温(25 ℃)处理作为对照组,每个重复包含随机选择的30粒种子;整个热冲击实验共使用随机选出的种子3 000粒。
在烘箱中先放入10个平面玻璃培养皿,按60 ℃(10,20,40 min)、80 ℃(5,10,20 min)、100 ℃(2.5,5,10 min)共9种处理方式分别设定好处理温度,待达到设定温度后,迅速将种子放入培养皿中,每个培养皿放30粒种子,避免种子叠放;达到处理时间后,迅速将种子取出,放入预备好的其他培养皿中,9种处理方式依次进行。待所有处理的种子自然冷却至室温后,进行萌发实验。
此外,为检验各处理对种子活性的影响,再另外随机挑选1 000粒大小均匀、色泽一致的饱满种子,分别采用60 ℃(10,20,40 min)、80 ℃(5,10,20 min)、100 ℃(2.5,5,10 min)共9种方式在烘箱中进行干热处理。每个处理5个重复,每个重复包含随机选择的20粒种子。冷却至室温后,连同常温对照组将种子在蒸馏水中浸泡48 h。然后用0.3%的TTC溶液对种子活力进行检测。
1.3.2萌发实验
在直径7 cm的培养瓶底部铺上双层滤纸,用蒸馏水润湿,而后将不同温度和时间处理的种子均匀放入培养瓶中进行萌发实验。
培养条件:温度25 ℃恒温,光照强度1 250 lx,光照时间每天14 h。每天对种子的萌发数量进行统计并记录(种子萌发以胚根突破种皮2 mm为准)。同时适当补充水分,保持培养瓶底部湿润,以种子周围不出现水膜为宜,连续观察3周(21 d)。
1.3.3种子活力测定
用解剖刀将浸泡好的种子沿胚的中线切成两半,将其中一半置于准备好的培养皿中并加入0.3%的TTC溶液浸没种子,而后在40 ℃恒温条件下染色40 min,最后用蒸馏水冲洗2~3次,并将种子置于解剖镜下观察染色情况,同时记录种子空壳、坏死、着色情况,以探究不同温度和时间处理对种子活力的影响。种子着色越深,则活力越强。
1.4 数据处理
参照余娇娥等[33]、董沁方等[36]、常云霞等[37]的方法,计算种子萌发率、萌发势以及热冲击效应指数(RI)公式如下:
萌发率(%)=(n/N)× 100%;
萌发势(%)=(m/N)× 100%;
其中,n为萌发种子数;m为萌发实验开始前7 d内的萌发种子数;N为供试种子数。
当T≥C时,RI=1-C/T;
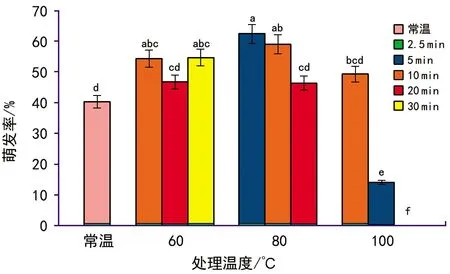
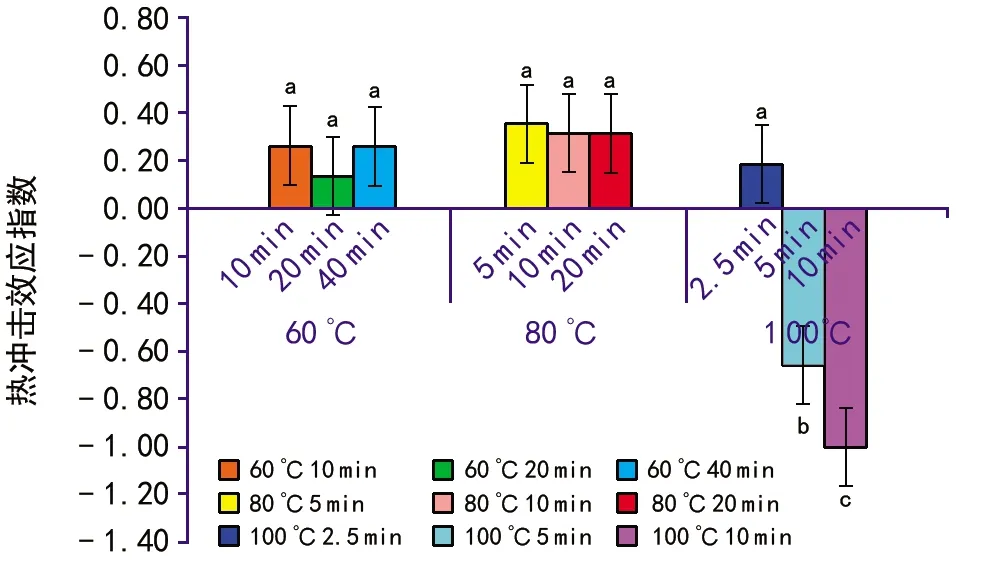
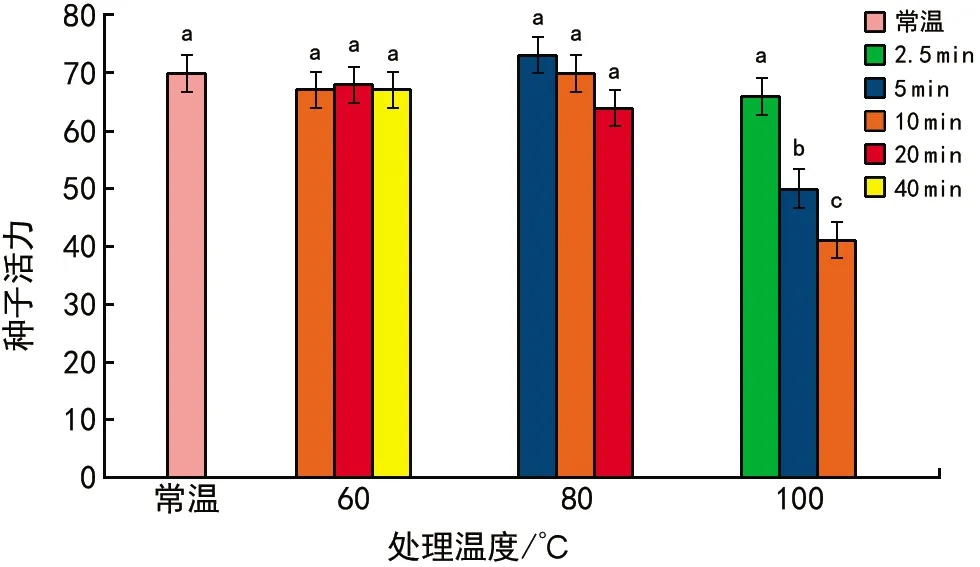
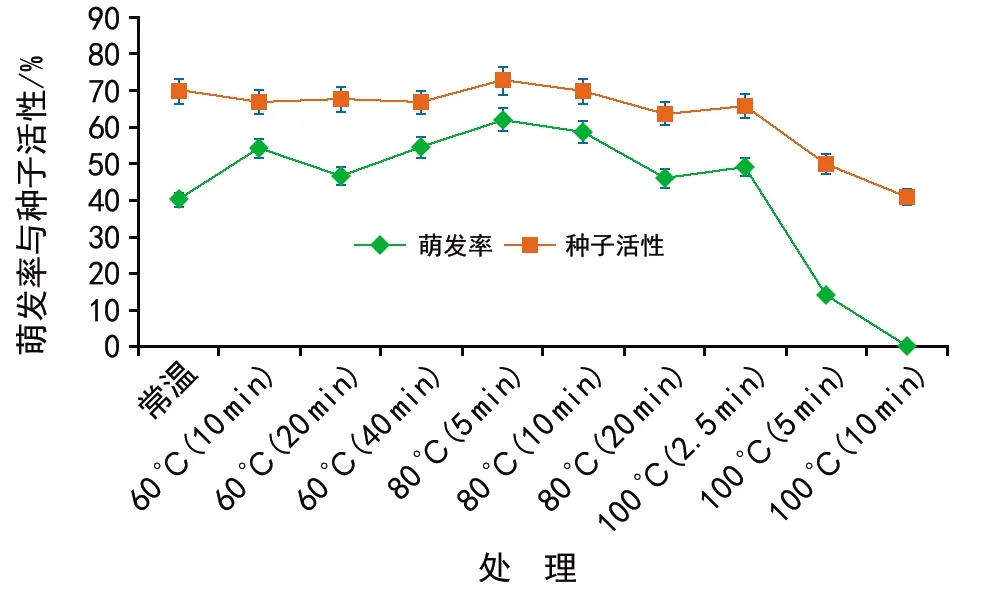
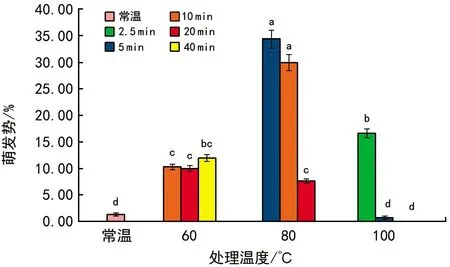
当T 其中,C为对照值,T为处理值。当RI>0时,为促进作用,当RI<0时为抑制作用。RI的绝对值代表作用强度的大小。 种子活力(%)=(染成红色的种子数/供试种子数)×100%。 用Excel 2010软件统计和作图,用SPSS 22.0软件通过95%水平上进行单因素方差(ANOVA)分析,采用Duncan检验法进行多重比较及差异显著性检验(α=0.05),并进行主效应方差分析,图表数据为平均值±标准差。 2.1.1萌发率的变化 经过60 ℃(10,20,40 min)、80 ℃(5,10,20 min)、100 ℃(2.5,5,10 min)以及常温对照10种方式处理后,疏序黄荆种子萌发率在80 ℃(5 min)处理时达到最大,为(62.33±3.13)%;100 ℃(10 min)处理时萌发率最低,为0(图1)。从整体上来看,随着处理温度和时间的变化,种子萌发率呈先上升后下降的趋势。除100 ℃(5 min)和100 ℃(10 min)2种处理方式萌发率低于常温对照外,其余各组萌发率均高于常温对照组,萌发率分别提高了14.00%,3.97%,14.33%,19.63%,18.67%,3.63%,9.00%。其中,60 ℃(10 min)、60 ℃(40 min)、80 ℃(5 min)、80 ℃(10 min)4组的萌发率显著高于常温对照组;而100 ℃(5 min)和100 ℃(10 min)2组的萌发率显著低于包括常温对照在内的其余各组。 图1 高温处理对疏序黄荆种子萌发率的影响 2.1.2热冲击效应指数 可以用热冲击效应指数(RI)来表示热冲击对疏序黄荆种子萌发率的影响,热冲击效应指数大于0,表明热冲击对种子萌发有促进作用;小于0,则表明热冲击对种子萌发有抑制作用。 实验结果(图2)表明,60 ℃(10,20,40 min)、80 ℃(5,10,20 min)、100 ℃(2.5,5,10 min)9种处理方式的热冲击效应指数分别为0.257 7,0.135 7,0.262 2,0.352 9,0.316 4,0.316 4,0.182 4,-0.652 9,-1.000 0。除100 ℃(5 min)和100 ℃(10 min)2组热冲击效应指数RI小于0,抑制了种子萌发外,其余各组均大于0,在不同程度上促进了种子萌发。80 ℃(5 min)时热冲击效应指数达到最大。热冲击效应指数在整体上呈现出先上升后下降的趋势。 图2 不同温度和时间处理疏序黄荆种子的热冲击效应指数 2.1.3主效应方差分析 为了研究不同温度和时间处理对疏序黄荆种子萌发率的影响,采用SPSS软件中的一般线性模型,对萌发前不同温度和时间处理的疏序黄荆种子萌发率进行单变量双因素方差分析。 数据分析结果(表1)表明,温度和时间对疏序黄荆种子萌发率的主效应均达到极显著水平。根据各因素F值的大小,萌发前的处理温度影响最大(F=148.092,p<0.001),其次是处理时间的影响(F=32.553,p<0.001),最后是温度和时间互作的影响(F=1.346,p=0.226)。 表1 萌发率主效应方差分析结果 2.1.4萌发前高温处理对疏序黄荆种子活力的影响 从种子的解剖和染色情况来看,除100 ℃(5 min)和100 ℃(10 min)2种处理方式外,其余处理组之间以及他们与常温对照之间活性种子数无显著性差异(图3)。经100 ℃(5 min)和100 ℃(10 min)处理后,活性种子数显著低于常温对照组(图3)。在100 ℃(10 min)处理时活性种子数最低,为(41±0.84)%。表明种子对高温具有一定的耐受性限度,过高的温度会导致种子活力下降,从而影响种子萌发率。在实验中,种子萌发率与种子活力随不同高温处理表现出相似的趋势(图4)。 图3 高温处理对疏序黄荆种子活力的影响 图4 不同高温处理下疏序黄荆种子萌发率和种子活力的变化趋势 实验采取前1/3期间(7 d)内的萌发数占供试种子数的百分比计算萌发势。经过60 ℃(10,20,40 min)、80 ℃(5,10,20 min)、100 ℃(2.5,5,10 min)以及常温对照10种方式处理后,除100 ℃(5 min)和100 ℃(10 min)外,其余各组萌发势均高于常温对照组(图5)。萌发势在80 ℃(5 min)处理时达到最高;100 ℃(10 min)处理时萌发势达到最低。 图5 高温处理对疏序黄荆种子萌发势的影响 在一定的温度范围内,短时高温处理可以明显提高疏序黄荆种子的萌发速度。萌发势整体呈先上升后下降的趋势,在80 ℃(5 min)处理时达到最大,而温度过高则不利于种子的萌发,反而出现抑制效应。 余娇娥等研究发现,经60 ℃和80 ℃处理后,黄茅种子萌发率显著提高[33];霍冬芳等发现,坡柳种子经过40 ℃以上的高温处理后,具有明显的热冲击效应,在80 ℃(10 min)处理时萌发率达到最大[32];Williams等研究发现,80~100 ℃的短时高温处理可以打破Chamaecristamimosoides,Crotalariacalycina,Crotalariamontana,Indigoferahirsuta,Tephrosiajuncea等植物的种子休眠,从而促进种子萌发[20];Gashaw等对埃塞俄比亚西部的稀树草原进行研究时发现,一些植物种子在短时高温处理后表现出萌发率提高的现象[26]。 在本实验中,除100 ℃(5 min)和100 ℃(10 min)2种处理方式萌发率低于常温对照外,其余各组萌发率均显著提高,在80 ℃(5 min)处理时达到最大,为(62.33±3.13)%,表现出一定的热冲击效应。说明疏序黄荆种子也有对高温的适应对策。热冲击不仅提高了疏序黄荆的萌发率,还在一定程度上提高了其萌发势。除100 ℃(5 min)和100 ℃(10 min)2种处理方式萌发势低于常温对照组外,其余各组萌发势均高于对照,说明热冲击处理可以缩短种子萌发所需时间。同时,实验结果也表明,疏序黄荆种子对高温也有一定的耐受性限度,当超过一定的温度及处理时间后,就会对种子产生损害,从而影响种子萌发。 就热冲击效应产生的的机制而言,Zuloaga-Aguilar等[18],Jeffery等[30]的研究表明,热冲击可以改变种皮结构,使种皮的透水性增强,增加气体交换,从而促进种子吸水萌发;常云霞等在云南松的热冲击实验中发现,经过短时高温处理后,种子中的超氧化物歧化酶(SOD)、过氧化酶(POD)和过氧化氢酶(CAT)活性均显著提高[37]。说明热冲击可以激活与种子萌发相关的酶系统,从而促进种子萌发。Zuloaga-Aguilar[18],Nelson[38]等在研究烟雾等化学信号对种子萌发的影响时发现,外界的刺激与种子的休眠状态密切相关。也就是说,处于深度休眠状态的种子对外界刺激不敏感。而霍冬芳等对经过不同存储时间的坡柳进行热冲击实验,发现萌发率并没有显著变化[32]。此外,火对植物种子的影响还与种子的形状、大小、质量等密切相关[16,20,39,40]。因此,关于疏序黄荆种子的热冲击效应是否与内部酶系统活性的变化、休眠状态、种子质量、大小、形状等因素有关,还有待进一步研究。 He等通过对木炭等的研究分析表明,火对植物的干扰历史在3.5亿年以上[19]。所以,在如此长的时期内,植物必然进化出了一系列对火的适应机制。虽然Lamont等[17]也指出,一些植物的火适应性状可能并非是火诱导的,也可能是由干旱诱导的,干旱对植物的影响可以看成是火的一种温和形式。但就强度与时间而言,干旱造成的高温持续时间长,可是强度不大;而火造成的高温持续时间虽然没有干旱的持续时间长,但其温度远高于干旱产生的高温。因此,干旱与火所造成的影响是有区别的,是否能将易火生境中的植物火适应性状用干旱来进行解释,还有待进一步验证。 干热河谷作为全球最容易发生火灾的地区之一,对当地优势植物进行火相关研究十分必要,可以为干热河谷地区的植被恢复、人工播种等提供必要的参考。在疏序黄荆的播种过程中,可以采取80 ℃(5 min)的热冲击处理,从而提高种子的萌发率,减少恢复成本。2 结果与分析
2.1 萌发前高温处理对疏序黄荆种子萌发率的影响

2.2 萌发前高温处理对疏序黄荆种子萌发势的影响
3 结论与讨论
猜你喜欢
小哥白尼(神奇星球)(2021年9期)2021-11-19
小哥白尼(神奇星球)(2021年5期)2021-07-28
小哥白尼(神奇星球)(2021年3期)2021-07-22
小哥白尼(神奇星球)(2021年4期)2021-07-22
保鲜与加工(2021年1期)2021-02-06
疯狂英语·新悦读(2020年10期)2020-11-26
爆笑show(2015年3期)2015-05-08
汽车维修与保养(2015年12期)2015-04-18
汽车维修与保养(2015年6期)2015-04-17
汽车维护与修理(2015年6期)2015-02-28
