
高原鼠兔和高原鼢鼠肠道微生物组及常见抗生素抗性基因的比较
2020-10-27 02:42冯天舒俞巧玲王逸洁苏军虎
野生动物学报 2020年4期
冯天舒 俞巧玲 周 蕊 王逸洁 苏军虎 李 欢,3*
(1.兰州大学公共卫生学院,兰州,730000;2.甘肃农业大学草业学院,兰州,730070;3.兰州大学草地微生物研究中心,兰州,730000)
哺乳动物的胃肠道定植有数百种有益于健康的微生物[1]。通常这些动物的肠道在出生时是无菌的,但很快就会从母体、饮食、周围环境和其他个体中获得微生物[2]。肠道微生物群大约由数百到数千种微生物组成[3],并具有多种功能,包括多糖的消化,纤维素的分解,能量的平衡以及免疫调节等功能[1,4-5]。鉴于肠道菌群对动物健康的重要作用,它被认为是哺乳动物的一个新“器官”[6-7]。除了以上功能,肠道菌群在动物的适应性进化过程中也发挥了重要作用。比如,青藏高原哺乳动物肠道微生物组进化出了很强的食物利用能力和纤维素降解活性,并且能够产生更多的短链脂肪酸[8],而短链脂肪酸是纤维素等多糖类物质经哺乳动物肠道微生物发酵产生的主要代谢产物,在宿主对营养物质的利用、免疫调节、脂质代谢和糖异生等生理功能中起着关键作用[9]。肠道微生物在具备众多功能的同时,也受到诸多因素的影响,比如膳食、环境以及宿主的种类均能影响哺乳动物肠道菌群的组成[10-11],而肠道菌群组成的改变可以导致其功能发生变化[12],继而可能影响动物的生理和适应性。生活在青藏高原的野生动物的肠道微生物一样会受到这些因素的影响。
高原鼠兔(Ochotonacurzoniae)和高原鼢鼠(Eospalaxfontanieriibaileyi)均是青藏高原(Qinghai-Tibet Plateau)高寒草甸(alpine meadow)生态系统中重要的物种,在高寒草甸物质流动和能量循环中发挥着重要作用[13-14],但它们的食性和栖息环境存在差异。高原鼠兔属于哺乳纲(Mammalia),兔形目(Lagomorpha),鼠兔科(Ochotonidae),鼠兔属;它们在洞穴中生活;是植食性动物,喜食地面上的植物,以植物叶、茎、花、根、芽为食,较少摄入种子和粗老纤维化或含水量少的器官或组织,主要以禾本科(Gramineae)和豆科(Leguminosae)植物为食[15]。由于它与牲畜争夺青草,进而导致牧场退化,因而一直被认为是有害的物种[16]。然而,一些生态学家指出,高原鼠兔有助于加速土壤氮循环,对生态系统作出了积极贡献;因此,高原鼠兔被列为青藏高原重点物种并被誉为生态系统工程师[17]。高原鼢鼠属于啮齿目(Rodentia),仓鼠科(Cricetidae),鼢鼠亚科(Myospalacinae),鼢鼠属;它们通常在地下生活;也属于植食性动物,但通常以植物根、皮、茎、叶、果实和种子为食,喜食伴生杂草,对禾本科和莎草科(Cyperaceae)摄入较少。高原鼢鼠和高原鼠兔生态功能相似,并被认为是高寒草甸土壤运动的重要媒介[18-19]。由于青藏高原平均海拔在4 000 m以上,是地球上最高的高原,食物匮乏,具有独特的缺氧、寒冷、强紫外线等恶劣气候和地理环境[20-21]。这一系列的环境压力会导致动物生理活动的改变,进而可能会影响高原鼠兔和高原鼢鼠肠道微生物的组成和功能。
作为20世纪最重要的医学发现之一,抗生素(antibiotic)在控制感染性方面发挥了重要作用,然而,由于近年来抗生素的滥用,导致了一大批耐药性致病菌的出现。有研究表明仅有少量的抗生素被有效利用,大部分抗生素及其在动物体内诱导出的抗生素抗性基因随着动物的尿液和粪便直接排出体外。抗生素抗性基因在环境中持续存在以及在不同介质中传播往往引起比抗生素本身更大的危害[22-23],抗生素的滥用增加了人和动物微生物群中耐药基因的流行率[24]。近年来,抗生素耐药已成为全球主要的公共卫生问题[25]。抗生素耐药基因的历史可分为“抗生素前”时期和“抗生素”时期,前者抗生素耐药性的获得主要通过复制和突变,是一个长期的过程;而后者则是一个短暂的过程,大规模生产并使用抗生素在各生态系统中对细菌施加了强大的选择性压力[26]。宿主可以通过水平基因转移从其他细菌获得抗生素抗性基因(antibiotic resistance genes,ARGs),也可由自身产生[26]。有证据说明肠道细菌可以相互交换耐药基因,还可以通过与结肠的细菌相互作用获得并传播抗生素耐药基因[27]。因而,在哺乳动物生命活动中发挥着重要作用的肠道微生物也有可能通过贮存和传播抗生素耐药基因对健康产生危害。例如,土霉素(oxytetracycline)属于四环素(tetracycline)类,有研究发现,蜜蜂(Maryland bees)长期接触这种抗生素后,在其肠道微生物群中发现了四环素抗性基因[28]。目前,有基于培养和亚基因组的研究证实了在人类微生物中存在一个抗生素耐药性的基因库,在对人类微生物群的研究中发现在肠道微生物中携带一些抗药性基因,如ermG、ermB、ermF、tetM、tetQ和vanB[27,29]。尽管高原鼢鼠和高原鼠兔是野生动物,它们的肠道微生物也有可能携带大量的抗生素抗性基因,并可以通过粪便排入水体或土壤环境中,进入人类环境并危害人类健康。另外,由于青藏高原地区存在牦牛(Bosgrunniens)与高原鼠兔和高原鼢鼠互食粪便的现象,因此,其携带的抗生素抗性基因也可以通过牦牛肉或奶制品传递给人类。尽管检测野生动物肠道抗生素基因十分重要,但是还鲜有关于野生动物携带哪些抗生素耐药基因的报道。
本研究中,首先通过高通量测序分析各分类水平下高原鼠兔和高原鼢鼠肠道菌群的组成和多样性,分析其优势菌群,并比较两组间菌群结构和多样性的差异。接着通过定量PCR(qPCR)量化几种抗生素抗性基因在野生动物肠道内的浓度,最后通过冗余分析和Spearman秩相关系数判断抗生素抗性基因和肠道菌群的相互作用关系。拟解决以下科学问题:(1)高原鼢鼠和高原鼠兔肠道微生物的组成和多样性是否有差异;(2)两物种中有哪些抗生素基因得以表达以及表达量的差异;(3)抗生素抗性基因与肠道微生物的关系。本论文旨在为降低和阻止野生动物肠道抗生素抗性基因的传播和蓄积提供科学理论参考。
1 材料与方法
1.1 样品收集
在青海泽库(海拔4 331 m,n=16)和果洛(3 856 m,n=8;3 694 m,n=16)用绳套法捕捉高原鼠兔,它们的平均体重为(139.05±41.10)g,雄性∶雌性为1.22∶1;在甘肃临潭(3 120 m,n=20)和天祝(2 980 m,n=18),用弓形夹捕捉高原鼢鼠,其平均体重为(145.42±25.47)g,雄性∶雌性为1∶1.24。经兰州大学伦理委员会和动物保护协会批准后,对所有动物实施安乐死,即在野外用乙醚将动物麻醉致死并解剖动物,用无菌剪刀剪下盲肠,并立即用无菌镊子将肠道的内容物挤到50 mL无菌管中,所有样品均暂时冻存在-20 ℃便携式冰箱中,并于24 h内运回实验室,在-40 ℃冰箱保存。本次研究共收集高原鼠兔肠道内容物样本40份和高原鼢鼠肠道内容物样本38份。根据之前我们的研究,高原鼠兔的食物主要有棘豆属(Oxytropis)、毛茛属(Ranunculus)、瘤果芹属(Trachydium)、马先蒿属(Pedicularis)和矮嵩草属(Kobresia)植物[30];而高原鼢鼠的食物主要有蔷薇科(Rosaceae)(蕨麻Argentinaanserina,雪白委陵菜Potentillanivea)和禾本科(披碱草Elymus)植物[31]。
1.2 肠道微生物和抗生素基因测定
1.2.1 提取DNA
采用Ezup土壤基因组DNA提取试剂盒(Sangon Biotech,China)提取盲肠内容物总DNA。
1.2.2 PCR扩增
使用通用引物515F(5′-GTGYCAGCMGCCGCGGTA-3′)和909R(5′-CCCCGYCAATTCMTTTRAGT-3′)扩增微生物的16S rRNA基因[30],再使用含独特12 bp条形码的正向引物对每个PCR产物进行标记。用10 ng DNA模板、1×PCR缓冲液、1.5 mM氯化镁、0.4 μM的脱氧核苷酸三磷酸、1.0 μM的引物,0.25 U的Ex-Taq(大连TaKaRa)和无菌水进行25 μL PCR反应,每个样品做2个重复。热循环程序包括在94 ℃下初始变性3 min;然后在94 ℃下变性40 s,56 ℃下退火60 s,72 ℃下延长60 s,共循环30次;最后在72 ℃下延长10 min。
1.2.3 16S rRNA基因测序和生物信息学分析
PCR扩增后,将2个PCR产物混合在一起,使用1.2%琼脂糖凝胶进行电泳。用SanPrep-DNA凝胶提取试剂盒(Sangon Biotech,China)分离纯化正确的条带(—400 bp),并用Nanodrop 2000分光光度计定量,之后将所有样品等量混合。按照制造商的说明,使用TruSeq DNA试剂盒制备测序文库。纯化后的样品文库按照Illumina文库制备方案进行稀释、变性、再稀释、与PhiX(等于最终DNA量的30%)混合,然后应用Illumina Miseq测序仪进行高通量测序。
在QIIME平台上处理原始数据(http://qiime.org/tutorials/tutorial.html)。基于FLASH软件对其进行双末端拼接(paired-end序列的拼接)。根据barcode条形码进行样品拆分。将fastq序列转换为fasta格式的序列,进一步用FASTS-Toolkit软件剔除低质量的序列,和参考序列比对后去除嵌合体。使用Uclust对所有序列进行聚类,将相似度大于97%的序列聚为一种OTU。筛选出OTU代表性序列并对其进行物种注释,用RDP数据库对每一个OTU代表序列进行分类鉴定。剔除只含有1条序列的OTU或者不属于细菌的OTUs。为了标准化样本,使用daisychopper脚本将每个样本抽取相同的序列数(5 092个序列)。使用Origin 2017,绘制门水平和属水平上的堆积图表示群落组成。计算群落的Chao指数(Chao index)、香农多样性(Shannon)、可观察到物种数(observed OTUs)和辛普森指数(Simpson)表示α多样性。利用unweighted UniFrac和weighted UniFrac距离进行主坐标分析和计算群落相似度。
1.2.4 抗生素抗性基因DNA的提取及含量的测定
采用Ezup土壤基因组DNA提取试剂盒(Sangon Biotech,China)提取样本抗生素抗性基因的总DNA。用分光光度计(NanoDrop ND-2000c,Thermo Fisher Scientific,USA)和1.5%琼脂凝胶电泳法测定DNA的浓度和质量。在-80 ℃保存高质量提取的总DNA,利用安徽微分基因科技有限公司的WaferGen SmartChip实时定量PCR系统对抗生素抗性基因进行高通量定量PCR(HTqPCR)检测。每100 nL反应混合物含有50 nL的1 LightCycler 480 SYBR Green I Master Mix(Roche Inc.,USA),20 nL的2 ng/mL DNA模板,1 nL的0.1 mg/mL牛血清白蛋白,500 nM的正向和反向引物,以及19 nL的无核酸酶PCR级水。对于每一个引物组,扩增是在三联体中进行的,并且包括一个非模板对照。热处理方案包括95 ℃初始变性10 min;然后在95 ℃变性30 s,60 ℃退火30 s,共循环40次,程序自动生成熔解曲线分析[30]。抗生素抗性基因的引物如表1所示。
1.3 数据处理
使用SmartChip qPCR软件(版本2.7.0.1)分析HT-qPCR的结果。分析中不包括具有多个熔化峰或扩增效率在1.8—2.2范围之外的样本。对观察显著荧光信号所需的周期数(阈值周期,Ct)进行了限制。只有Ct<31个周期且有2个以上重复出现扩增的样本才被认为是阳性。
1.4 统计分析
本研究所用的统计软件为SPSS 21.0。用单因素方差分析比较两物种间主要肠道微生物相对丰度的差异以及种间Alpha多样性的差异;用Mann-Whitney检验比较两物种间抗生素抗性基因(antibiotic resistance gene,ARGs)拷贝数的差异;通过weighted UniFrac 距离矩阵和 unweighted UniFrac 距离矩阵比较两物种的Beta多样性。使用PerMANOVA分析计算体重、性别和物种对肠道微生物群落结构的影响;使用冗余分析描述抗生素耐药基因与肠道微生物相对丰度的关系,并用Spearman秩相关系数判断抗生素抗性基因和肠道菌群的相关性,显著性水平α=0.05。

表1 抗生素抗性基因的引物
2 研究结果
2.1 高原鼠兔和高原鼢鼠肠道微生物的组成
本次研究鉴定到38个门,758个属,按97%的相似度聚类共筛选出15 814个OTU(剔除只含有1条序列的OTU或不属于细菌的OTUs),在门水平和属水平分别选择相对丰度大于1%和0.1%的菌群作为优势门和优势属,用相对丰度描述肠道菌群的组成,其结果见图1。
图1(A)是门水平高原鼠兔和高原鼢鼠肠道微生物的组成,其中主要的肠道微生物(平均相对丰度≥1%)有拟杆菌门(Bacteroidetes)(45.24%),厚壁菌门(Firmicutes)(43.92%),变形菌门(Proteobacteria)(4.47%),螺旋体门(Spirochaetes)(2.12%)和放线菌门(Actinobacteria)(1.22%);图1(B)是属水平两物种肠道微生物的组成,其主要的肠道微生物(平均相对丰度≥1%)有S24-7中某属(31.22%),梭菌目(Clostridiales)的某科某属(12.10%),普雷沃氏菌(Prevotella)(7.09%),瘤胃球菌(Ruminococcus)(6.20%),颤螺菌属(Oscillospira)(6.04%),毛螺菌科(Lachnospiraceae)(4.10%)某属,瘤球菌科(Ruminococcaceae)某属(3.09%),拟杆菌目(Bacteroidales)某科某属(2.17%),丹毒科(Erysipelotrichaceae)某属(2.09%),密螺旋体(Treponema)(1.95%),大肠杆菌(Escherichia)(1.47%)和YRC22属(1.16%)。
图2(A),(B)和(C)分别代表了在门,属和OTU水平两物种共有的和特有的微生物。本次研究中,在门水平,仅出现在高原鼠兔中的有20个门,仅出现在高原鼢鼠中的有1个门,同时出现在2个物种中的门有17个;在属水平,高原鼠兔特有的属有503个,而高原鼢鼠特有的属有54个,共有的属有201个;在OTU水平,高原鼠兔特有的OTU的个数为10 622个,高原鼢鼠特有的OTU的个数为4 287个,共有的OTU共905个。原始的16S rRNA 基因下机fastq序列已经递交到欧洲核苷酸数据(European nucleotide archive),登录号为PRJEB39107。
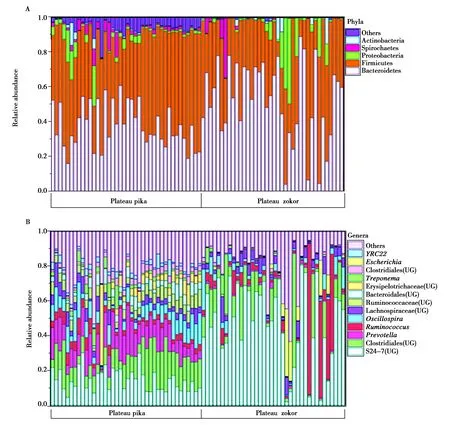
图1 高原鼠兔和高原鼢鼠肠道菌群的组成Fig.1 Composition of gut microbes of plateau pika and plateau zokor
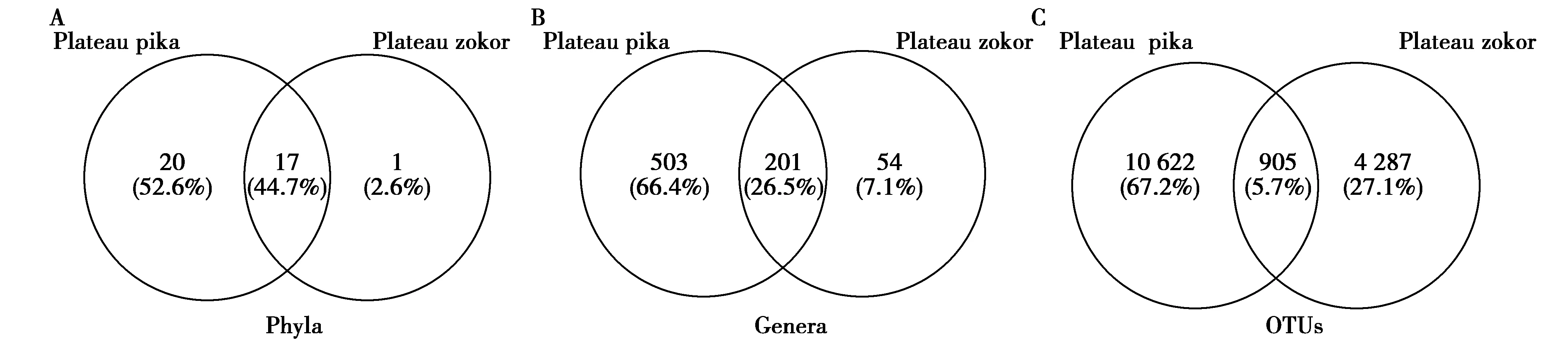
图2 高原鼠兔和高原鼢鼠共有及特有的肠道菌群种类数Fig.2 Number of species of shared and unique gut microbes in plateau pika and plateau zokor
为了探究高原鼠兔或高原鼢鼠主要的肠道菌群在两物种间是否存在差异,本研究在门(平均相对丰度≥0.1%)、属(平均相对丰度≥1%)和OTU(平均相对丰度≥0.1%)水平分别比较了两物种主要肠道微生物,找出有显著差异的肠道菌群,这些肠道菌群的相对丰度在高原鼠兔和高原鼢鼠中有显著差异。由于OTU水平下有差异的微生物类群较多,因此,仅取平均相对丰度差异最大的20个,结果见图3。在门,属和OTU水平两动物物种主要肠道菌群的差异。在门水平,高原鼢鼠的拟杆菌门的平均相对丰度更高;而高原鼠兔的厚壁菌门,放线菌门,疣微菌门(Verrucomicrobia)和软壁菌门(Tenericutes)的平均相对丰度更高。在属水平,高原鼢鼠的拟杆菌属(Bacteroides)和S24-7中某属平均相对丰度高于高原鼠兔;而高原鼠兔的普雷沃氏菌,颤螺菌属,YRC22,瘤球菌科某属,拟杆菌目某科某属和丹毒科的平均相对丰度高于高原鼢鼠。在OTU水平,高原鼠兔的OTU1_Prevotella(普雷沃氏菌),OTU2_Bacteroidetes(拟杆菌门),OTU3_Clostridiales(梭菌目),OTU4_Clostridiales(梭菌目),OTU5_Prevotella(普雷沃氏菌),OTU6_Prevotella(普雷沃氏菌),OTU7_Oscillospira(颤螺菌属),OTU8_Prevotella(普雷沃氏菌),OTU9_Clostridiales(梭菌目)、OTU10_Prevotella(普雷沃氏菌)更丰富;而高原鼢鼠的OTU11_S24-7,OTU12_Plebeius,OTU13_Bacteroidetes(拟杆菌门),OTU14_Lachnospiraceae(毛螺菌科),OTU15_S24-7,OTU16_copri,OTU17_S24-7,OTU18_S24-7,OTU19_S24-7,OTU20_Clostridiales(梭菌目)更丰富。
2.2 高原鼠兔和高原鼢鼠肠道微生物多样性比较
本次研究用Chao指数(Chao index),观察到的物种数(observed OTUs),香农指数(Shannon),辛普森指数(Simpson)4个α多样性指数描述并比较两物种的α多样性。高原鼠兔肠道菌群的α多样性显著高于高原鼢鼠,结果如图4所示。比较高原鼠兔和高原鼢鼠的β多样性,由表2可知,性别和体重对肠道微生物群落结构没有显著影响,但是物种对肠道微生物群落结构有显著影响,图5也显示高原鼠兔和高原鼢鼠群落结构有显著差异。
2.3 高原鼠兔和高原鼢鼠体内抗生素抗性基因含量
共选取高原鼢鼠和高原鼠兔各7只检测抗生素抗性基因,我们总共检测了11种抗生素基因,但是在高原鼠兔和高原鼢鼠中只检测到了6类抗生素抗性基因(图6),分别是tetQ(抗四环素类),tetM-01(抗四环素类),tetG-01(抗四环素类),sul1(抗磺胺类),sul2(抗磺胺类)和floR(多重耐药),除了tetQ的含量在两物种间有差异外,其余抗生素抗性基因在两物种间均无差异。
2.4 高原鼠兔和高原鼢鼠体内抗生素抗性基因和肠道菌群的关系
本研究分别在门(图7-A),属(图7-B)和OTU水平(图7-C)判断高原鼠兔和高原鼢鼠主要肠道微生物和抗生素抗性基因的关系(门水平:平均相对丰度≥0.1%;属水平:平均相对丰度≥1%;OTU:平均丰度≥0.1%,在两物种中平均相对丰度差别最大的10个)。其相关关系见表3,表4,表5。在门水平,tetQ与拟杆菌门(Bacteroidetes)的相对丰度呈正相关,与蓝细菌(Cyanobacteria)和酸杆菌(Acidobacteria)的相对丰度呈负相关,其余抗生素抗性基因与肠道菌群的相关性均不显著;在属水平,tetQ与普雷沃氏菌、瘤球菌科某属、拟杆菌目某属以及YRC22的相对丰度均呈负相关,sul1与毛螺菌科某属的相对丰度呈正相关,floR与颤螺菌属的相对丰度呈正相关;在OTU水平,tetQ与OTU1_Prevotella、OTU2_Bacteroidetes、OTU3_Clostridiales、OTU4_Clostridiales和OTU5_Prevotella的相对丰度呈负相关,与OTU16_copri、OTU17_S24-7、OTU18_S24-7、OTU19_S24-7和OTU20_Clostridiales的相对丰度呈正相关,tetM-01与OTU16_copri的相对丰度呈正相关。

表2 性别、体重及物种对群落结构的影响
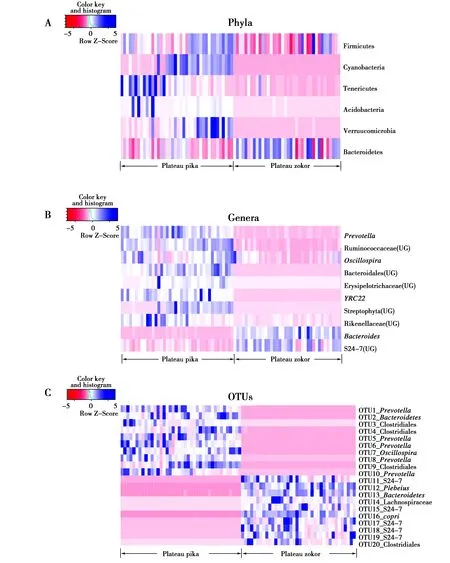
图3 主要的肠道微生物在高原鼠兔和高原鼢鼠中的差异Fig.3 Differences of main gut microbes between plateau pika and plateau zokor
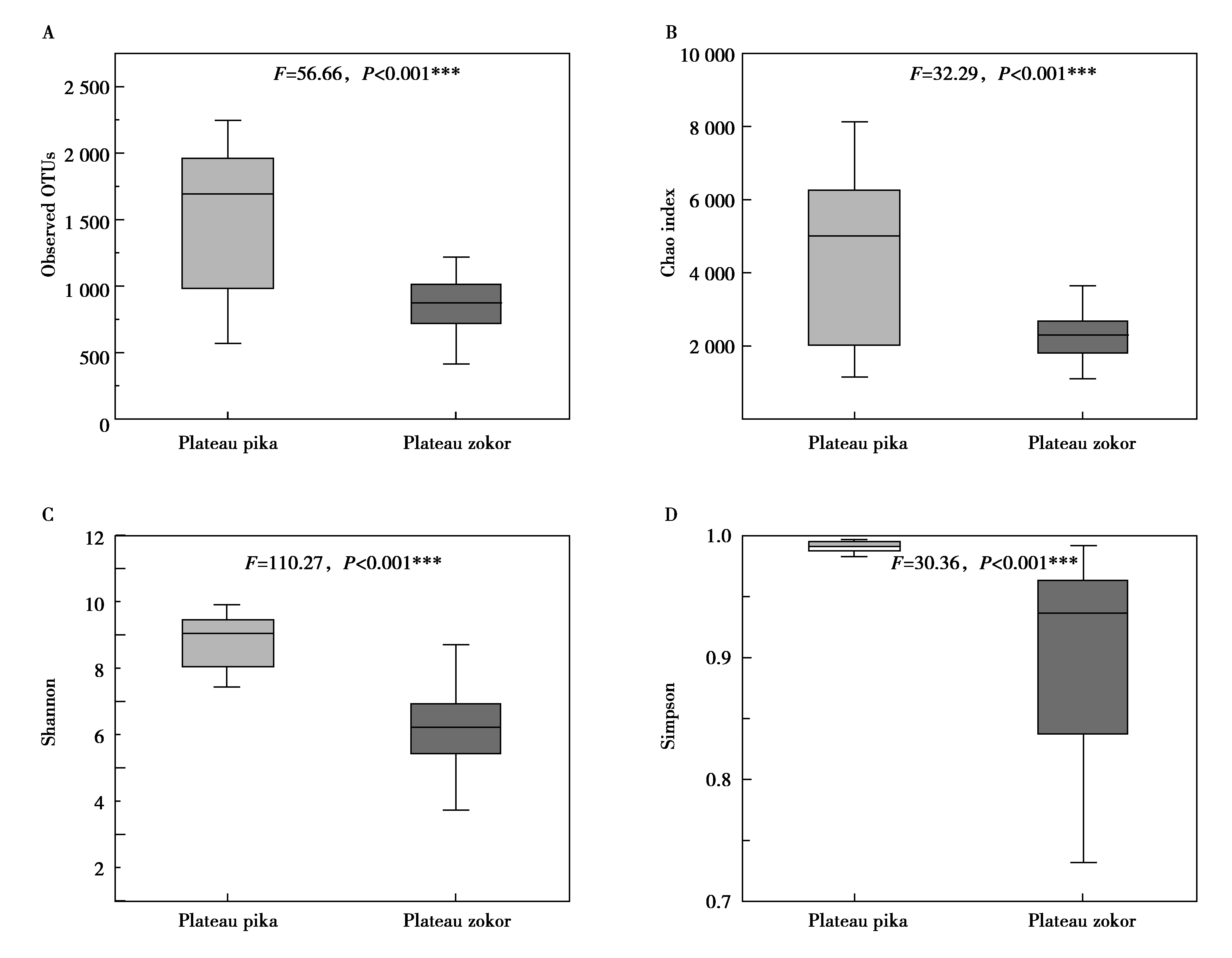
图4 高原鼠兔和高原鼢鼠肠道菌群α多样性的比较Fig.4 Comparison of gut microbes α diversity between plateau pika and plateau zokor
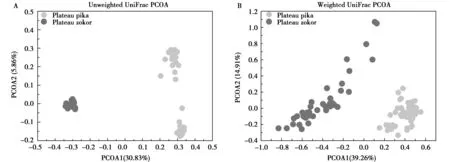
图5 高原鼠兔和高原鼢鼠肠道菌群群落结构的差异Fig.5 Differences of community structure of gut microbes between plateau pika and plateau zokor

图6 高原鼠兔和高原鼢鼠体内抗生素抗性基因的含量Fig.6 Quantity of antibiotic resistance genes in plateau pika and plateau zokor
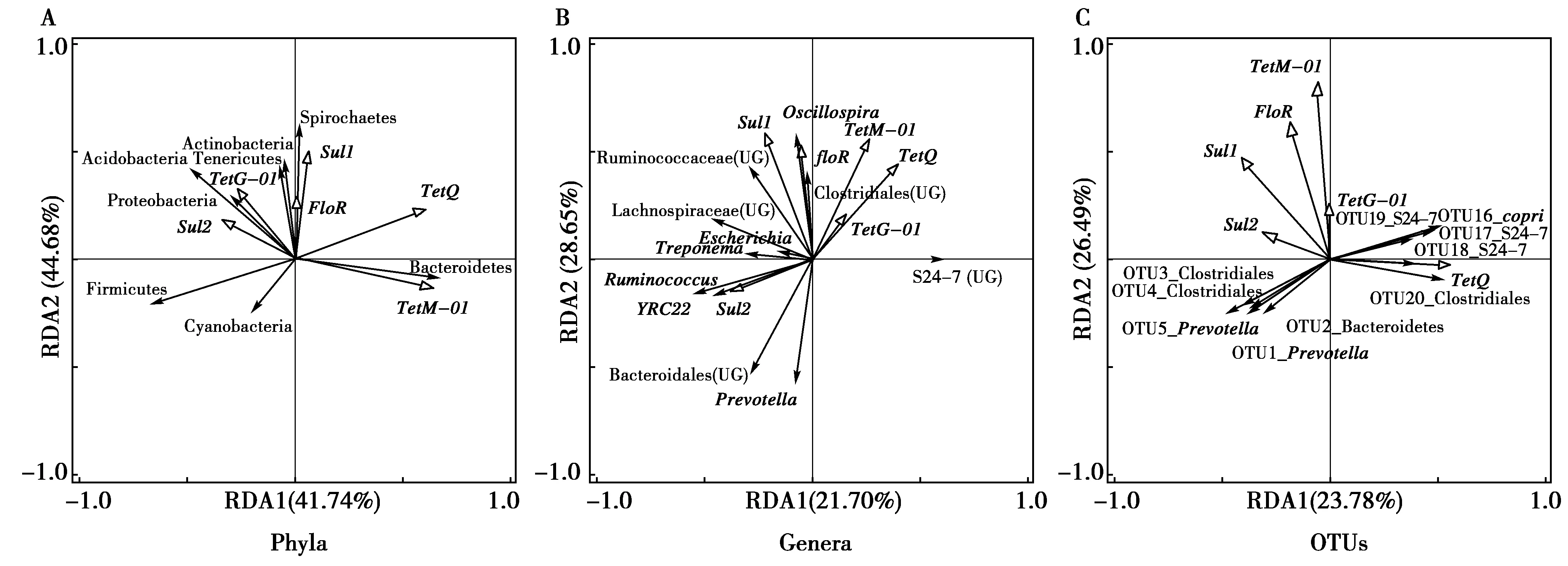
图7 高原鼠兔和高原鼢鼠肠道微生物与抗生素抗性基因的关系Fig.7 Relationship between gut microbes and antibiotic resistance genes in plateau pika and plateau zokor

表3 门水平抗生素抗性基因和肠道微生物的关系

表4 属水平抗生素抗性基因和肠道微生物的关系
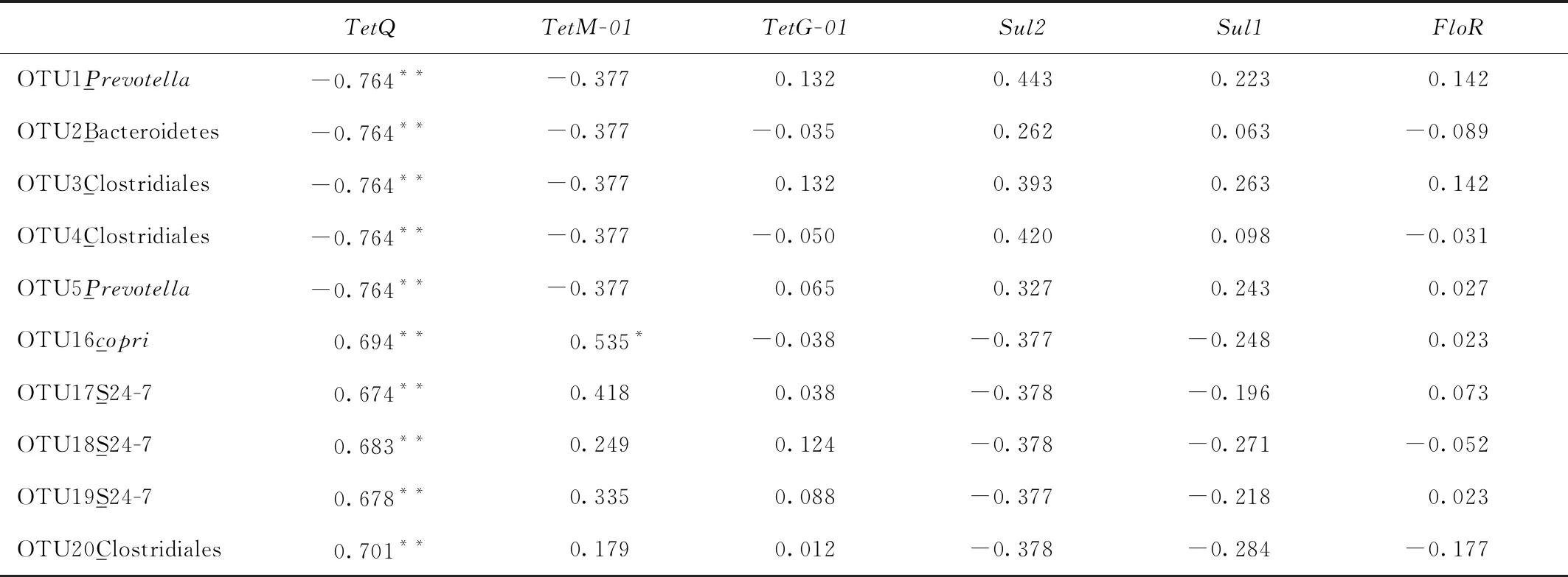
表5 OTU水平抗生素抗性基因和肠道微生物的关系
3 讨论
3.1 高原鼠兔和高原鼢鼠肠道微生物组成和多样性有差异
高原鼠兔和高原鼢鼠肠道菌群的组成在门、属和OTU水平均有差异,这一差异有可能是由二者属于不同的物种,且膳食结构和生活环境不同造成的,因为宿主的种类、膳食结构以及环境等因素都是肠道微生物结构的重要影响因素[11]。第一,高原鼠兔属于鼠兔科;而高原鼢鼠属于仓鼠科,二者属于不同的物种。第二,高原鼠兔主要栖息在地上,以植物叶、茎、花、根、芽为食,较少摄入种子;而高原鼢鼠主要栖息于地下,以植物根、皮、茎、叶、果实和种子为食,这两个物种的食物与生活的环境有差异。物种、饮食及生活环境的差异可能导致两物种肠道菌群的组成有差异。
在本次研究中,高原鼠兔和高原鼢鼠的α多样性也有显著差异,高原鼠兔肠道菌群的α多样性高于高原鼢鼠。而肠道微生物往往因宿主基因型、宿主生理状态、宿主生活方式和宿主所处的环境因素等的改变而改变[32],两物种的基因型、生理状态和生活环境的差异可能导致二者肠道微生物多样性的不同。宿主的膳食也是影响肠道微生物多样性的重要因素,食物中的营养成分可能影响肠道微生物的多样性[33-34]。另外,比起高原鼢鼠,高原鼠兔生存的外部草原环境更复杂,海拔更高,气候更加恶劣,食物更匮乏,因此可能会接触到更多的环境微生物,有可能获得更多的环境微生物定植,进而提升了肠道微生物的多样性。在其他研究中,尽管高原鼠兔食物的多样性低于达乌尔鼠兔(Ochotonadaurica),但其肠道微生物多样性却高于后者。这可能是由于高原鼠兔在高海拔地区需要更多的能量,而其肠道微生物需要更加充分地发酵简单的食物,促进高原鼠兔对高海拔环境的适应性,而肠道微生物多样性提高可以提升发酵效率[30]。同样,比起高原鼢鼠,高原鼠兔的生活环境海拔更高,因而肠道菌群的多样性更高,这可能是由于海拔影响了食物的组成[35],继而影响了肠道微生物的多样性。
高原鼠兔和高原鼢鼠的群落结构有显著差异,两物种间群落结构差异度大,但同一物种内群落结构相似度高。由于高原鼠兔和高原鼢鼠分属不同的物种,基因型不同,这一差异可能导致肠道菌群群落结构的不同[32]。如在实验室环境中,来自同一繁殖群体或供应商的幼鼠基因相似度高,其微生物群落结构比其他小鼠具有更高的相似度[36]。膳食结构也是肠道菌群群落结构的重要影响因素,在自然环境中膳食结构更相似的个体肠道菌群具有更高的相似度[35],高原鼠兔和高原鼢鼠饮食结构的差异可能是造成这一现象的原因。
3.2 部分肠道菌群同时在高原鼠兔和高原鼢鼠肠道内富集,这些微生物提高了它们消化植物性食物的能力
在不同分类水平上,高原鼠兔的肠道微生物种类比高原鼢鼠更为丰富。高原鼠兔和高原鼢鼠有一部分共有的肠道菌群,也有各自特有的微生物群。有研究发现不同动物机体内都有特定微生物群,大约70%的微生物具备种群特异性,但其丰度不超过0.5%,动物机体内的微生物群的系统发育和功能组成在出生后趋于平衡,在断奶后逐渐平稳,微生物种类和数量相对稳定。其中拟杆菌门、厚壁菌门和变形菌门3个门类属于优势菌群,也是核心微生物。它们调节了肠道菌群生物多样性,有利于肠道菌群的稳定[37]。
在门水平,厚壁菌门和拟杆菌门是高原鼠兔和高原鼢鼠的优势菌群。且甘肃鼢鼠(Eospalaxfontanieriicansus))肠道菌群在门水平相对丰度位居前二的也是厚壁菌门和拟杆菌门[38-39];而厚壁菌门和拟杆菌门也是其他植食性哺乳动物如马(Equus)[40]、驴(donkey)[41]、梅花鹿(Cervusnipponhortulorum)[42]的优势菌群。甘肃鼢鼠又称罗氏鼢鼠,属于仓鼠科,鼢鼠属,是一类长期生活在地下的杂食性动物,其食物往往是富含纤维素的植物根系[43],食性与高原鼢鼠相似;马、驴和梅花鹿均是常见的植食性动物,其食物富含纤维素。因而,纤维素的消化吸收对以上几种动物而言尤为重要。肠道中的厚壁菌门和拟杆菌门对动物消化食物中的纤维素有促进作用[42]。
在属水平,类杆菌科(Bacteroidaceae)S24-7的某属、普雷沃氏菌、瘤胃球菌以及颤螺菌属是优势菌群,S24-7分布局限,主要存在于恒温动物的肠道内,且数量上在肠道菌群中占据主导地位,且S24-7中有多种细菌可以增加降解碳水化合物酶的相对含量[44]。高原鼢鼠和高原鼠兔均属于恒温动物,且它们的食物中富含纤维素等碳水化合物,因而S24-7平均相对丰度高。普雷沃氏菌和瘤胃球菌均属于瘤胃纤维性分解菌[45],它们往往拥有不同的纤维素酶和半纤维素酶基因[46],帮助宿主消化植物纤维素并从中汲取能量。其中普雷沃氏菌往往不能直接消化纤维素,但在低pH下可以降解纤维素衍生物羧甲基纤维素,同时,普雷沃氏菌催化结构域能和来自热单孢菌(Thermomonsporafusca)的结合结构域的重组酶结合,并拥有至少10倍的纤维素酶活性[47]。而颤螺菌属则在纤维素发酵过程中也具有一定作用[48]。由于高原鼠兔和高原鼢鼠食物中的主要成分为纤维素,因而以上几类细菌有较高的相对丰度。
3.3 在高原鼠兔和高原鼢鼠体内均检出抗生素抗性基因
本研究在高原鼠兔和高原鼢鼠体内检测到抗生素抗性基因共6种,其中四环素类抗性基因3种,分别是tetQ,tetM-01和tetG-01;磺胺类的抗性基因2种,分别是sul1和sul2;还有一类多重耐药基因floR。动物可以由自身肠道微生物合成抗生素抗性基因,比如动物体内产生抗生素的细菌可以编码抗生素抗性基因,以保护它们免受自身产生的生物活性分子的影响,抗生素耐药基因产生后不可避免地在病原菌中传播[49]。动物也可以从外界环境中获得抗生素抗性基因,由于抗生素在畜牧业和养殖业中的长期滥用,在养殖动物肠道内诱导出可以编码抗生素抗性基因的抗性菌株,这些菌株随粪便排泄,再经雨水冲刷,地表径流和大气扩散等多种途径进入到环境中并被动物摄取,这些编码抗生素抗性基因的菌株是环境中抗生素抗性基因最重要的来源。抗生素抗性基因可以通过质粒的水平交换从一种细菌转移到人类或家畜微生物群[50-51],因此,高原鼠兔和高原鼢鼠体内的抗生素抗性基因可能源自以上两种途径。
3.4 肠道微生物和抗生素抗性基因的关系
有研究发现,抗生素抗性基因普遍存在于农场动物的肠道微生物中[52-53],而高原鼠兔和高原鼢鼠与牦牛相互食粪的行为可能导致其肠道菌群中存在抗生素抗性基因。动物微生物群中的耐药细菌因此可以作为抗生素抗性基因重要的贮存库[54]。而这些病原菌可以通过水平基因转移或交换质粒将抗生素抗性基因转移到肠道菌群中使肠道菌群携带抗生素抗性基因,有研究表明人类肠道中大量的微生物充当着抗生素抗性基因的储存库[55],因此高原鼠兔和高原鼢鼠的肠道微生物也有可能成为抗生素抗性基因的贮存库。
拟杆菌、颤螺菌属等肠道菌群的相对丰度与抗生素抗性基因呈正相关,它们有可能是抗生素抗性基因的贮存库,或者与携带抗生素抗性基因的病原微生物有协同作用,而蓝细菌、放线菌、普雷沃氏菌、瘤球菌科的某属、拟杆菌目的某属YRC22等肠道微生物的相对丰度与抗生素抗性基因呈负相关,这些肠道菌群则可能与携带抗生素抗性基因的病原菌相互拮抗,阻止了抗生素抗性基因的积累。通过调节高原鼠兔和高原鼢鼠的肠道微生物,比如适当减少拟杆菌的相对含量或增加放线菌的相对含量,可能阻止抗生素抗性基因的富集与传播,降低抗生素抗性基因向人类传播的机会,减少抗生素抗性基因对人体的危害。
致谢:感谢实验室各位老师和同学在实验和数据分析上提供的帮助。
猜你喜欢
当代水产(2022年7期)2022-09-20
今日农业(2022年14期)2022-09-15
中老年保健(2022年2期)2022-08-24
中国饲料(2022年5期)2022-04-26
昆明医科大学学报(2022年3期)2022-04-19
中国饲料(2021年17期)2021-11-02
新疆农业科学(2020年9期)2020-10-14
家教世界·创新阅读(2020年4期)2020-06-03
当代工人·精品C(2020年1期)2020-05-20
家教世界·创新阅读(2020年1期)2020-05-11
