
胡椒瘟病菌Phytophthoracapsici孢子囊诱导及发育过程观察
2020-10-29 07:35高圣风杨开虎陆大倩刘爱勤苟亚峰孙世伟王政孟倩倩
热带作物学报 2020年9期
高圣风 杨开虎 陆大倩 刘爱勤 苟亚峰 孙世伟 王政 孟倩倩
摘 要:孢子囊的产生和发育过程是研究胡椒瘟病菌(Phytophthora capsici)及其病害防控的重要基础。本研究分析不同诱导条件下胡椒瘟病菌孢子囊的产生情况,并通过光学显微镜和共聚焦显微镜观察孢子囊及其内部游动孢子的发育过程。结果发现:(1)3种诱导方法均能诱导出大量孢子囊,其中在V8-A平板上光照和抹伤双重诱导法获得的孢子囊数量最多,其次是在V8-A平板上光照诱导法,获得孢子囊数量最少的是V8液体光照诱导法,但仅差异最大的2个处理间达到显著水平;(2)显微镜观察发现,孢子囊由气生菌丝顶端逐步膨大形成,初始为近球形逐渐发育成倒洋梨形,孢子囊成熟后从顶端排出大量游动孢子,偶尔可见孢子囊顶端直接发育出芽管;(3)共聚焦显微镜观察发现,首先孢子囊内部原生质体被膜结构隔裂成大约数十个独立小格,然后在每个格子中积累数倍于细胞核的DNA,最后每份细胞核DNA发育成一个游动孢子。本研究从微观角度揭示P. capsici孢子囊发育外观及内部的形态特征,为胡椒瘟病菌后續致病机制研究和胡椒瘟病田间防控提供技术基础。
关键词:胡椒瘟病;辣椒疫霉;孢子囊诱导;发育过程
中图分类号:S432.1 文献标识码:A
Abstract: Sporangium is an important basis for the Phytophthora foot rot disease control and the pathogen (Phytophthora capsici) research in black pepper, but the sporangium-inducing methods and the micro development processes have not been reported yet. Three sporangium-inducing methods were tested and the sporangia development processes were observed by light microscope and laser confocal microscope. Abundant sporangia could be detected by all the three methods. The most sporangia were obtained by the method of lighting and glass rod-plastering induction on V8-A plates, followed by the lighting induction on V8-A plates and lighting induction in V8 liquid. Under light microscope, sporangia initially formed at the top of the hypha with the shape of small sphere, and gradually formed into an inverted pear shape. When matured, the sporangia released a large number of zoospores from an apical pore. Interestingly, a few sporangia germinated directly and formed a germ tube. Under laser confocal microscope, the protoplast of a sporangium was separated into dozens of small cells by membrane structure at first, and then multiple nuclear DNA was accumulated in each cell, and finally the multinuclear cell developed into several mononuclear zoospores. The study revealed the morphological characteristics of the P. capsici sporangium development progress would provide sporangium-inducing methods for further pathogenic mechanisms research and field disease management.
Keywords: black pepper phytophthora foot rot; Phytophthora capsici; sporangia induction; development process
DOI: 10.3969/j.issn.1000-2561.2020.09.021
胡椒(Piper nigrum Linn.)是一种经济价值极高的食品香料作物,是海南省第三大经济作物。胡椒瘟病具有极强的毁灭性和传播性,在20世纪70年代曾导致中国胡椒种植面积缩减20%,其病原物是辣椒疫霉菌(Phytophthora capsici)[1-2]。2009—2010年对我国胡椒主产区海南省胡椒病害发生情况调查发现,胡椒瘟病在海南省11个胡椒主要种植市(县)均普遍发生、危害严重[3-4],而且最近几年胡椒瘟病呈现蔓延加重趋势,是影响胡椒产业稳定发展的重要瓶颈之一。
孢子囊和游动孢子是P. capsici的再侵染源,可以通过气流和水流快速传播,造成病害蔓延流行。孢子囊的产生不仅是P. capsici在田间发生流行的关键前提,也是实验室科学研究的重要基础,尤其在侵染致病、遗传操作等相关研究上至关重要。在国外,P. capsici的孢子囊诱导技术报道较多,主要是在V8蔬菜汁培养基上黑暗培养后光照诱导孢子囊产生[5-7]。在国内,除了V8蔬菜汁培养基,黑麦培养基、大豆培养基、胡萝卜培养基等也被用于诱导P. capsici孢子囊[8-11],其诱导条件也不仅限于单独光照,兰成忠等[12]在光照诱导的基础上提出了抹伤诱导;但是未见研究胡椒瘟病病原菌孢子囊诱导技术的相关报道。受限于孢子囊诱导技术,目前胡椒瘟病的致病性接种也主要采用菌丝块伤口接种方法[13-14],尚未使用与P. capsici自然侵染一致的游动孢子接种法[6]。国内外通过光学显微镜、电子显微镜等手段研究发现P. capsici的孢子囊形态主要是倒洋梨形,也常见卵形、长卵形、球形、不规则形等性状[15-18],但是对其孢子囊发育及其内部游动孢子发育过程尚未见报道。
本研究在前人研究的基础上,分析不同诱导条件下胡椒瘟病菌孢子囊的产生情况,并通过光学显微镜和共聚焦显微镜进一步观察孢子囊和游动孢子的发育过程,从微观角度揭示胡椒瘟病菌的形态特征,为胡椒瘟病致病机制等相关研究提供基础。
1 材料与方法
1.1 材料
1.1.1 供试菌株 胡椒瘟病菌由本研究室从胡椒病株中分离纯化得到,于马铃薯葡萄糖琼脂(PDA)试管斜面培养基中室温保存。
1.1.2 培养基 胡椒瘟病菌保藏及活化使用PDA培养基:马铃薯200 g,切成小块,在800 mL去离子水中煮沸15 min,用双层纱布过滤,滤液中加入葡萄糖和琼脂粉各20 g,用去离子水定容至1000 mL,121 ℃灭菌20 min[2]。
胡椒瘟病菌孢子囊诱导培养基:(1)10% V8蔬菜汁(V8)培养基,离心后的V8蔬菜汁100 mL, CaCO3 1g,用去离子水定容至1000 mL,121 ℃灭菌20 min[7];(2)10% V8蔬菜汁琼脂(V8-A)培养基,配制方法与V8培养基基本相同,仅在灭菌前添加琼脂粉20 g/L[7]。
1.1.3 荧光染料 激光扫描共聚焦显微观察使用2种荧光染料:(1)4,6-二脒基-2-苯基吲哚(4,6-diamidino-2-phenylindole, DAPI),能够与双链DNA结合,在波长330~405 nm的紫外光下激发出波长461 nm的蓝光;(2)苯乙烯染料(N-(3- Triethylammoniumpropyl)-4-(6-(4-(Diethylamino) phenyl) hexatrienyl) Pyridinium Dibromide, FM?4- 64FX),能夠结合细胞质膜,在波长510~570 nm的绿光下激发出波长734 nm的红光,2种染料均购自赛默飞世尔科技(中国)有限公司。
1.2 方法
1.2.1 胡椒瘟病菌孢子囊诱导 从保藏的试管斜面中挑取少量菌种转接入PDA培养基中,于28 ℃恒温培养箱中培养2 d,再次转接入新的PDA平板中,于28 ℃恒温培养箱中培养5 d,作为以下孢子囊诱导方法的出发菌种。
V8液体培养光照诱导:将胡椒瘟病菌接种在V8-A平板上,置于28 ℃恒温培养箱中培养7 d,切成直径约1 cm的菌丝块浸入V8液体培养基中,28 ℃条件下暗培养7 d后,挑取菌丛用无菌去离子水清洗后浸入无菌去离子水中,在28 ℃条件下光照培养,24 h后开始取样,每隔12 h取样1次,做成临时装片,用光学显微镜检测孢子囊数量。
V8-A平板培养光照诱导:将胡椒瘟病菌接种在V8-A平板上,置于28 ℃恒温培养箱中暗培养7 d,然后在28 ℃条件下光照培养,24 h后开始取样,每隔12 h取样1次,用光学显微镜检测孢子囊数量。
V8-A平板培养光照和抹伤双重诱导:将胡椒瘟病菌接种在V8-A平板上,置于28 ℃光照生化培养箱中培养7 d,用无菌玻璃棒涂抹菌丝后再次置于28 ℃光照生化培养箱中培养,24 h后开始取样,每隔12 h取样1次,用光学显微镜检测孢子囊数量。
1.2.2 孢子囊光学显微镜观察 孢子囊数量分析:在载玻片上滴加无菌去离子水15 μL,挑取少量菌丝平铺于载玻片上,盖上盖玻片后置于奥林巴斯IX71倒置式显微镜载物台上,用200倍视野进行观察。每个处理设置3个样本重复,每个样本制作5个临时装片,镜检时选取菌丝分布均匀一致的视野利用cellSens Dimension软件统计单位面积(mm2)中的孢子囊数量。
孢子囊发育过程观察:在Φ15mm玻底培养皿中加入无菌去离子水50 μL,挑取少量菌丝放入培养皿中使菌丝平铺于玻底,四周放置吸足无菌水的脱脂棉保湿,盖上盖子后置于奥林巴斯IX71倒置式显微镜载物台上,用400倍视野利用cellSens Dimension软件进行观察并采集图像,每20 min采集一次,直至孢子囊成熟或游动孢子释放。
1.2.3 孢子囊染色及激光扫描共聚焦显微镜观察 细胞核酸的染色及其观察:DAPI染料用无菌去离子水配制成终浓度为10 mmol/L的母液,避光保存于-20 ℃冰箱中。将DAPI母液和无菌去离子水按照1∶5000体积稀释配置成染色液,染色液现配现用。挑取少量待检测菌丝浸入染色液中5 min,在无菌水中清洗3次后制成临时装片,置于Olympus Flv10智能激光扫描共聚焦显微镜的载物台上。选取波长330~405 nm的激发光进行显微观察,使用Olympus Fluoview V4.2软件采集图像。
细胞膜染色及其观察:FM?4-64FX染料用无菌去离子水配制成终浓度为164 μmol/L的母液,避光保存于-20 ℃冰箱中。将FM?4-64FX母液和无菌去离子水按照1∶10体积稀释配置成染色液,染色液现配现用。挑取少量待检测菌丝浸入染色液中1 min,在无菌水中清洗3次后制成临时装片,置于Olympus Flv10智能激光扫描共聚焦显微镜的载物台上。选取波长510~570 nm的激发光进行显微观察,使用Olympus Fluoview V4.2软件采集图像。
1.3 数据处理
采用SPSS 16.0(SPSS Inc., Chicago, IL, USA)软件进行数据处理,采用单因素方差分析(one- way ANOVA),费雪最小显著差异检验(Fishers least-significant difference test, LSD)(P<0.05)。
2 结果与分析
2.1 孢子囊诱导效果分析
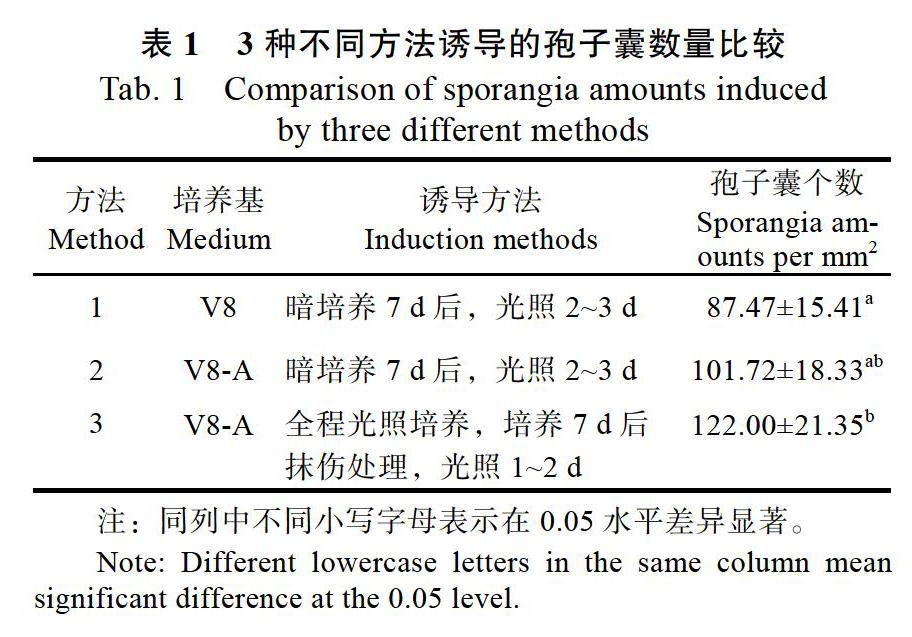
分别采用3种不同的方法开展胡椒瘟病菌孢子囊诱导试验。显微镜下检测发现,3种诱导方法均能将胡椒瘟病菌诱导出大量孢子囊;其中V8-A平板培养光照和抹伤双重诱导方法(方法3)获得的孢子囊数量最多,其次是V8-A平板培养光照诱导方法(方法2)和V8液体培养光照诱导方法(方法1),但是仅方法1和方法3之间有显著性差异(P<0.05),其他处理间差异均不显著(表1)。
2.2 孢子囊形态及其发育过程观察
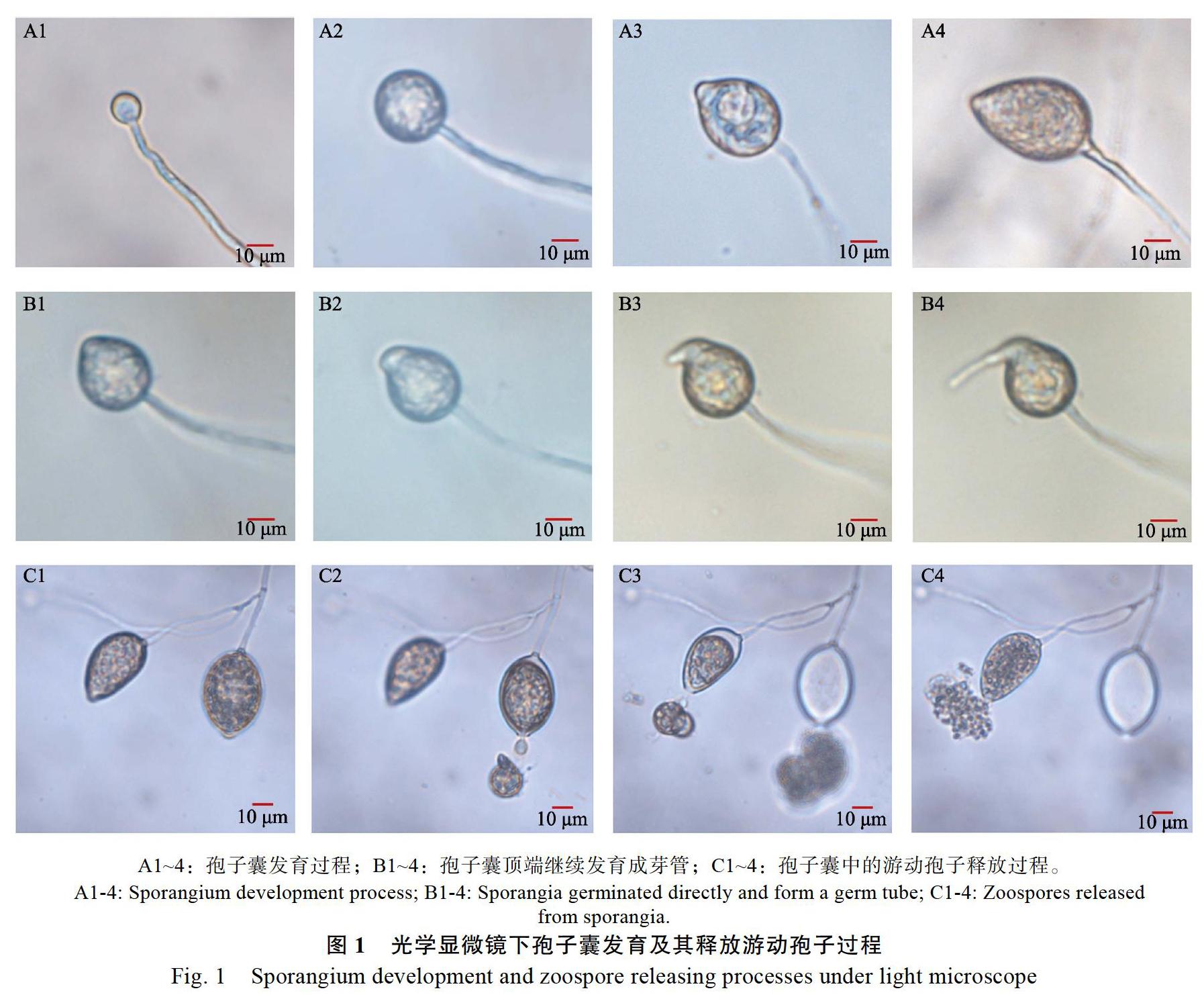
光学显微镜下观察孢子囊的发育过程发现,孢子囊产生于其菌丝顶端,初始为圆球状并逐渐膨大发育成倒洋梨形的孢子囊(图1A1~A4);偶尔可见一些孢子囊顶端再次发生形态变化,在排孢孔处向外延伸逐渐发育成形状类似菌丝的芽管(图1B1~B4);孢子囊发育成熟后从顶端排孢孔中排出游动孢子,游动孢子数量达上百个甚至数百个,刚释放的游动孢子初始拥挤成团,然后逐渐向四周分散(图1C1~C4)。
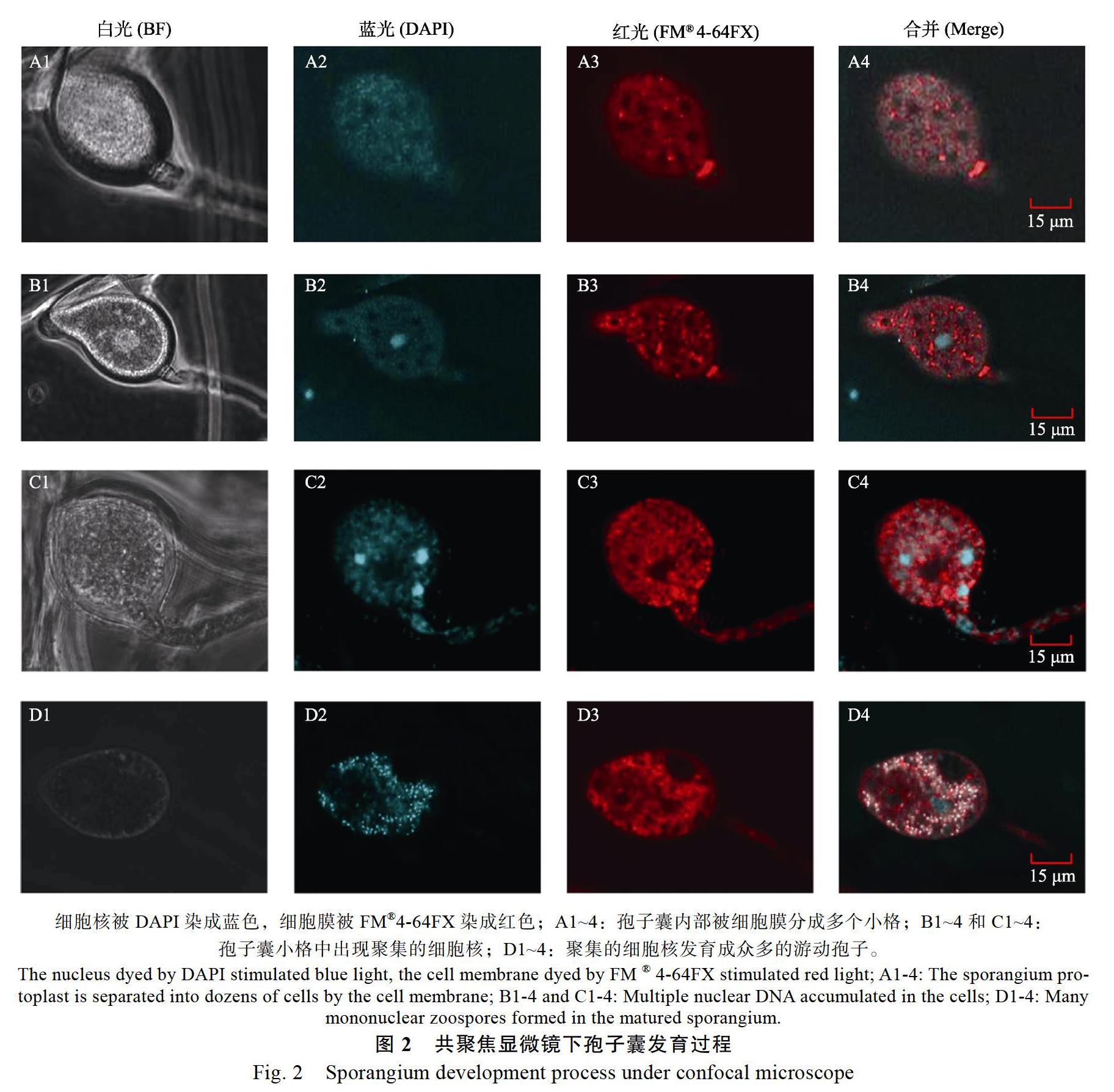
经过DAPI和FM?4-64FX 2种荧光染料染色后,在智能激光扫描共聚焦显微镜下可以看到细胞核和细胞膜分别被染成蓝色和红色(图2)。孢子囊形成初期,其内部原生质体被膜结构隔裂成大约数十个彼此独立的小格,小格呈蜂巢状排列(图2A1~A4);然后,在格子中逐渐累积大量DNA,形成一个体积数倍于细胞核的DNA团(图2B1~B4,C1~C4);小格内DNA团分散并发育成单核的游动孢子,每个孢子囊内游动孢子数量达上百个甚至数百个(图2D1~D4)。
3 讨论
众所周知,常规培养条件难以获得大量的P. capsici孢子囊。近年来,P. capsici孢子囊诱导技术日渐成熟,已通过文献报道或者以专利形式公布了各种方法[8-12]。但是国内外研究的P. capsici菌株主要来源于温带的辣椒、茄子等作物,与来源于热带的胡椒瘟病菌在遗传学系统发育树上归属不同的分类亚群[19]。本研究将辣椒疫霉孢子囊诱导方式简单归为3类,即液体培养基中光照诱导、固体平板上光照诱导和固体平板上光照、抹伤双重诱导(表1)。研究发现使用V8或V8-A培养基培养时,3种诱导方法均可以获得大量的孢子囊,说明来源于热带的胡椒瘟病菌与温带的P. capsici在孢子囊诱导条件上差异不大。3种方法诱导的孢子囊数量分别为87.47、101.72和122.00个/mm2,总体差距不大,可约略视为同一水平;结合操作难易程度和易污染程度,建议开展遗传操作等研究时采用不易污染的液体培养基光照诱导方法,开展致病性接种等研究可采用易操作的其他2种方法。此外,在显微观察过程中细胞核被DAPI染成蓝色,细胞膜被FM?4-64FX染成红色;A1~4:孢子囊内部被细胞膜分成多个小格;B1~4和C1~4:孢子囊小格中出现聚集的细胞核;D1~4:聚集的细胞核发育成众多的游动孢子。
发现,同一个玻片样本中的孢子囊发育进程并不完全一致,会出现不同发育阶段的孢子囊混杂的现象(尚未发表);结合多次镜检结果,初步认为这3种方法中双重诱导法的孢子囊发育进程最整齐,但该结果还需要更多统计数据的支撑。
光学显微镜观察发现胡椒瘟病菌的孢子囊形态主要是倒洋梨形,少数为长圆形、不规则形等(图1),与前人报道结果一致[15-17],但张开明等[18]报道的多数是舟型、少数为梨形或不规则形。本研究观察发现极少量的胡椒瘟病菌可以在孢子囊顶端发育出菌丝状结构,在其他P. capsici菌株上尚未发现类似报道;但是在同属的致病疫霉(P. infestans)、德雷疫霉(P. drechsleri)中均有孢子囊直接萌发芽管的报道[20-21],因此本研究将该菌丝状结构暂定为芽管。但是,该结构是否像孢子萌发的芽管一样可以直接侵染植株,还需要更加深入的研究。
通过激光扫描共聚焦显微镜观察发现,胡椒瘟病菌孢子囊内部被膜结构分隔成数十个小格,然后在格子中累积成数倍于细胞核的DNA团,最后DNA团分散并发育成众多游动孢子(图2)。该结果在P. capsici中尚未见类似报道。陶恺报道的大豆疫霉(P. sojae)孢子囊内部同样是先由细胞膜将原生质体分隔成数十个彼此独立空间;而不同的是,每个独立空间只有一个细胞核,发育成单个游动孢子[22]。通过反复观察胡椒瘟病菌的孢子囊及其游动孢子的释放过程,发现每个孢子囊至少能释放出约200个游动孢子,远大于其膜结构分隔出的独立空间个数,认为胡椒瘟病菌的游动孢子发育过程与大豆疫霉可能存在差异(尚未发表)。但是,是否所有P. capsici的游动孢子发育过程均与胡椒瘟病菌一致,还需要后续更多的研究。
对于胡椒瘟病菌孢子囊发育的时间周期,本研究已做一些探索。通过显微镜对视野中的特定菌丝进行长时间定位监测发现,从菌丝末端膨大到发育成一个倒洋梨形的孢子囊大约需要8~12 h;但是完整监测一个孢子囊的发育全程难度极大,成功的样本数量不多,该结果还需要更多数据支撑。另外,监测到大量的孢子囊释放过程,发现该过程较为短暂,从游动孢子开始释放到孢子囊放空仅需要大约15~30 s;但是该过程受环境温度变化影响较大,迅速降温可将该过程缩短至10 s以内(尚未发表)。对于显微镜观察结果,值得注意的是,显微镜观测环境与孢子囊诱导环境、自然环境之间的差异可能影响观察结果。但是生动具体的微观观察结果对认识微生物形态变化都具有重要的指导意义。
参考文献
[1] 刘爱勤. 热带特色香料饮料作物主要病虫害防治图谱[M]. 北京: 中国农业出版社, 2013: 2-12.
[2] 桑利伟, 刘爱勤, 谭乐和, 等. 海南省胡椒瘟病病原鉴定及发生规律[J]. 植物保護, 2011, 37(6): 168-171.
[3] 桑利伟, 谭乐和, 刘爱勤, 等. 海南省胡椒主要病害现状初步调查[J]. 植物保护, 2010, 36(5): 133-137, 148.
[4] 桑利伟, 刘爱勤, 谭乐和, 等. 胡椒瘟病田间发生规律观察[J]. 热带作物学报, 2010, 31(11): 1996-1999.
[5] Lewis Ivey M L, Miller S A. Use of the vital stain FUN-1 indicates viability of Phytophthora capsici propagules and can be used to predict maximum zoospore production[J]. Mycologia, 2014, 106(2): 362-367.
[6] Kim M J, Shim C K, Kim Y K, et al. Evaluation of watermelon germplasm for resistance to phytophthora blight caused by Phytophthora capsici[J]. The Plant Pathology Journal, 2013, 29(1): 87-92.
[7] Hord M J. Effects of physical and chemical factors on the germination of oospores of Phytophthora capsici in vitro[J]. Phytopathology, 1991, 81(12):1541-1546.
[8] 许亚池, 王述彬, 刁卫平, 等. 一种便捷高产的辣椒疫霉菌游动孢子产生方法[J]. 江苏农业科学, 2015, 43(10): 172-173, 258.
[9] 张 荣, 辛光云, 刘爱媛. 辣椒疫霉菌保存及游动孢子诱导技术研究[J]. 安徽农业科学, 2008(20): 8679-8680.
[10] 王晓敏, 巩振辉, 逯红栋, 等. 辣椒疫霉菌孢子诱导技术研究[J]. 西北农业学报, 2006, 15(2): 59-62.
[11] 兰成忠, 刘裴清, 李本金, 等. 辣椒疫霉菌产孢培养基及诱导方法筛选[J]. 热带作物学报, 2013, 34(9): 1776-1780.
[12] 兰成忠, 陈庆河, 李本金, 等. 一种简便、高效诱导辣椒疫霉菌产生游动孢子的方法[P]. 中国: CN102391980A. 2012-03- 28.
[13] 高圣风, 刘爱勤, 桑利伟, 等. 枯草芽胞杆菌VD18R19在胡椒上的定殖动态及促生作用和对胡椒瘟病的防治效果[J]. 中国生物防治学报, 2017, 33(5): 650-657.
[14] 桑利伟, 刘爱勤, 高圣风, 等. 防治胡椒瘟病生物农药筛选及其盆栽药效试验[J]. 热带作物学报, 2017, 38(5): 915-918.
[15] Williams W T, Webster R K. Electron microscopy of the sporangium of Phytophthora capsici[J]. Canadian Journal of Botany, 1970, 48(2): 221-227.
[16] Aragaki M, Uchida J Y. Morphological distinctions between Phytophthora capsici and P. tropicalis sp. nov.[J]. Mycologia, 2001, 93(1): 137-145.
[17] 冯俊涛, 韩立荣, 范瑞娟, 等. 枯茗酸对辣椒疫霉病菌生长发育的影响[J]. 中国农业科学, 2012, 45(13): 2628-2635.
[18] 张开明, 郑服丛, 黎乙东, 等. 中国胡椒疫霉种及交配型的研究[J]. 热带作物学报, 1991(2): 69-76.
[19] Bowers J H, Martin F N, Tooley P W, et al. Genetic and morphological diversity of temperate and tropical isolates of Phytophthora capsici[J]. Phytopathology, 2007, 97(4): 492-503.
[20] 唐德志, 孙毓彬, 何苏琴. 德雷疫霉侵染甜瓜叶片过程的研究[J]. 西北农业学报, 1992(1): 89-91.
[21] Fry W. Phytophthora infestans: the plant (and R gene) destroyer[J]. Molecular Plant Pathology, 2008, 9(3): 385-402.
[22] 陶 愷. 大豆疫霉侵染过程的分子细胞学研究[D]. 南京: 南京农业大学, 2012.
