
烟粉虱MED隐种BtβNAC基因克隆及表达分析
2020-10-31 02:06杜红旭王放放刘旭白润娥
河南农业大学学报 2020年5期
杜红旭,王放放,刘旭,白润娥
(1.河南农业大学农学院,河南 郑州 450002;2.河南农业大学植物保护学院,河南 郑州 450002)
生物体内新生蛋白多肽的合成受到严格调控,其中,新生多肽偶联合体(Nascent polypeptide associated complex,NAC)是与核糖体紧密结合,防止新生多肽与细胞内蛋白异常结合的功能蛋白复合体[1-4]。NAC属于高丰度细胞质蛋白,广泛存在于哺乳动物、酵母和植物细胞内[5]。在生物体内,NAC蛋白是由ɑ和β亚基构成的异二聚体,两类亚基均有各自的功能[6]。ɑ亚基具有转录激活活性,以及与DNA、rRNA和tRNA结合的活性;β亚基亦称为即通用转录因子3(Btf3),该亚基虽然缺乏转录激活的功能,但能与RNA聚合酶II相结合[5,7]。酿酒酵母的ɑ和β亚基均与新生蛋白多肽直接偶联,但是只有β亚基与核糖体相结合[7]。现有研究表明,NAC蛋白β亚基的功能主要包括:(1)促进核糖体与线粒体互作,确保延胡索酸酶和苹果酸脱氢酶等线粒体蛋白保持正确的靶向[5];(2)为转运至内质网和线粒体的新生多肽提供保护性内环境[8-10];(3)与新生多肽互作,辅助其形成正确折叠,发挥分子伴侣功能[11-12]。在β-NAC蛋白缺失的秀丽线虫体内,编码核蛋白、细胞溶质蛋白和线粒体蛋白的mRNAs被错误地靶向引导至内质网膜,并且秀丽线虫的平均寿命缩短了10 d[13]。此外,β亚基(BTF3)与哺乳动物的细胞凋亡密切相关[14-16];秀丽线虫细胞凋亡抑制因子ICD(NAC蛋白β亚基)研究表明,过量表达ICD可以阻止细胞凋亡,RNAi干扰ICD的表达导致线虫细胞的异常凋亡[17-19];酵母β亚基参与转录调控和线粒体蛋白的输出[11,20-21]。NAC蛋白的β亚基已在人类、哺乳动物、酵母和秀丽线虫,以及拟南芥[22-23]、烟草[24]、水稻[25]和小麦[26]等植物中有研究报道,尚未见到关于烟粉虱NAC蛋白的功能及其编码基因的研究报道。本研究对烟粉虱MED隐种NAC蛋白β亚基编码基因的序列和特征,以及该基因在烟粉虱不同发育阶段和吡虫啉胁迫条件下的表达情况进行了研究,期望为进一步研究昆虫NAC蛋白的功能及化学防治提供一定的理论依据。
1 材料与方法
1.1 供试昆虫与试剂
1.1.1 供试材料 烟粉虱MED隐种(寄主植物为烟草Nicotianatabacum)饲养于河南农业大学植物保护学院化学生态学实验室。
1.1.2 试剂及仪器 pGM-T和pMAL-c2x载体质粒为河南农业大学植物保护学院化学生态学实验室保存;感受态细胞Trans1-T1、Top Green qPCR SuperMix荧光定量试剂盒购置于TransGen Biotech(北京)公司;BL21感受态细胞、FastKing RT Kit (With gDNase) 购置于TIANGEN(北京)公司;总RNA提取试剂Trizol、PrimerSTAR Max购置于TaKaRa(大连)公司;限制性核酸内切酶EcoRI、SalI等购置于NEB(北京)公司;试验所用引物均自行设计,并由尚亚生物技术有限公司(郑州)合成。
1.2 烟粉虱MED隐种NAC蛋白β亚基基因的克隆
1.2.1 总RNA提取和cDNA第一链合成 用Trizol法提取健康烟粉虱成虫的总RNA,后用FastKing RT Kit (With gDNase)试剂盒操作方法合成cDNA第一条链。
1.2.2 NAC蛋白β亚基基因克隆引物设计 根据蚂蚁来源的NAC蛋白β亚基的氨基酸序列[27],搜索GenBank的EST数据库,预测拼接获得一个包含完整开放读码框的烟粉虱NAC蛋白β亚基基因的cDNA序列并设计克隆引物(表1)。
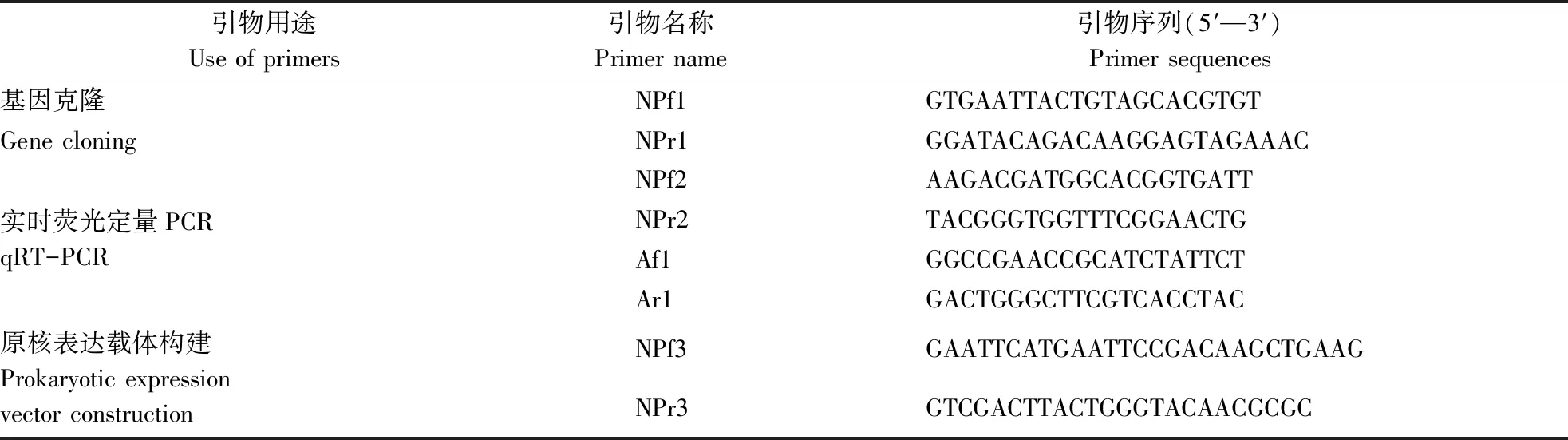
表1 烟粉虱BtβNAC基因引物Table 1 Primers used for the isolation and expression of the BtβNAC gene in Bemisia tabaci MED
1.2.3 cDNA与基因组DNA克隆 以cDNA第一条链为模板,利用Prime STARMax DNA Polymerase(TaKaRa,大连)进行PCR扩增,获得cDNA序列;以CTAB法提取的烟粉虱成虫DNA为模板,进行基因组DNA克隆,两者克隆产物由尚亚生物技术有限公司(郑州)完成测序验证后,提交NCBI GenBank获得登录号。
1.3 序列分析
利用SnapGene进行序列分析,利用Clustal×1.83和Boxshade 软件包对包括烟粉虱在内的βNAC蛋白的氨基酸进行序列联配和聚类分析。
1.4 烟粉虱β亚基mRNA表达谱分析
1.4.1 材料处理及取样 分别取处于不同发育期(卵、1龄、2龄、3龄、伪蛹和成虫)的烟粉虱100头,3次生物学重复,用液氮速冻后提取RNA。
1.4.2 药剂吡虫啉喷施处理 随机选取发育良好一致的烟粉虱成虫,接种于5叶期的烟草幼苗叶片,经过24 h适应生长后,喷施吡虫啉(1 mg·L-1),分别在0 、1/6 、1/2、1 、2 、4 、8 h取样50头,3次生物学重复,液氮速冻后提取RNA。
1.4.3 定量分析 根据烟粉虱β亚基基因的cDNA序列设计荧光定量引物NPf2/NPr2(表1),以actin作为内部参照(Af1/Ar1)。内参基因actin的PCR扩增条件与β亚基相同,除了退火温度调整至62 ℃。3次重复。以非处理组相对应基因的表达量为对照,按照2-△△Ct方法分析目标基因的相对表达量及其标准差。
1.5 烟粉虱NAC蛋白β亚基的原核表达
1.5.1 原核表达载体构建 用含有EcoR I和SalI酶切位点的引物NPf3/NPr3(表1)扩增NAC蛋白β亚基基因,PCR产物纯化回收连pGEM-T载体测序验证,同时用EcoRI和SalI酶切pGEM-T-βNAC与pMAL-c2x载体,分别回收目的片段和pMAL-c2x载体骨架用T4 DNA Ligase连接并转化至Trans1-T1感受态细胞,测序鉴定后将pMALc2x-BtβNAC成功重组质粒转化至表达感受态E.coliBL21。
1.5.2 pMALc2x-BtβNAC的表达及聚丙烯酰胺凝胶分析 挑取含pMALc2x-BtβNAC质粒的单克隆于5 mL LB液体培养基中(含50 mg·L-1Ampicillin),37 ℃培养12 h后,在无菌工作台中用移液器吸取过夜培养菌液500 μL,加入到50 mL含Ampicillin(50 mg·L-1)的新鲜LB液体培养基中继续振荡培养,当菌液OD600值达到0.5时添加异丙基-β-D-硫代吡喃半乳糖苷(IPTG终浓度0.3 mmol·L-1)进行诱导,收集诱导不同时期(0、0.5、1、3 h)菌液2 mL并提取诱导菌体总蛋白,用15%的SDS-聚丙烯酰胺凝胶电泳检测。
2 结果与分析
2.1 烟粉虱NAC蛋白β亚基基因克隆及序列分析
从烟粉虱成虫来源mRNA克隆获得1个与预期相符的特异性条带(图1),经测序结果分析确认该序列为目的片段,命名为BtβNAC。具体序列分析结果显示,BtβNAC基因全长856 bp,其中5’非翻译区长度为134 bp,3’非翻译区长度为272 bp,开放阅读框编码氨基酸149个,相对分子质量约为16.16 kD,等电点约为8.97,属于碱性蛋白。从烟粉虱基因组DNA中扩增获得BtβNAC基因的基因组片段(图1),测序结果证实该片段长度为4 430 bp,经与cDNA序列比对分析显示BtβNAC基因包含2外显子和1内显子(图2),该序列已提交到GenBank数据库,登录号MK229180。
2.2 烟粉虱NAC蛋白β亚基结构分析
利用生物信息学软件(http://www.expasy.org/cgi-bin/prosite/)在线对推导的氨基酸序列进行分析结果显示,该蛋白含有典型的NAC功能域(A32~L97),表明该蛋白隶属于NAC蛋白家族,C端缺少α亚基特有的UBA功能域,在N端含有β亚基特有的RRK-(X)2-KK基序(图 2),因此判定克隆所得基因属于NAC蛋白β亚基。此外,该蛋白还存在3个蛋白激酶C磷酸化位点(3~5,20~22,31~34)和1个酪氨酸激酶磷酸化位点(31~34)。
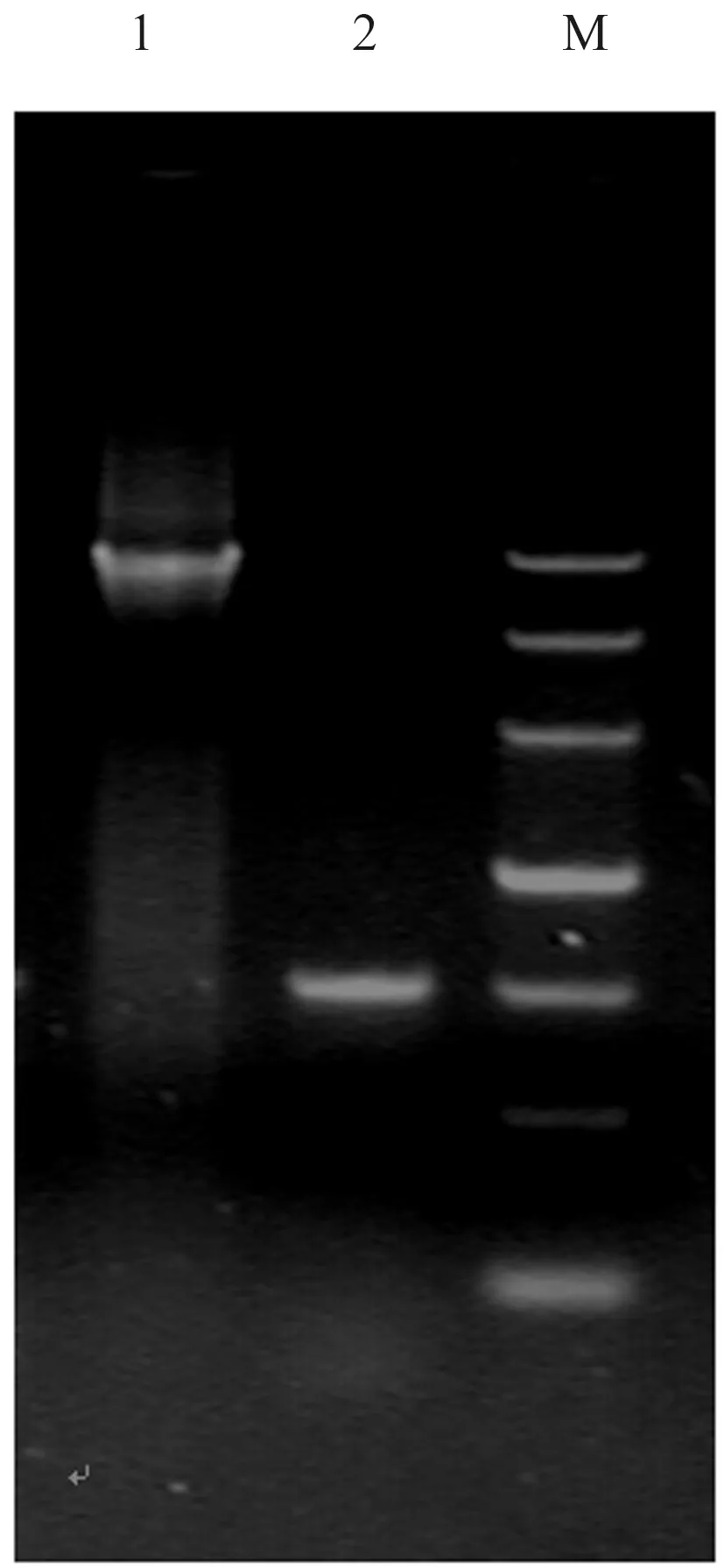
M:DL5000;1:烟粉虱BtβNAC gDNA;2:烟粉虱BtβNAC cDNA M:DL5000;1:cDNA of BtβNAC from Bemisia tabaci;2:Genomic DNA of of BtβNAC from Bemisia tabaci图1 烟粉虱BtβNAC的cDNA、基因组DNA扩增Fig.1 The result of amplification of cDNA and genomic DNA of BtβNAC from Bemisia tabaci


图2 烟粉虱BtβNAC基因序列及其推导的氨基酸序列Fig.2 Nucleotide sequence of the BtβNAC gene from Bemisia tabaci and its deduced amino acid sequence
NAC功能域聚类分析显示(图3),烟粉虱NAC蛋白β亚基与其他已知β亚基的氨基酸相似性介于28.9%~89.9%之间,与蚂蚁Camponotusfloridanus(EFN63323),印度跳蚁Harpegnathossaltator(EFN88288),散白蚁Reticulitermesflavipes(AAP33157)和红火蚁SolenopsisinvictaBuren(EFZ09605)相似性最高(89.9%),与刺舌蝇Glossinamorsitansmorsitans(ADD19794)、大西洋鲑Salmosalar(ACI69109)、人Homosapiens(NP_001032726)、家鼠Housemouse(NP_663430)和非洲爪蟾Xenopuslaevis(NP_001088356)的相似性居中,分别为84.4%、80%、71.1%、71.1%和71.1%,而与酿酒酵母的相似性最低,分别为33.3%(EGD1,X78725)和28.9%(BTT1,X78724)。

图3 已知部分NAC蛋白NAC domain分析黑色方框表示有100%的相似性,深灰色表示有50%的保守氨基酸Fig.3 Comparison of NAC domain sequences and dendrogram of ERF proteins Boxes in black represent 100% similarity,while dark grey indicates 50% conserved amino acids
2.3 烟粉虱β亚基基因的转录表达分析
NAC蛋白β亚基的转录表达对烟粉虱的生长发育关联密切。从图4可知,β亚基在烟粉虱MED隐种的各个生长阶段均可检测到表达,且不同生长阶段表达量存在显著差异。例如,与样品卵中β亚基的表达量相比较,1龄、2龄和3龄若虫,以及伪蛹和成虫期体内的表达量分别为卵的118.14、8.78、17.78、4.99、66.21倍,分析结果表明,β亚基基因的转录表达与烟粉虱生长发育密切相关。
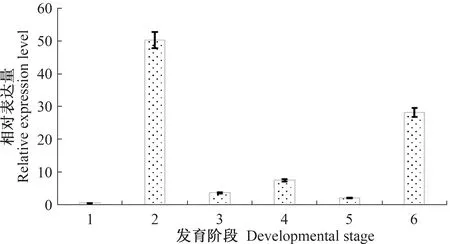
以卵中表达量作为1,选取actin作为标准化内参,用2-△△Ct法计算相对表达量。图中数据为平均值±标准误。1:卵;2:1龄若虫;3:2龄若虫;4:3龄若虫;5:伪蛹;6:成虫。The relative expression level of BtβNAC in different developmental stages of Bemisia tabaci MED is determined by 2-△△Ct method compared with that in the eggs,and the actin is selected as the housekeeping gene.Data in the figue are mean±SE. 1:eggs;2:1st instar;3:2nd instar;4:3rd instar;5:Pupa;6:adult.图4 烟粉虱BtβNAC不同发育时期相对表达量Fig.4 Relative expression level of BtβNAC in different developmental stages of Bemisia tabaci
2.4 烟粉虱β亚基的表达受吡虫啉胁迫诱导
在喷施1 mg·L-1吡虫啉胁迫条件下,烟粉虱MED隐种成虫体内NAC蛋白β亚基的表达量总体受到抑制,并呈现多峰曲线变化趋势(图5)。处理10 min时β亚基的表达量迅速降至对照的49.67%,处理30 min时该基因的表达量短暂回升至对照的92.63%,但随着胁迫时间的延长该基因表达量又迅速下降,如处理1 h和2 h时的表达量分别为对照的35.32%和52.39%,在胁迫4 h表达量回升至峰值的110.01%,此后表达量迅速回落,处理6 h和8 h时的表达量分别为对照的34.88%和67.37%。
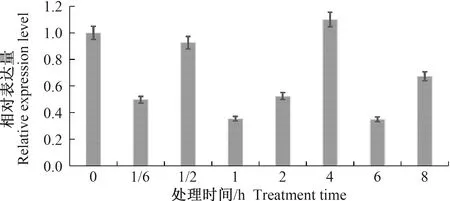
图5 吡虫啉处理条件下烟粉虱成虫体内β-NAC的相对表达量Fig.5 Relative expression level of β-NAC of Bemisia tabaci adult under imidacloprid stress
2.5 β亚基融合蛋白体外表达
在构建完成测序证实重组表达载体pMALc2x-BtβNAC正确之后,利用大肠杆菌菌株BL21对pMALc2x::BtβNAC融合蛋白进行体外表达,分别提取各样品总蛋白,采用SDS-PAGE蛋白质电泳检测。检测结果显示(图6),在异丙基-β-D-硫代吡喃半乳糖苷(0.3 mmol·L-1)诱导下,重组载体pMALc2x-BtβNAC菌体总蛋白样品中可检测到分子量约为58.8 kD的特异性蛋白条带,与预期pMALc2x::BtβNAC融合蛋白分子量大小相符;该结果证实异丙基-β-D-硫代吡喃半乳糖苷(IPTG)可强烈诱导pMALc2x::BtβNAC融合蛋白的表达,如诱导0.5 h时表达量即达到峰值(泳道3),延长诱导时间并不能增加表达量,例如诱导1 h(泳道4),3 h(泳道5)后的表达量未见增加,反而略有下降趋势;融合蛋白pMALc2x::BtβNAC同时存在于上清液和沉淀中,且沉淀中的含量略高于上清液。
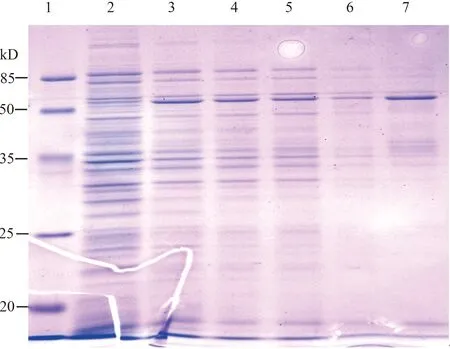
1:蛋白质相对分子质量标准物;2 to 5:分别为IPTG诱导0、0.5、1、3 h 样品总蛋白;6:超声波破碎后上清液总蛋白;7:超声波破碎后沉淀总蛋白。1:Protein molecular weight marker;2 to 5:Ttotal protein from cells induced by IPTG for 0,0.5,1,3 h,respectively;6:Ttotal protein from the supernatant;7:Ttotal protein from the pellet.图6 烟粉虱β-NAC体外表达SDS-PAGE电泳检测Fig.6 Expression of β-NAC of Bemisia tabaci in vitro detected by SDS-PAGE
3 讨论
存在于在真核生物的异源二聚体型NAC蛋白均由α和β亚基构成[28],虽然两者间氨基酸序列相似性只有26%左右,但是均含有高度保守的NAC功能域[29],本研究的烟粉虱BtβNAC同样也具有1个由66个氨基酸构成的NAC功能域,并且与其他动物来源的β亚基的NAC功能域相似性介于71.1%~89.9%,然而与酿酒酵母的EGD1和BTT1相似性只有33.3%和28.9%,表明NAC功能域的分化处于物种进化的早期阶段。与其他物种来源β亚基缺少UBA功能域类似,BtβNAC同样不含该功能域,而是在N端含有RRK-(X)2-KK基序,表明β亚基在物种间具有高度的一致性。
对于生物体来说及时表达正确折叠和定位的功能蛋白质至关重要[30],生物体细胞内已开发出多种启动、调节和监控重要蛋白合成及成熟的调控机制,包括蛋白质的N端加工、膜定位,以及新生肽的折叠等[31]。已有研究表明,NAC蛋白基因在生物体内的不同组织中均能表达,但表达水平的差异与发育程度密切相关[32],这可能是由于NAC通过与合成至核糖体的新生肽直接结合参与新生肽的早期折叠有关。本研究结果同样证实BtβNAC在烟粉虱的各发育阶段均表达,尤其在1龄若虫和成虫阶段表达量较高,显著高于卵、2龄、3龄和伪蛹。吡虫啉作为一种内吸性杀虫剂,通过竞争性结合烟酸乙酰胆碱酯酶受体干扰昆虫体内运动神经系统的化学信号传递,使其麻痹死亡。本研究结果显示,在喷施低浓度吡虫啉条件下,烟粉虱成虫内体BtβNAC基因的转录表达呈抑制下调模式,这可能是在吡虫啉处理条件下烟粉虱体内的新陈代谢下降,新生蛋白合成表达量降低,对于主要作为新生肽分子伴侣功能的BtβNAC需求量降低所致。本研究过程构建的原核表达载体pMALc2x-BtβNAC,其融合蛋白pMALc2x::BtβNAC在宿主菌BL21中高效稳定表达,其表达产物同时以可溶性蛋白和沉淀形式存在,这为后续纯化BtβNAC蛋白,制备抗体等奠定了基础。
猜你喜欢
上海蔬菜(2022年6期)2022-12-31
麦类作物学报(2022年3期)2022-05-19
临床与实验病理学杂志(2022年3期)2022-04-06
河北果树(2021年4期)2021-12-02
环境昆虫学报(2021年5期)2021-10-19
四川蚕业(2021年4期)2021-03-08
核农学报(2021年2期)2021-01-28
世界最新医学信息文摘(2020年68期)2020-12-25
烟台大学学报(自然科学与工程版)(2020年1期)2020-02-08
植物保护(2018年5期)2018-12-05
