
青藏高原多年冻土区不同水分条件的高寒草甸根系功能性状对增温的响应
2020-11-12 07:29王根绪唐川川黄克威赵小祥杨晓明
生态学报 2020年18期
杨 凯,王根绪,张 涛,唐川川,黄克威,赵小祥,杨晓明,杨 燕,*
1 中国科学院、水利部成都山地灾害与环境研究所,成都 610041 2 贵州大学新农村发展研究院,贵阳 550025 3 中铁西北科学研究院有限公司,兰州 730000 4 中国科学院大学,北京 100049
工业革命以来,化石燃料燃烧和森林砍伐等人类活动导致全球气温增加了0.87℃,模型预估2030—2052年将增加1.5℃[1]。在过去的50年里,青藏高原的平均地表温度以每10年0.35℃的速率增加了1.8℃,增温速率高于全球平均水平,是全球气候变化的敏感区[2]。由于高海拔和冻土的存在,位于青藏高原多年冻土区的高寒生态系统尤为脆弱[3],因此研究全球气候变化对青藏高原多年冻土区生态系统的影响较其他陆地生态系统更具有独特性与超前性,使得青藏高原成为研究陆地高寒生态系统对气候变化响应模式及适应机制的天然实验室。
气候变暖导致气温和土壤温度升高[4],引起冻土退化并造成土壤水热条件季节性变化[5],将直接或者间接地影响植物的生理代谢过程[6-7]、改变植物体内元素含量分配[8]以及植物资源获取策略[9]和生物量分配格局[10],进而影响生态系统的群落结构和物种多样性。北美洛基山高寒苔原的模拟试验结果显示[11],增温与补充土壤水分同时处理时,植物群落生产力增加,单独增温降低高寒苔原的地上生产力,表明土壤水分可以改变苔原植被生产力对增温的响应模式。高寒地区土壤水分会限制植物生长模式对增温的响应,气候变暖在不改变植物生产量的情况下,促进了植物早期物候和生长速率,但在秋季由于水分胁迫加剧导致生长提前结束[12]。同时,土壤水分作为影响高寒草地多样性和生产力的主要驱动因子[13],可以调节植物功能群多度和叶片功能性状对增温的响应[14]。覆盖北极苔原的植物叶片功能性状对增温的响应的结果表明,在不受土壤水分限制的条件下,长期增温将引起植物叶片功能性状向资源获取型转变,即高比叶面积和低叶片干物质量[15]。这些结果强调了植物地上功能性状对温度响应过程中存在对土壤水分的权衡,然而却少有研究探究地下根系功能性状是否且如何响应温度和水分的变化。
草地生态系统是全球碳平衡的重要组成部分[16],根系生物量占草地植物总生物量的一半以上,在碳循环和养分循环中起着重要作用[17-18]。根系碳含量是植物碳元素分配的结果,受温度、土壤含水量、养分条件等环境因素的影响[19-20]。暖干化胁迫导致青藏高原高寒草甸将更多碳化合物向根系分配[21],而不同草甸类型根系碳含量对增温的响应不一致,增温增加矮嵩草草甸根系碳含量,而降低了金露梅灌丛草甸根系碳含量[22]。植物根系的碳分配对增温的差异性响应,与可获取水分密切相关,进一步强调了植物碳分配对增温响应过程中水分的重要性[23]。涵盖不同气候区的根系化学计量学特征的综合分析发现,高寒植物根系氮含量低,是由于低温限制了植物对养分的吸收利用[24]。增温满足了植物对低温的要求[25],增加根系碳含量,从而提高根系的C∶N比值。同时,暖干化气候会降低多年冻土区莎草科植物根系的氮含量[26],提高藏嵩草地下部分C∶N比[27]。前人研究多侧重于增温对植物C、N、P等养分含量的影响,而结合根系化学计量学和根系形态特征,从植物地下资源利用策略的角度来探究植物根系对增温的响应机制研究相对较少,尤其是在青藏高原多年冻土区,对不同土壤水分含量的高寒草甸根系功能性状的研究。
青藏高原草地生态系统多分布在海拔3000 m以上,其中分布面积约为0.48×106km2的高寒草甸和0.049×106km2的高寒沼泽草甸大多位于多年冻土区域,是青藏高原分布广泛的两种草地生态系统[28-29]。分别以小嵩草(Kobresiapygmea)和藏嵩草(Kobresiatibetica)为优势种的高寒草甸和沼泽化草甸所含热值高、营养丰富,是青藏高原重要的草地资源[30]。本研究以风火山地区生长季(5—9月)土壤水分差异24%的小嵩草草甸和藏嵩草沼泽化草甸对研究对象,对比分析两种草甸根系功能性状对模拟增温的响应,试图从根系功能性状所表征的植物资源利用策略的角度,初步探究土壤水分差异是否可能影响两种草甸植物对增温的响应模式和适应策略,解决如下科学问题:不同土壤水分含量的草甸地下生物量和根系功能性状对增温的响应模式,以及基于根系功能性状所指示的两种草甸对增温的适应策略。
1 材料与方法
1.1 研究区域概况
研究地点位于青藏高原腹地长江源多年冻土和高寒草甸均较典型的风火山地区(92°50′—93°30′E,34°40′—34°48′N),气候属青藏高原干旱半干旱气候区,年均温-5.3℃,年降雨量270 mm,降水主要发生在植物生长季(5—9月),年蒸发量1478 mm,冻结期为每年的9月至次年4月[29]。主要植被类型为高寒草甸和高寒沼泽草甸,其中高寒草甸主要分布于山地的阳坡、阴坡、圆顶山、滩地和河谷阶地,分布上限可达5200 m左右;高寒沼泽主要分布在海拔3200—4800 m的河畔、湖滨、排水不畅的平缓滩地[31]。
1.2 试验设计
2012年8月,在研究区域选择植被分布相对均匀一致的小嵩草高寒草甸和藏嵩草沼泽化草甸作为研究对象,分别建立100m×100 m样地。采用开顶增温室(Open top chambers, OTCs)模拟环境温度增加。增温室采用透明有机玻璃(PLEXIGLAS, Evonik Degussa, Essen, Germany, made in Shanghai, China)加工成正六边形圆台状,底面积约为1m×1m,高40 cm,圆台底边长106 cm,开口处边长60 cm,斜边与地面呈60°夹角[32]。在小嵩草草甸和藏嵩草沼泽化草甸样地中,各随机布设6个试验小区,试验小区间隔距离为5—8 m。在小区内随机布设对照样方和OTC增温样方各一个,对照和增温样方相隔2—3 m。同时,通过在温室和对照样方内布设传感器和数据采集器(MG-EM50, Decagon, Pullman, USA),以测定温室和对照样方地上20 cm空气温度(℃)和地下5 cm土壤相对含水量(体积分数,%)及土壤温度(℃),测定间隔为每30 min一次。每个样地设置6个小区,每个小区分别包括1个1 m×1 m的对照样方和OTC增温样方。每个样方再划分为4个50 cm×50 cm的副样方,随机选取1个副样方来布设内生长袋。2015年9月中旬生长季节结束,用直径5 cm 的土钻,在随机选取的副样方内随机钻取3个深度20 cm的孔,将内生长袋(网眼2 mm的尼龙网缝制成直径5 cm,长度为24 cm的圆柱状袋子)垂直放入孔中。按照无根土土壤的含水量及土壤容重称取所需土壤质量垂直填入生长袋内,生长袋口露出地面约4 cm,插红色标签以标记位置和编号。因此,每个样方布设了3个重复的内生长袋,对照和增温共计36个内生长袋。
1.3 样品采集与指标测定
2016年9月中旬,搜集上一年埋设的36个内生长袋,取样后迅速编号放入4℃冷藏箱带回实验室分析。在实验室将生长袋沿袋壁竖直剪开,用尺量取上下各10 cm两层,在分界处切开后分别用60目标准土壤筛(0.28 mm孔径)将根系与土壤分离,得到0—10 cm和10—20 cm两土层根系样品。用水冲净后根据颜色、柔韧性和是否有根毛附着辨别活根。随机分别选出两层根系中一部分样品,使用数字化扫描仪(Epson scanner)进行扫描,并用专业根系扫描分析系统WinRHIZO(WinRhizo Pro STD 1600, Regent Inc.Canada)对根系图像分析获得根长、根表面积和根体积等形态指标,扫描后的根放置在65℃鼓风烘箱中烘干至恒重,用以计算比根长(Specific Root Length, SRL)、比根面积(Specific Root Area, SRA)、根组织密度(Root Tissue Density, RTD)。未用于扫描的根系经同样方法烘干获取干重后与扫描根系烘干样品合并,使用球磨仪(Retsch, MM400)磨碎后并通过80目筛,经元素分析仪(Elementar, VarioMacrocube, Germany)测定植物根系的C、N含量,测定结果用单位质量的养分含量表示[33]。通过根扫描形态指标与扫描部分干重计算得到以下指标[34]:
SRL=Rlength/Rmass;
SRA=Rarea/Rmass;
RTD=Rmass/Rvolume
式中,SRL(cm/g)为比根长,SRA(cm2/g)为比根面积,RTD(g/cm3)为根组织密度,Rlength(cm)、Rarea(cm2)、Rvolume(cm3)和Rmass(g)分别为扫描图像根系总长度、扫描图像根系总面积、扫描图像根体积和扫描根干重。
1.4 数据分析
所有数据经Excel记录整理,用R(3.4.3, http://www.R-project.org/)完成数据的统计分析和作图。其中,采用单因素方差分析增温和植被类型对根系生物量、比根长(SRL)、比根面积(SRA)、根组织密度(RTD)、根系C、N含量及根系C:N比值的影响,采用双因素方差分析探索植被类型与增温处理对根系功能性状的交互效应,所有统计结果均以P<0.05为显著差异标准。
2 结果
2.1 两种草甸气温和土壤水分的差异
自然条件下,小嵩草草甸和藏嵩草沼泽化草甸的地上20 cm处气温和地下5 cm处土壤水分存在差异。4年(2013—2016年)的气象数据显示,藏嵩草沼泽化草甸年均气温较小嵩草草甸低-0.31℃,年均土壤水分高出小嵩草草甸2.11%(体积分数)。在植物生长发育尤为重要的生长季(5—9月),藏嵩草沼泽化草甸土壤水分高出小嵩草草甸7.43%(体积分数),气温高出0.34℃(图1)。可见,生长季期间,两种草甸气温的差异为8.72%,而土壤水分的差异达到23.9%。
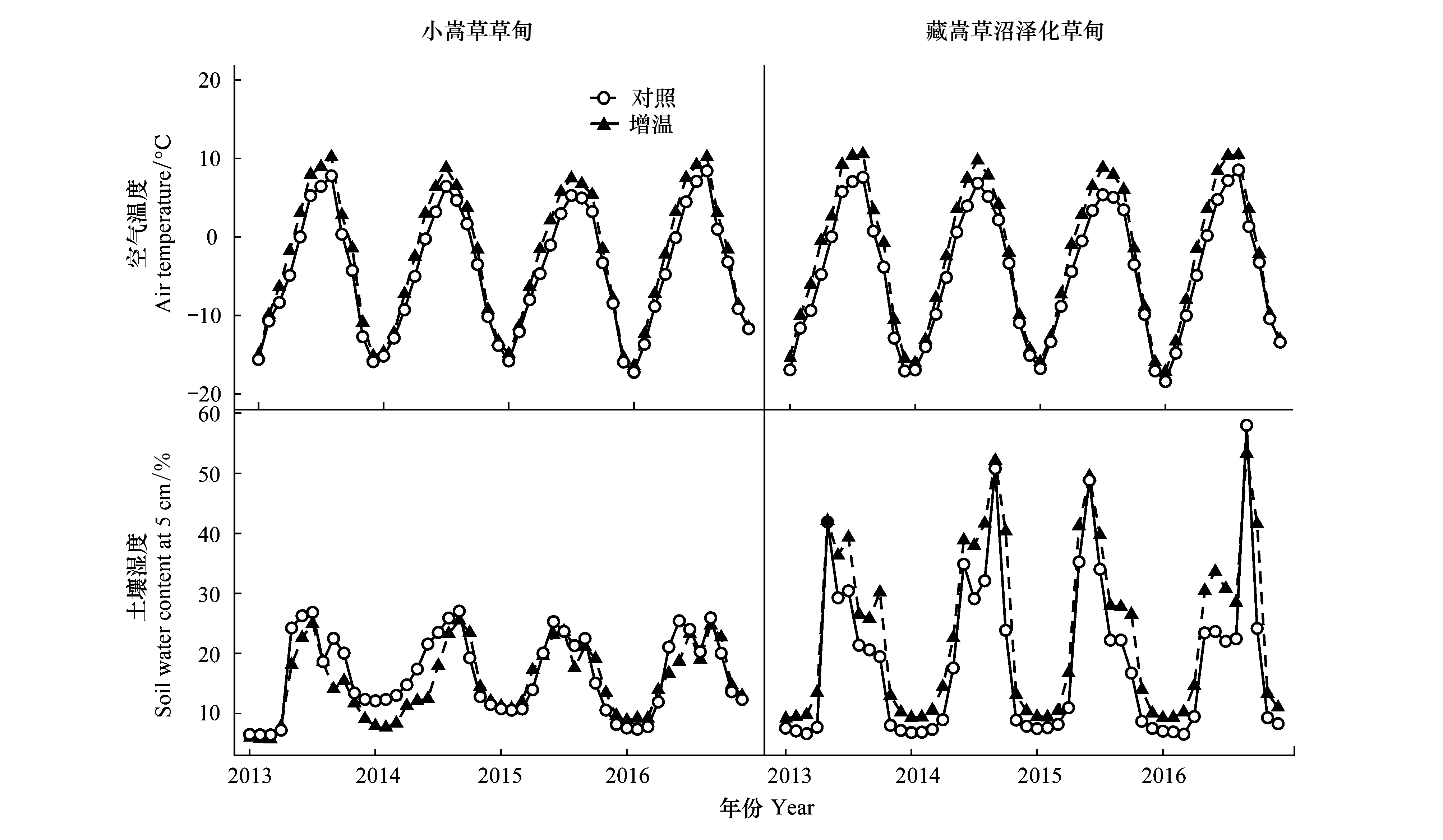
图1 两种草甸月平均气温和月平均土壤水分
2.2 模拟增温对两种草甸气温和土壤水分的影响
2013—2016年的气象数据显示,模拟增温处理提高了小嵩草草甸和藏嵩草沼泽化草甸的年均气温,增幅分别为1.85℃和2.22℃(图1)。同时,与对照样方相比,OTCs中小嵩草草甸年均土壤湿度降低1.31%(体积分数),而藏嵩草沼泽化草甸年均土壤湿度增加5.03%(体积分数)。生长季期间,较对照相比,模拟增温处理后小嵩草草甸和藏嵩草沼泽化草甸气温分别升高了2.5℃、2.93℃;同时,降低了小嵩草草甸土壤水分3.71%(体积分数),增加了藏嵩草沼泽化草甸土壤水分5.31%(体积分数)。可见,由于开顶式温室的增温作用,两种草甸全年和生长季的空气温度升高;土壤水分对增温的响应存在差异,增温降低了小嵩草草甸土壤水分,却增加了土壤含水量较高的沼泽化草甸土壤水分,在生长季差异更为明显。
2.3 增温对两种草甸根系生物量的影响
自然条件下0—10 cm和0—20 cm土壤层中,藏嵩草沼泽化草甸根系生物量分别高出小嵩草草甸46.6%、34.7%,差异达到显著水平(P<0.05)。与对照相比,增温处理后小嵩草草甸根系生物量在0—10 cm和0—20 cm分别增加31.0%和28.3%(P<0.05),说明增温导致小嵩草草甸生物量转移向根系,主要体现在0—10 cm浅层土壤中(图2),而对藏嵩草草甸根系生物量无影响。

图2 增温对两种草甸在不同土壤层中根系生物量的影响
2.4 增温对两种草甸根系形态的影响
自然条件下0—10 cm土壤层中,藏嵩草沼泽化草甸比根长与比根面积均显著低于小嵩草草甸(P<0.05),在增温后差异变得不显著;两种草甸根组织密度无差异,增温后,藏嵩草沼泽化草甸显著(P<0.05)低于小嵩草草甸(图3)。10—20 cm土壤层中,两种草甸比根长与比根面积无差异,增温后,藏嵩草沼泽化草甸显著高于小嵩草草甸;藏嵩草草甸根组织密度在自然状况和增温后均显著(P<0.05)低于小嵩草草甸。
模拟增温后,较对照相比(图3),10—20 cm土壤层,藏嵩草沼泽化草甸比根长和比根面积分别显著(P<0.05)增加86.43%和89.78%;小嵩草草甸根组织密度在0—10 cm、10—20 cm分别显著(P<0.05)提高了19.66%和23.43%。可见,增温处理对藏嵩草根系形态的影响主要体现在提高比根长和比根面积,促进对土壤水分和养分的吸收;对小嵩草草甸的影响主要体现在增加根组织密度,提高其抗胁迫能力与根系寿命。
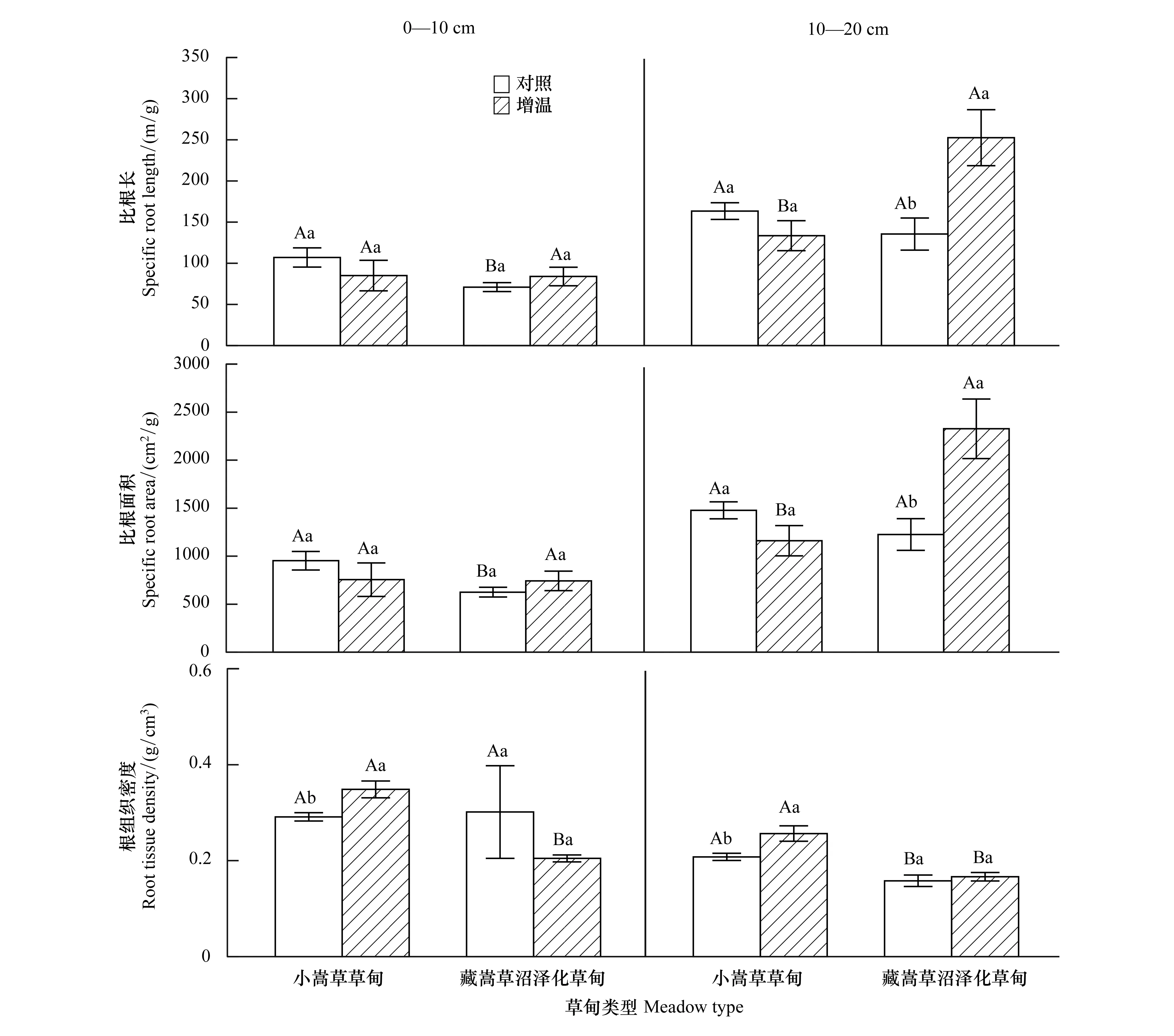
图3 增温对两种草甸根系形态特征的影响
2.5 增温对两种草甸根系C、N含量及化学计量比的影响
自然条件下0—10 cm和10—20 cm土壤层中,藏嵩草沼泽化草甸根系C、N含量均显著低于小嵩草草甸(P<0.05),而根系C∶N比值在各土层均无差异,增温后,藏嵩草沼泽化草甸根系C含量在各土壤层均显著高于小嵩草草甸(P<0.05),根系N含量和C∶N比值无差异(图4)。
模拟增温后,较对照相比,小嵩草草甸根系C含量在各土壤层均显著降低(P<0.05),而藏嵩草沼泽化草甸根系C含量在各土壤层均显著增加(图4)。增温处理增加了藏嵩草沼泽化草甸在10—20 cm根系N含量(P<0.05),却降低小嵩草草甸在10—20 cm根系N含量。增温显著提高了两种草甸在10—20 cm根系C∶N比值。可见,两种草甸根系C、N含量对温的响应不一致,但均提高了两种草甸对N素的利用。
2.6 增温与植被类型的交互效应分析
增温处理与植被类型的交互效应显著时,说明两种草甸对增温的响应存在显著差异,结合单因素分析结果可对交互效应给出进一步解释,即两种草甸对增温的差异性响应是否会改变植被类型间原有差异。在0—10 cm和10—20 cm土壤层中,两种草甸根系碳含量存在极显著(P<0.01)的交互效应(表1),结合图4可以看出,自然条件下藏嵩草沼泽化草甸根系C含量显著低于小嵩草(P<0.05),由于两种草甸根系C含量对增温的响应相反,即显著提高了藏嵩草沼泽化草甸根系C含量并显著降低了小嵩草根系C含量(P<0.05),使得增温处理后藏嵩草沼泽化草甸根系C含量在各土壤层均显著高于小嵩草。在10—20 cm土壤层中,比根长与比根面积存在极显著(P<0.01)交互效应(表1),结合图3可见,自然条件下藏嵩草沼泽化草甸比根长和比根面积均低于小嵩草草甸,由于二者对增温的响应不一致,即增温显著提高了沼泽化草甸比根长和比根面积而对小嵩草无影响,导致了增温后沼泽化草甸比根长和比根面积显著高于小嵩草草甸。同样地,10—20 cm土层中,自然条件下两种草甸根系N含量无差异,增温处理显著提高了沼泽化草甸根系N含量而对小嵩草草甸无影响(图4),导致沼泽化草甸根系N含量在增温后显著高于小嵩草草甸。可见,增温处理对根系C、N含量、比根长及比根面积的影响强于植被类型,两种草甸对增温不一致的响应改变自然条件下原有的差异。
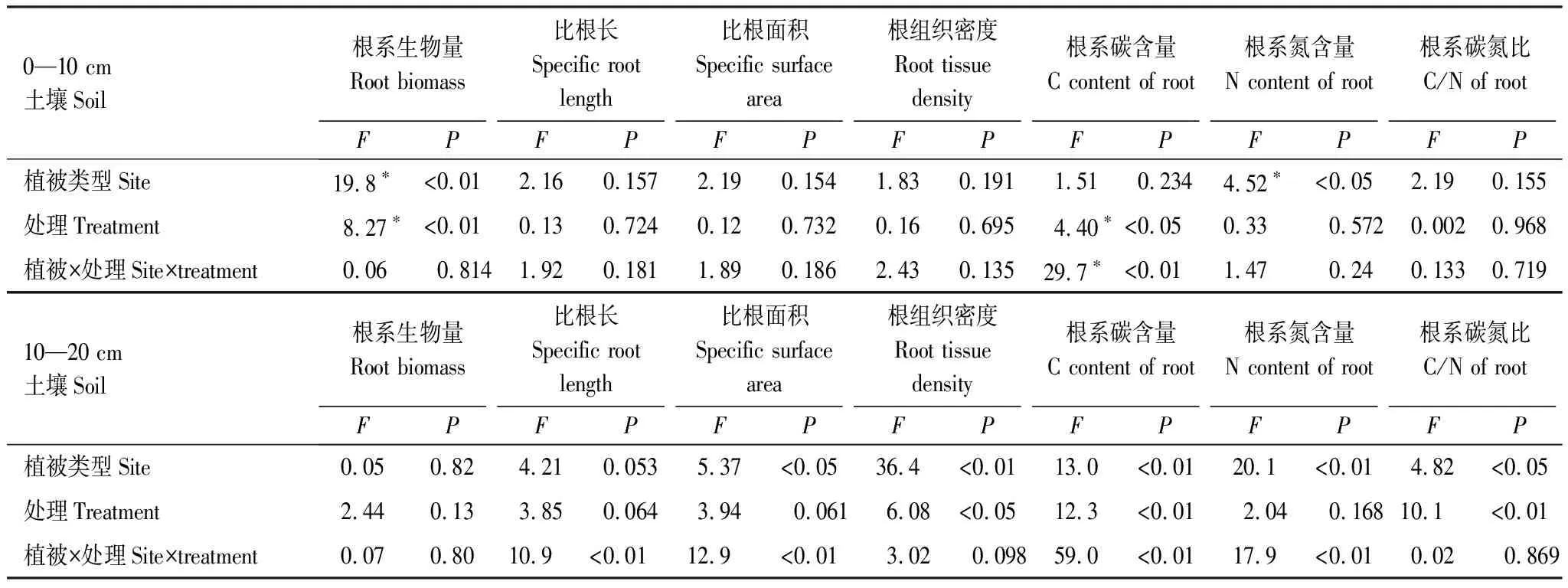
表1 基于一般线性模型的双因素方差分析

图4 增温对两种草甸根系碳、氮含量及碳氮比的影响
3 讨论
3.1 增温对两种草甸根系生物量的影响
本研究中,增温处理增加了小嵩草草甸0—20 cm土壤根系生物量,并主要体现在0—10 cm浅层土壤中。该研究结论与在海北站[35]和风火山[36]高寒草甸的模拟增温试验结果一致,即增温引起生物量向地下转移,并集中于表层。Loik[37]认为增温满足了高寒地区受低温限制的植物对温度的需求,促进光合作用从而促进植物地上、地下部分的生长。而Makiranta等[38]的研究表明,增温主要通过改变地下土壤水分来影响植物的物候节律和生物量生产,水分含量会影响植物对增温的响应。由于试验点位于青藏高原多年冻土区,植物受低温胁迫,植物根系的地下分布特征与土壤水热格局均受活动层冻融作用的影响[39]。岳广阳等[40]研究了多年冻土区植物根系对活动层温度变化的响应,认为植物生长所需水分一部分来自于活动层季节性融化过程,温度调控冻土冻融作用,改变土壤水热过程,影响植物根系生长发育与垂直分布格局。模拟增温引起小嵩草草甸向暖干化气候发展,减少浅层土壤水分,水分的降低伴随着可利用养分的减少,导致浅层土壤养分水分胁迫。胁迫环境下,小嵩草草甸生物量分配策略发生改变,将更多的生物量转移向地下用于吸收水分和养分,使得0—10 cm和0—20 cm根系生物量的显著增加。藏嵩草沼泽化草甸土壤水分的增加可能因为增温使得冻土活动层厚度增加和冻结水的融化,引起水分蒸散发加剧补给了浅层土壤中水分[41],同时,藏嵩草沼泽化草甸较为发达的浅层根系形成的草皮层具有较强的持水保水作用[42]。增温、增水条件下的沼泽化草甸将更多的生物量用于发育地上部分,以获得对光照等资源更强的竞争力,因此其根系生物量无显著变化。从土壤水分、养分角度与植物生物量分配策略角度可解释两种草甸根系生物量对增温的响应差异。
3.2 增温对两种高寒草甸根系功能性状的影响
根系的功能性状特征与其资源获取策略紧密相关,并对资源条件与气候的变化在时空上做出响应[43],比根长、比根面积、组织密度以及根系碳、氮含量等被认为是植物适应环境的有效性状[44]。一般认为[45]资源获取型植物具有高比根长、比根面积和根系氮含量等特征,从而实现对水分和养分的快速获取,资源保守型植物则对应低比根长与高根组织密度,以低代谢活动强度和长根系寿命以持续获取资源。本研究中,在增温处理后,显著增加了藏嵩草沼泽化草甸10—20 cm的比根长和比表面积,反映出沼泽草甸植物加强对10—20 cm土壤层资源的获取,这是由于增温引起沼泽化草甸10—20 cm土壤中水分和氮素等养分增加的结果[31],沼泽化草甸植物根系变长、表面积加大以提高对水分和养分的吸收。暖干化小气候加剧了小嵩草草甸土壤的贫瘠[31],小嵩草草甸提高0—10 cm和10—20 cm根组织密度以延长根系的寿命,实现对资源的持续利用,以维持生存[46]。两种草甸根系功能性状对增温的响应不同,反映了植物资源获取策略存在对环境因子的权衡,本研究中环境因子主要为土壤水分的差异。
增温一方面会改变土壤中养分元素的含量与分配格局,另一方面改变植物根系形态特征与资源获取策略[9],影响根系对土壤养分吸收与利用。在本研究中,两种草甸根系C、N含量对增温的响应不一致,即小嵩草草甸各土壤层根系C含量均显著降低,藏嵩草沼泽化草甸根系C含量及10—20 cm根系N含量均显著提高。增温引起两种草甸土壤水分改变的方向不同,导致小嵩草草甸与沼泽化草甸有各自特异的土壤水热过程[39]和养分格局[31];同时,增温后两种草甸的资源获取策略存在差异,这可能是导致两种草甸植物根系C、N含量差异主要原因。小嵩草草甸植物根系C含量在0—10 cm与10—20 cm均随温度升高而显著降低,究其原因,可能包括以下三个方面:一是由于增温导致土壤的暖干化,土壤养分的减少,增加了植物根系死亡率[25]。二是增温引起的土壤干旱加剧,影响了碳水化合物在韧皮部中的运输[47],使得植物向根系的碳分配受影响[48]。三是增温提高了根系的呼吸速率,使得根系的生长和维持需要消耗更多的碳水化合物[41]。增温后,藏嵩草沼泽化草甸根系C含量在0—10 cm和10—20 cm土壤层中均显著增加,主要原因是在水分和养分较为富足的条件下,温度升高加强光合作用,促进光合产物生产[48],有更多的碳水化合物能分配到地下[49];沼泽化草甸根系N含量在10—20 cm显著增加是由于其比根长与比根面积的提高,促进对土壤养分的吸收。C∶ N的比值大小表示植物吸收单位养分元素所同化碳的了能力,可以在一定程度上反映植物对养分元素的吸收利用率[50]。在本研究中,温度升高显著提高了小嵩草草甸和藏嵩草沼泽化草甸10—20 cm根系C∶N比值,这表明了增温条件下,小嵩草草甸与藏嵩草沼泽化草甸均能更有效地利用氮素。考虑到土壤水热与养分条件的差异,两种草甸提高养分利用速率的原因存在不同。增温引起小嵩草草甸土壤干旱加剧,与对照相比,可利用氮在土壤中流动与扩散受阻[51],同时较为干旱的土壤增大了对根系伸长的机械损伤,小嵩草草甸植物提高氮素利用效率减缓土壤中N素限制。在土壤水分富足的藏嵩草沼泽化草甸中,增温提高了有机物降解和氮素矿化速率[51],无机态氮含量增加,养分水分充足,温度升高增强种间竞争,沼泽化草甸植物形成对资源快速获取与充分利用的策略以提高对土壤水分、养分的竞争力。
3.3 增温处理与植被类型的交互作用
不同生态系统或植被类型在生境条件,群落组成和结构,土壤化学性质等差异都可能是植物对增温的响应不一致的成因[52]。从整体上看,本研究中,小嵩草草甸和藏嵩草沼泽化草甸在根系形态特征和根系碳氮含量两方面对增温的响应存在不一致(表1)。根系形态特征与植物资源获取策略有密切关系[9],资源获取型植物具有高比根长、比根面积的高吸收功能形态[53],而在贫瘠土壤中生长缓慢的植物具有高组织密度的长寿命根系[33]。土壤水分差异的小嵩草草甸与藏嵩草沼泽化草甸,根系形态特征对增温的不一致响应。小嵩草草甸土壤水分较为缺乏,是限制高寒草甸植物地下部分生长大于的关键因子[31],增温增加了土壤水分的散失,养分的流动扩散也因此受限制,土壤中营养资源匮乏[25],根系组织密度提高,降低对根系的碳输入,采取缓慢生长,提高寿命的保守型资源获取策略。藏嵩草沼泽化草甸,土壤中根系密集,保水持水有机物质多,植被盖度大[42],增温促进冻土融化提供一定补给,土壤养分水分富足。沼泽化草甸植物根系伸长、表面积增大,快速吸收土壤资源,促进植物生长发育。不同的资源获取策略也导致两种草甸根系N含量对增温的响应差异,资源获取型沼泽化草甸能更多的从土壤中获得养分,从而根系N含量随增温显著提高,并显著高于资源保守型小嵩草草甸。增温与植被类型对两种草甸根系C含量交互作用极显著(P<0.01),结合单因素方差分析结果(图4),一方面表明,两种草甸根系C含量对增温的响应不一致,另一方面说明增温改变了二者根系C含量原有的差异。其主要原因可能是,增温引起两种草甸生物量分配策略的差异,水分养分较为富足的藏嵩草沼泽化草甸,生物量更多用于地上部分生长发育;暖干化胁迫小嵩草草甸将更多的生物量分配向地下,快速积累的根系生物量稀释了根系中C、N元素。
4 结论
温度升高在一定程度上促进了小嵩草草甸与藏嵩草沼泽化草甸根系生物量的积累,但两种草甸具有特异的水热条件、土壤养分供给,资源的差异导致两种草甸采取不同的资源获取策略和生长策略以适应环境变化,具体体现在根系形态与化学计量学等功能性状特征对增温的响应不一致。在未来气候变暖的情境下,小嵩草草甸植物会采取延长根系寿命、降低资源获取速率的资源保守型策略以适应区域气候暖干化发展;而藏嵩草沼泽化草甸则会提高对养分的吸收利用,加大对资源的获取,以维持植物地上、地下部分的快速生长。但仅仅从形态学与化学计量学角度还不能够充分揭示不同植被对温度的响应模式与差异,未来需要结合植物物候学和植物生理学,考虑土壤养分、冻土冻融、植物物候、植物生理代谢过程,进一步探究青藏高原多年冻土区高寒生态系统对气候变暖的响应机制,为高寒生态系统应对气候变化,采取适应性管理等方面提供科学依据。
致谢:感谢中铁西北院风火山观测站、中国科学院贡嘎山高山生态系统观测试验站提供的帮助。
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
农业工程学报(2022年5期)2022-06-22
海外文摘·文学版(2021年3期)2021-05-19
今日农业(2020年19期)2020-12-14
活力(2019年21期)2019-04-01
中国科技信息(2015年2期)2015-11-16
植物营养与肥料学报(2014年1期)2014-03-11
塔里木大学学报(2014年3期)2014-03-11
自然资源遥感(2014年2期)2014-02-27
