
马铃薯StCDPKs基因家族成员鉴定与表达分析
2020-11-17 02:18洪旭升李世贵杨江伟罗红玉张宁司怀军
甘肃农业大学学报 2020年5期
洪旭升,李世贵,杨江伟,罗红玉,张宁,司怀军,3
(1.甘肃农业大学生命科学技术学院,甘肃 兰州 730070;2.甘肃农业大学农学院,甘肃 兰州 730070;3.甘肃省干旱生境作物学省部共建国家重点实验室培育基地,甘肃 兰州 730070)
钙依赖性蛋白激酶(calcium-dependent protein kinase,CDPKs)是细胞钙调节信号级联反应的关键枢纽,参与植物感知非生物、生物胁迫刺激的应激信号和生长发育过程中的激素调节等[1].CDPKs以Ca2+标记编码的信息调控下游特定目标蛋白的磷酸化,从而将Ca2+感应与特定单个蛋白的响应功能结合在一起.这些单体酶具有丝氨酸/苏氨酸(S/T)蛋白激酶催化域(KD),其特征子域(I-XI)通过自抑制域或连接区(AD)与钙调蛋白样域融合(CLD).CDPKs已经在绿藻、被子植物,包括双子叶植物和单子叶植物中被鉴定出来[2-3].除了植物界,CDPKs只存在于顶复门寄生虫和纤毛虫中[4-5].
被子植物中,CDPKs家族由16~47个成员组成.通常,其成员在系统发育上被分为4个亚家族(I-IV),其第二和第三亚家族可分为2组(a和b)[6].CDPKs基因在原始被子植物中具有多样性,但KD、AD和CLD在进化中维持序列高度保守导致了许多CDPKs之间的功能相似性、序列冗余和重叠.CDPKs的激酶活性和表达模式依赖于其N端的可变域(NTV)、亚细胞定位、动力学参数、底物特异性、Ca2+浓度等,这表明不同基因可以感知不同的刺激并发挥特定的作用.来自I和II的CDPKs亚型在CLD中含有4个EF-hand基序,而来自组III或组IV的一些亚型仅含3个EF-hand基序[7].
研究表明CDPKs基因可响应逆境胁迫,参与信号转导及植物的免疫反应,在植物生长发育过程中发挥重要作用.在拟南芥抗盐碱胁迫中AtCDPK3基因具有重要作用[8-9],AtCDPK4基因与ABA通路中的转录因子相互作用调节植物的耐旱性[10],AtCDPK6/9/10/11/12/32基因均参与ABA信号通路中的干旱应答过程[11-16],而AtCDPK5基因参与植物免疫调节反应[17].CDPKs基因的功能在水稻中也被大量报道,如OsCDPK1基因能够提高水稻的抗旱性,调节赤霉素的合成和促进种子的萌发[18].OsCPK12基因与提高耐盐性、增强抗病能力、诱导活性氧生成和叶片衰老有关[19-20].OsCPK31基因参与淀粉的合成,并促进籽粒的早充[21].茄属植物番茄中,CDPKs基因在抗病方面有较多研究,如SlCDPK10和SlCDPK18基因具有抗白叶枯病菌、稻瘟病菌和丁香假单胞菌等作用[22].
由于灌溉方式、肥料使用不当以及工业污染,土壤盐碱度有所提高[23].高盐度通常是由于土壤溶液中Na+和Cl-的浓度高,导致高渗和高离子条件,从而阻碍植物从土壤吸收水和养分[24].大多数农作物都是糖类植物,而盐分胁迫会影响其生长和生产力[25].
Fantino等[26]、龚记熠等[27]和Gromadka等[28]分别在马铃薯中鉴定出26、21和23个StCDPKs基因家族成员;龚记熠等[27]的鉴定中一个基因不属于StCDPKs基因家族;一个StCDPKs基因在Fantino等[26]的鉴定中不存在;而Gromadka等[28]的鉴定结果与Fantino等[26]的鉴定相一致.本研究通过两类隐马尔科夫模型来鉴定马铃薯StCDPK基因家族成员,分析该家族成员的基因结构、Motif、保守结构域、进化和组织表达特异性,并利用qRT-PCR分析盐胁迫下StCDPKs基因的表达情况,以期为深入研究马铃薯StCDPKs基因在盐胁迫中的功能提供基础.
1 材料与方法
1.1 试验材料
本试验以马铃薯栽培品种‘大西洋’原种为试验材料,以盆栽的方式种植,其中营养土和蛭石比例为2∶1,利用自然光照并在正常控水的条件下生长,培养75 d后分别采取根、茎、叶、花和块茎,速冻保存于-80 ℃冰箱;盐胁迫处理使用200 mmol/L NaCl浇灌植株[29],分别在处理0、1、2、4、8和16 h时采取植株叶片,速冻后保存于-80 ℃冰箱.
1.2 马铃薯StCDPKs基因的筛选与鉴定
从Ensembl中(http://asia.ensembl.org/index.html)下载马铃薯全蛋白组序列,在Pfam(https://pfam.xfam.org/)中下载CDPKs基因2个主要结构域的隐马尔科夫模型文件(HMM文件):蛋白激酶(PF00069)和EF-hand结构域(PF13499),以及其他8个EF-hand的HMM文件.通过执行HMMER程序来搜索并提取所有拥有蛋白激酶和EF-hand结构域的蛋白序列[30].用在线分析软件InterPro(http://www.ebi.ac.uk/interpro/)对候选基因做进一步验证,用ExPASy(https://web.expasy.org/protscale/)分析StCDPKs蛋白的理化性质.
1.3 马铃薯StCDPKs基因结构分析及进化树构建
用在线工具GSDS 2.0(http://gsds.cbi.pku.edu.cn/)可视化并分析马铃薯StCDPKs基因的外显子-内含子结构[31].用在线服务器MEME server(http://alternate.meme-suite.org/)对已鉴定StCDPKs基因的Motif进行检测,并在InterPro中识别保守结构的名称.用MEGA 6.0软件构建马铃薯StCDPKs蛋白序列的系统进化树,并用TBtool绘制成可视化图谱.选取27个马铃薯CDPKs基因、36个拟南芥CDPKs基因和28个番茄CDPKs基因蛋白序列,用MEGA 6.0构建系统发育进化树.
1.4 马铃薯StCDPKs基因的转录分析
从PGSC(the potato genome sequencing consortium)数据库(http://solanaceae.plantbiology.msu.edu/pgsc_download.shtml)中下载相应的马铃薯SCDPKs基因的RNA测序数据.使用TBtools软件绘制马铃薯StCDPKs基因组织特异性转录和不同胁迫下转录的可视化图谱.
1.5 qRT-PCR分析马铃薯StCDPKs基因的表达
用Linux提取马铃薯StCDPKs基因的CDS序列,并利用DNAMAN设计定量扩增的引物.用TIANGEN的植物总RNA提取试剂盒(DP432)分别提取不同样品的总RNA,用cDNA合成试剂盒(KR118)反转录合成cDNA,用qRT-PCR试剂盒进行定量检测.使用2-ΔΔCT方法分析StCDPKs基因的相对表达量,用Origin软件作图.
2 结果与分析
2.1 马铃薯StCDPKs基因的筛选和理化性质
本研究共鉴定出27个马铃薯StCDPKs基因,其中StCDPK3基因和StCDPK5基因已在NCBI中注册.使用Fantino等[26]的命名方法,将Fantino等[26]中没有的一个基因命名为StCDPK27.27个基因中5个基因拥有两个转录本,可能是同一个基因通过不同的剪接形式翻译成两种蛋白质.马铃薯StCDPKs基因大多数有4个EF-hand,并两两成对,但StCDPK27基因只含有2个EF-hand(表1),说明StCDPKs在EF-hand结构域处有很高的保守性.大多数StCDPKs蛋白质的氨基酸为500~640个,只有3个蛋白氨基酸少于500个.1和10号染色体各分布有5个StCDPKs基因成员,其余StCDPKs基因分布于其他染色体,而9号染色体上没有StCDPKs基因.StCDPKs蛋白质中有47%的蛋白是稳定的.大多数StCDPKs蛋白呈中性,但是StCDPK25a和StCDPK26两个蛋白呈碱性,StCDPK6b呈酸性.说明StCDPK6/25/26基因的不同剪接体在不同pH条件下具有特定的功能.
2.2 马铃薯StCDPKs基因的结构、Motif与进化树分析
基因结构分析显示显示,11个马铃薯StCDPKs基因(CDPK4/6/8/11/14/15/16/17/18/19)位于同一分支上,它们大多含有7个外显子,只有StCDPK15基因含有8个外显子.StCDPK17/18/19基因的Motif中缺少前端一个未知功能结构域(图1-B),且这3个基因在同一分枝上.StCDPK1基因是最长的,拥有9个外显子,保守结构域与StCDPK6基因的相同.与StCDPK1基因在同一进化分枝上的基因都含有8个外显子,但StCDPK13和StCDPK10基因含有9个外显子.StCDPK12基因与StCDPK17/18/19基因的保守结构域相似.StCDPK20/21/22/23/245个基因进化有差异,StCDPK21基因含有7个外显子,而StCDPK20基因含有9个外显子,其余基因都含有8个外显子.这5个基因编码的蛋白在C末尾都多出一个Motif.说明StCDPKs基因的结构和Motif分布差异性较小.StCDPK27基因含有8个外显子,Motif和保守结构域与其他类似,主要结构缺少2个EF-hand,进化上不属于现有的4类亚家族,而与Ⅳ型亚家族接近,说明StCDPK27基因属于StCDPKs基因家族,但在进化上与其他基因有所区别.
以36个拟南芥、29个番茄和27个马铃薯的CDPKs蛋白构建进化树表明,进化树拥有4个亚家族(Ⅰ-Ⅳ型),而Ⅱ型分为两个小亚家族.StCDPK4/5/6/7/8/11/14/15/16/17/18/19聚类于Ⅰ型亚家族.StCDPK1/2/3/12与StCDPK9/10/13聚类于Ⅱa型和Ⅱb型亚家族,StCDPK20/21/22/23/24聚类于Ⅲ型亚家族,StCDPK25/26聚类于Ⅳ型亚家族,而StCDPK27没有聚类于4个亚家族中,但与Ⅳ型基因相近(图2).
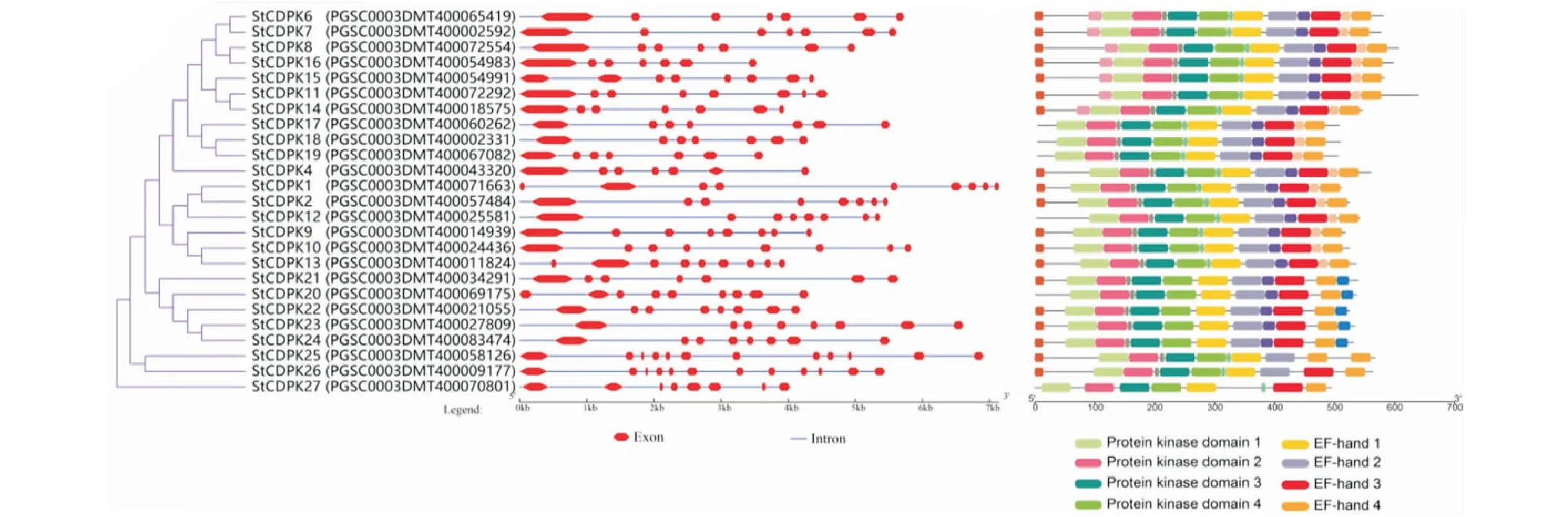
红色的不规则六边形代表外显子,蓝色的直线代表内含子.每个motif信息都由InterPro进行注释.The irregular hexagons in red represent exons,and the straight lines in blue represent introns.Each motif information is annotated by InterPro.图1 25个StCDPKs基因的结构(A)和Motif分布模式(B)Figure 1 Structural (a) and motif distribution patterns (b) of 25 StCDPKs genes
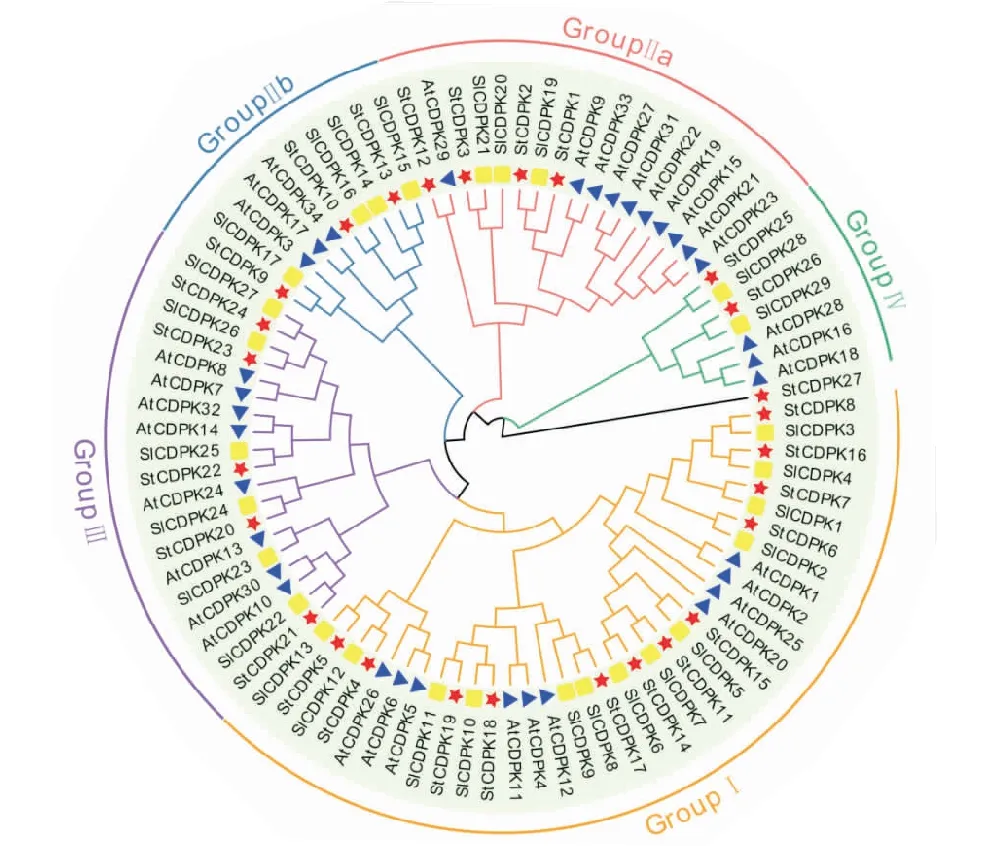
红色五角星为马铃薯,蓝色三角形为拟南芥,黄色正方形为番茄.The red pentagram represents potato,the blue triangle represents Arabidopsis,and the yellow square represents tomato.图2 36个拟南芥、29个番茄和27个马铃薯CDPKs蛋白通过MEGA6.0构建的系统进化树Figure 2 Phylogenetic tree constructed by MEGA6.0 using 36 A.thaliana,29 tomato,and 27 potato CDPKs protein sequences
2.3 马铃薯StCDPKs基因转录本分析
用马铃薯数据库PGSC中的RNA测序数据制作不同组织和不同胁迫下StCDPKs基因表达的可视化热图(图3~4).结果显示,StCDPK2基因在叶和叶柄中有较高的表达水平,果实和花中基本不表达.StCDPK4基因在根、叶片、萼片、心皮中表达较高,在块茎中表达较低,而在其他组织中有中等表达水平.StCDPK22基因在芽、叶柄、匍匐茎、心皮和未成熟果实中有较高的表达水平,而在花中有较低的表达水平.StCDPK25基因在块茎和匍匐茎中表达较高,其余组织为中下表达水平.StCDPK13和23基因在成熟花中有较高表达水平,这表明马铃薯StCDPKs基因家族可能参与调控马铃薯的生长发育.StCDPK22基因在生物和非生物胁迫中的表达量高于其他基因,StCDPK2/4/6/21/25基因表现出较高的表达水平,而7个基因(StCDPK1/7/9/17/18/23/24)的表达水平偏低,其余的12个StCDPKs基因在不同胁迫中基本不表达,这表明StCDPKs基因家族成员可能与马铃薯响应各种应激胁迫有关.

蓝色阴影表示基因的低表达,红色阴影表示基因的高表达,中间转录水平用黄色表达.Blue shading indicates low gene expression,red shading indicates high gene expression,and intermediate transcription levels are expressed in yellow.图3 StCDPKs基因在马铃薯不同组织器官中的表达量Figure 3 StCDPKs gene expression in different tissues and organs of potato

红色为高表达,蓝色为低转录或不表达,黄色为中度表达.每次处理时间为24 h.Red is high expression,blue is low transcription or no expression,yellow is moderate expression.Each processing time is 24 h.图4 StCDPKs基因在不同处理下的表达模式Figure 4 StCDPKs gene expression pattern under different treatments
2.4 马铃薯StCDPKs基因的表达模式分析
通过qRT-PCR分析马铃薯StCDPKs基因在根、茎、叶、花和块茎中的表达水平(图5),结果显示,StCDPK1/2/19基因在花中的表达量高于叶中,其他3个组织中的表达量都低于叶片.与热图中表达量有所差异:StCDPK1基因根、茎、花和块茎中的表达量均高于叶片;StCDPK2基因在块茎中的表达量高于叶片,而根、茎和花中则低于叶片;StCDPK19基因在根中的表达量低于叶片,茎、花和块茎中的表达量均高于叶片.在根中StCDPK8/10/11/21/25基因表达量低于叶片中,但是这5个基因在其他组织中的表达都高于叶片中.StCDPK17/20/223个基因与StCDPK8/10/11/21/25基因表达有类似的模式,这3个基因在茎中有较高的表达,而在根、叶、花和块茎中表达较低.StCDPK23基因相对于叶片,在花和茎块中表达较高,而在根和茎中较低.StCDPK12/16/18/244个基因在花和茎中的表达量高于叶片中的表达量,而在根和块茎则相反.StCDPK13/142个基因在茎和块茎中相对于叶片有较高的表达,但在根和花中较低.


显著性分析使用邓肯法,P<0.05.Significance analysis using Duncan method,P<0.05.图5 27个StCDPKs基因在不同组织中的相对表达量(以叶片的表达为对照)Figure 5 The relative expression levels of 27 StCDPKs genes in different tissues were compared with those in leaves
StCDPK3/4/6/7/26/276个基因在叶片中的表达量均低于根、茎、花和块茎,而StCDPK9和StCDPK15基因在叶片中表达量最高.StCDPK4/6/7基因在二倍体测序中的表达量与试验相符,说明二倍体马铃薯和四倍体马铃薯在基因表达上具有差异性.
用200 mmol/L的NaCl模拟盐胁迫处理马铃薯,在不同处理时间点采集样品进行StCDPKs基因定量分析.结果表明,StCDPK8/10/11/18/22基因在盐胁迫不同时间表达量都高于对照,StCDPK20/21基因在盐胁迫初期表达较高,但在16 h时表达量降低.StCDPK2/3/15/17/24/25/27基因在盐胁迫后期具有较高的表达量,而StCDPK9/12/13/19/23/26基因则在前期表达较高到后期降低.StCDPK1/4/7基因在不同时间表达不相同,StCDPK5/6/14基因在前期和后期表达高于中期,但StCDPK16基因则相反(图6).
3 讨论
CDPKs蛋白激酶在植物生长发育和逆境胁迫应答过程中具有极其重要的作用.目前,CDPKs基因在大约60多种植物中被鉴定出来,其中在拟南芥中鉴定出34个,烟草中鉴定出15个[32],番茄中鉴定出29个[22].本研究通过隐马尔科夫模型(PF00069和PF13499),使用HMMER程序比对目前最新的马铃薯全蛋白序列鉴定了马铃薯StCDPKs基因成员.Fantino等[23]从不同数据库中检索得到马铃薯CDPKs基因,其鉴定的StCDPK23和24基因含有3个EF-hand结构域,但这两个基因拥有4个EF-hand结构域.StCDPK22基因从进化上看拥有4个EF-hand结构域,但在InterPro预测拥有2个EF-hand.StCDPK27基因则有2个EF-hand结构域.27个基因中有5个(StCDPK4/6/13/25/27)含有2个转录本,其中StCDPK6/25基因两个转录本之间的氨基酸数各相差260和245个氨基酸,可能具有功能的特异性.


显著性分析使用邓肯法,P<0.05.Significance analysis using Duncan method,P<0.05.图6 27个StCDPKs基因在盐处理下不同时间的相对表达量(以0 h的表达为对照)Figure 6 The relative expression levels of 27 StCDPKs genes at different time under salt treatment were compared with that at 0 h
StCDPKs基因的EF-hand结构域具有较高的保守性,StCDPK1-5基因的保守结构域和Motif没有不同,但是在位置上有所差异.内含子和外显子个数差异不大,大部分基因含有7或8个外显子,而StCDPK3/11基因的外显子多达12个.梨的CDPKs基因大多数也有7或8个外显子,最多有11个[33].构建进化树分析了拟南芥、番茄和马铃薯CDPKs基因编码的蛋白之间的进化关系,从进化分枝上看StCDPKs基因分布于各个亚家族,其中Ⅰ型最多,而且相近的基因具有相似的功能.StCDPK4/5基因都具有抗病菌能力[34-35],而AtCDPK8/10基因能够提高植株的抗旱性[13,36].
马铃薯StCDPKs基因空间表达转录可视化图谱可分为3类.第一类表达具有明显的组织特异性,如StCDPK10/13/20基因在叶、茎、根等组织中很少有表达,而仅在成熟的花中具有较高的表达.小麦TaCDPK3基因在未成熟种子中未检测到表达,而TaCDPK13基因在叶、幼穗和未成熟的种子中有表达,而在根、茎中没有表达[37].第二类在各组织中表达都较低或者不表达,表达没有特异性.如StCDPK8/11/12/14/15/16基因等在不同组织中都表现出较低表达水平或不表达.拟南芥CDPK3/8/12基因在根、茎、叶和花中具有一致的表达性[38].第三类在各组织中都有转录而且具有明显的差异性.如StCDPK25基因在块茎中表达最高,在其他组织中表达没有一致性.以上结果表明,马铃薯StCDPKs基因在不同组织中各自的功能可能不同.
小麦TaCDPK7/12基因在盐胁迫、干旱胁迫和ABA信号通路扮演着重要角色[39].拟南芥AtCDPK1基因与AtCDPK1a基因在盐胁迫下表达量显著上调[40].盐胁迫条件下,相对于0 hStCDPK8/10/11/18/22基因在其他时期表达量均高,StCDPK8/10/11/18/22基因可能在盐胁迫的整个时期对提高植株的耐盐性具有积极作用.部分基因的表达模式恰恰相反,例如:StCDPK27基因在1 h表达降低,但随表达时间增加均增加,而StCDPK9/26基因的表达模式与之刚好相反,同样的StCDPK5/19与StCDPK16和StCDPK13与StCDPK17/14基因表达模式也相反.StCDPK2/3/15基因在盐胁迫前期表达较低,而到后期则有较高的表达.这说明在盐胁迫下,StCDPK8/10/11/18/22基因在胁迫整个时期发挥着重要作用,而StCDPK2/3/15基因在胁迫后期发挥着作用.
4 结论
本研究鉴定出27个马铃薯StCDPK基因成员,可分为4个亚家族,其基因结构相似,催化区域的EF-hand具有较高的保守性.StCDPKs基因在不同组织和盐胁迫下具有不同的表达模式.
猜你喜欢
电子科技大学学报(2022年5期)2022-10-29
今日农业(2021年15期)2021-11-26
天津市教科院学报(2021年5期)2021-11-10
生物学通报(2021年9期)2021-07-01
中国生殖健康(2020年4期)2021-01-18
种子(2019年7期)2019-08-13
中国生殖健康(2018年4期)2018-11-06
现代园艺(2017年19期)2018-01-19
中国医学创新(2017年7期)2017-03-31
江苏农业科学(2016年8期)2017-02-15
