
水稻粒型基因克隆研究进展及育种应用展望
2020-11-23 08:37康艺维陈玉宇张迎信
中国水稻科学 2020年6期
康艺维 陈玉宇 张迎信
(中国水稻研究所 国家水稻改良中心/水稻生物学国家重点实验室,杭州 311401;*通信联系人,E-mail:zhangyingxin@caas.cn)
据国家统计局数据,2019 年我国的稻谷播种总面积是 2969 万 hm2,占总粮食作物播种面积的25.58%,表明水稻生产在我国粮食生产中具有重要地位(http://www.stats.gov.cn)。随着我国人民生活水平的提高,稻米需求呈现多样化趋势,在产量仍作为重要指标的同时,稻米品质也受到越来越高的重视[1-3]。水稻产量的决定因素包括单株有效穗数、每穗实粒数和粒重,粒重主要取决于粒型和充实度。根据籽粒的三维结构将粒型构成划分为粒长、粒宽和粒厚[4]。细长粒稻米通常表现较好的外观品质,因此粒型也是影响稻米品质的重要因素。著名的 Basmati 系列香米[5]、我国广东的丝苗品种(http://www.ricedata.cn/variety)都是长粒优质稻的典型代表,且近年来大面积应用的优质不育系如野香 A[6]和泰丰 A[7]等也具有典型的长粒表型。近几十年来,研究者们定位和克隆了许多水稻粒型相关基因,极大地促进了粒型形成分子遗传机制的研究,并且为利用分子育种技术准确、快速、高效地对水稻粒型改良提供了可能。本文综述了已克隆的水稻粒型基因/QTL 及籽粒分子调控机制,并重点对粒型 QTL 进行了育种应用现状及潜力评价,以期为我国水稻育种家提供分子育种的参考信息。
1 水稻粒型QTL 研究进展
水稻粒型是典型的数量性状,研究者们普遍认为粒长、粒宽、长宽比一般受多基因控制,也有少数受单基因、双基因控制。还有研究表明控制粒型各性状的基因还存在互补和累加效应[8-10]。随着水稻基因组测序的完成,研究人员利用不同的遗传群体,如F2群体、回交群体(BC)、加倍单倍体群体(DH)、重组自交系(RIL)、染色体片段置换系(CSSL)、导入系(IL)和剩余杂合体(RH)等定位了大量与粒型相关的QTL。据不完全统计,在水稻12 条染色体上已定位了500 多个粒型相关QTL(http://www.ricedata.cn/index.htm)。利用存在于栽培品种间的自然变异,研究人员通过正向遗传学的方法已经克隆了18 个与水稻粒型相关的主效QTL,分布在第 2、3、4、5、6、7、8 和 9 染色体上(图1)[11-22]。
GW2、qSW5/GW5/GSE5、GS5、GW8和TGW2主要控制粒宽。GW2克隆自大粒粳稻品种WY3,编码一个环型E3 泛素连接酶,亚细胞定位于细胞质中[23]。GW2提前终止型的功能缺失导致颖壳细胞分裂速度加快、颖壳变宽,有利于胚乳灌浆,增加粒重,最终产生宽大的籽粒。克隆自宽粒粳稻品种日本晴、Asominari 等的qSW5/GW5/GSE5编码一种新型的油菜素内酯(Brassinosteroid,BR)信号调控因子[24-26]。功能缺失或表达量下降型的基因负调控小穗颖壳细胞的增殖,导致宽粒和重粒。位于第5 染色体短臂控制粒宽的GS5,克隆自籼稻恢复系珍汕97,它编码的蛋白含有PF00450 结构域,属于肽酶S10 家族的丝氨酸羧肽酶[27]。GS5表达量的增加可促进内外稃的细胞增殖和扩张,正向调控籽粒大小。GW8/OsSPL16位于第8 染色体长臂上,供体亲本为Basmati 385,是调控水稻粒型和稻米品质的QTL[28]。在Basmati 水稻品种中,OsSPL16启动子区10 bp 的缺失使OsSPL16表达下调,导致籽粒变长且外观品质变好。GW8和GS5都是由于启动子区的核苷酸变异影响了粒宽和粒重。TGW2是最新克隆的控制粒宽和粒重的QTL,亲本为93-11 和培矮64S,编码细胞数目调控因子OsCNR1,负调控粒宽和粒重。TGW2基因启动子上游1818 bp 处的碱基替换(G→A)可增强TGW2的表达[22]。
主要控制水稻粒长的9 个QTL 包括GS3、qGL3/GL3.1/qGL3-1、TGW6、OsLG3、GLW7、GL4、qLGY3/OsLG3b/GW3p6、TGW3/qTGW3/GL3.3和GL6。GS3是水稻中首个被克隆的粒长和粒重QTL[29-30]。GS3功能缺失等位基因单碱基替换(C→A)导致翻译提前终止,产生一部分氨基端具有类γ 结构域的蛋白,使籽粒变长。qGL3/GL3.1/qGL3-1克隆自粳稻大粒品种N411、Waiyin-3(WY3)、CW23 等,编码含有两个Kelch 功能域的蛋白磷酸酶,负调控水稻粒长[12,31,32]。TGW6的定位群体是用Kasalath 作供体亲本,日本晴作受体亲本构建的回交自交系[33]。功能缺失的TGW6通过影响胚乳细胞发育进程限制了籽粒长度进而负调控籽粒大小。Yu 等[13]用Ho-LAMap 方法从品种SLG-1 中克隆到OsLG3。OsLG3编码APETALA2/乙烯反应元件结合蛋白,通过促进细胞数目的增长正向调控粒长,且在不影响稻米品质的同时提高水稻产量。实验所用长粒品种 IRAT109、SLG-1、Haobuka 都有相同的启动子序列且不同于短粒品种,说明OsLG3启动子区的自然变异是导致粒长和粒重变化的原因。GLW7是第一个通过GWAS 技术在水稻中克隆到的QTL,编码OsSPL13,通过促进颖壳细胞扩大正向调控籽粒大小[34]。GL4是克隆自非洲野生稻W1411 的一个调控粒长和粒重的QTL,通过调控内外颖纵向细胞的伸长,控制非洲栽培稻的粒长,同时也能调控种子落粒性[35]。
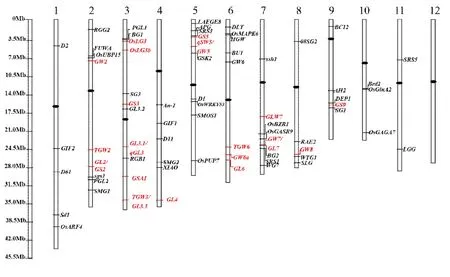
图1 已克隆水稻粒型相关基因在12 条染色体上的分布Fig.1.Distribution of cloned genes for grain size on rice chromosomes.
qLGY3/OsLG3b/GW3p6的供体亲本为 SLG-1、L-204、广占63-4S 等长粒品种[14-16]。该基因编码的转录因子OsMADS1 是G 蛋白下游调控因子。单倍型和基因渗入分析表明该基因经过驯化,OsMADS1lgy3在热带粳稻中的存在能使粒长明显增加。TGW3/qTGW3/GL3.3克隆自大粒品种 JZ1506、CW23 和南洋占等,编码类 SHAGGY41 激酶OsSK41/OsGSK,OsSK41 通过磷酸化 OsARF4 来负调控颖壳的细胞扩张进而调控粒长[17-19]。GL6供体亲本为W1943,受体亲本是广陆矮4 号[20]。GL6编码植物特异的 PLATZ 转录因子,调控水稻粒长和小穗数,通过促进幼穗细胞的增殖正调控粒长。GL6编码的蛋白通过与RPC53 和TFC1 互作,参与介导 RNA 聚合酶Ⅲ促进核糖体合成,这丰富了调控水稻籽粒大小的分子机制。
同时控制粒长和粒宽的 QTL 为GL2/GS2、GL7/GW7、GW6a和GS9。GL2/GS2克隆自大粒品种RW11 和“宝大粒”(BDL)等,是一个半显性主效QTL,编码的转录因子OsGRF4 属于GRF 家族蛋白成员,主要通过促进细胞扩张和少量细胞增殖负调控籽粒大小[23,36-37]。OsGRF4 的氨基酸突变改变了OsmiR396 的靶向位点,进而破坏OsmiR396对OsGRF4 的抑制作用,使OsGRF4超量表达,导致籽粒变大,产量提高。GL7/GW7的供体亲本为P13、天丰 A。GL7的串联重复片段可以上调基因的表达水平,调控细胞的纵向伸长,使水稻籽粒变得更细长,也提高了稻米的外观品质[38]。GW7编码一个TONNEAU1 募集基序蛋白,其表达量上调,能促进籽粒的纵向细胞分裂,并减少横向细胞分裂,导致籽粒变得细长[39]。GL7/GW7控制细胞分化的方式还需要进一步的研究。GW6a编码的OsglHAT1蛋白具有内源组蛋白乙酰转移酶活性,能通过增加细胞数目和加速灌浆速率来增大颖壳,从而增加粒重和产量[40]。尽管GW6a在粒重、产量和植株生物量方面具有有利于农艺性状的效应,但能使GW6a表达量提高的等位基因还未得到利用。Zhao 等[21]用Qingluzan 11 和日本晴构建的染色体片段置换系CSSL N138 与日本晴杂交构建定位群体并克隆得到GS9。发现GS9通过调控颖壳细胞的分化负调控籽粒大小,将gs9等位基因导入武运粳27 中,籽粒变细长,稻米的外观品质显著提高。
2 水稻粒型调控的分子机制
稻米生长在颖壳内,颖壳提供给稻米生长发育的环境但也限制稻米的生长。颖壳受母体组织控制,分子机制涉及一些信号通路,如泛素蛋白酶体途径、G-蛋白信号通路、MAPK 信号通路、植物激素信号和转录因子等[11,41-42]。科学家们从化学诱变和T-DNA 插入等突变体库中鉴定了一些突变体,研究这些突变体有助于完善粒型形成分子机制及调控网络的研究。
参与泛素蛋白酶体途径的基因:WTG1编码与人类OTUB1 同源的蛋白,具有泛素化活性,抑制OsSPL14 的K63 位多聚泛素化。wtg1-1突变体通过减少颖壳细胞的扩张产生宽、厚、短的籽粒[43],并且茎秆变粗、穗变大,被认为是新株型主效 QTLNPT[44]。OsUBP15表达下调或功能缺失都能产生更窄小的籽粒[45]。此外,还有已克隆的编码环型 E3泛素连接酶的 QTLGW2,通过将底物锚定到蛋白酶体将其降解,负调控细胞分裂。
参与G 蛋白信号通路的基因:D1编码GTP 结合蛋白(G 蛋白)的α 亚基,功能缺失型突变体的G 蛋白失活,籽粒短而圆[46]。G 蛋白β 亚基RGB1的敲除转基因植株籽均变小且不育种子增多[47]。DEP1/qPE9编码的蛋白包含一个完整的氨基酸类γ结构域但缺乏羧基端富半胱氨酸区域,导致圆锥花序直立,籽粒大小和粒重略微降低,被认为是一种功能获得型等位基因[48]。最近的研究表明,过表达DEP1会导致籽粒变大,而下调或者敲除DEP1会导致籽粒变小和直立的圆锥花序[49]。RGG2编码G蛋白γ 亚基,可参与赤霉素合成途径,负调控水稻籽粒大小和产量[50]。在已克隆 QTL 中,GS3是通过此途径调控籽粒大小的。
参与 MAPK 信号通路的基因:OsMKK10,OsMKK4和OsMAPK6是一个控制水稻籽粒大小的级联反应。功能缺失突变体smg2-1/OsMKK10,smg1-1/OsMKK4和dsg1/OsMAPK6由于减少了颖壳细胞的增殖而使籽粒变小[51]。GSN1编码丝裂原活化蛋白激酶磷酸酶 OsMKP1 , 负调控OsMKKK10-OsMKK4-OsMPK6 级联途径[52]。OsRac1通过影响细胞分裂来调控水稻籽粒大小和产量,与OsMAPK6互作,OsMAPK6可以调节水稻的细胞分裂和籽粒大小[53]。
参与植物激素信号途径的基因[11,54]:位于第1染色体的D61/OsBRI1[55]和D2[56];第 2 染色体的PGL2/OsBUI1[57];第 3 染色体的BG1[58]和PGL1[59];第 4 染色体的XIAO[60]和D11[61];第 5 染色体的GSK2[62]、APG[59]和SMOS1[63];第 6 染色体的DLT[64]和BU1[65];第7 染色体的OsBZR1[66];第8 染色体的OsBAK1/08SG2[66,67]和SLG[68];第 9 染色体的SG1[69]。其中,与 BR 信号相关的基因最多,有D61/OsBRI1、OsBAK1/08SG2、OsBZR1、GSK2、XIAO等。在已克隆QTL 中,GS5 蛋白数量增加会抑制OsBAK1-7 与OsMSBP1 互作而产生的胞吞作用,促使BR 信号增强,进而导致籽粒变大。
转录调控因子OsGIF1编码GRF 互作蛋白[70],调控胚乳中淀粉的形成。gif1突变体的粒重变轻。AH2编码SHAQKYF 类MYB 转录因子,是调控水稻颖壳外层细胞发育的基因,并对籽粒产量和品质均有影响[71]。在已克隆的粒型相关 QTL 中,多数为转录因子(表1)。
miRNA 是植物生长和发育的重要调节因子,近来发现在籽粒调控中也起到重要作用。miR1432 负调控水稻籽粒大小[72]。突变体fuwa与野生型植株相比株高略矮,茎秆变粗壮,分蘖数减少,穗变短但直立、紧密,籽粒变宽、变厚、变短,胚乳质量增加、灌浆速率加快[73]。HGW调节水稻抽穗期和粒重[74],在杂合基因型的突变体中,由于颖壳细胞数目减少使籽粒宽度减少,千粒重下降。
水稻籽粒大小由一个复杂的网络来控制,这个网络融合了多种发育和环境信号。尽管最近的研究已经明确了一些籽粒大小关键调控因子和分子调控途径,但对整个调控网络的认知仍然有限和零散。因此,有必要探索已知籽粒大小调控因子的上下游成分,以及已确定的调控通路之间可能的联系。
3 主要粒型基因的功能标记
分子育种为精确定向改良某一性状提供了有力的工具,并正在成为水稻育种的新兴方向之一。分子标记辅助选择(marker-assisted selection, MAS)是利用与目标基因共分离或连锁的分子标记进行辅助选择育种的方法,目前已在作物育种中得到了广泛应用,在回交育种中具有定点且高效的优点,且不受基因类型的限制。例如,聚合OsLG3SLG和gw8Basmati的品种云广8 号,不仅产量提高,还实现了米质的优化[13]。聚合OsMADS1lgy3和dep1-1、gs3基因是同时提高水稻产量和米质的有效策略[14]。功能标记是基于基因内部序列变异而开发的一种共分离分子标记,能准确检测出水稻材料中的目标基因且能高效地筛选出有利基因和基因型。
研究者根据已克隆的基因设计了功能标记,如GS3第 2 外显子的 CAPS 功能标记 SF28[75]、GS5的 CAPS 标记 GS5-1 和 dCAPS 标记 GS5-2[76]、GW8启动子的 InDel 标记 GW8-1[77]、GS9的 InDel 标记NIL-F、NIL-R 和 5′-R[21]、qGL3第 10 外显子的ARMS-PCR 标记 qGL3-SNPF1/qGL3-SNPR1[78]。Zhang 等[79]利用 TD70 和 Kasalath 在GW2、GS3、qGL3、GS5、GW8和TGW6中的差异,开发了 dCAPS标记 GW2-F/GW2-R,CAPS 标记 GS3-F/GS3-R,dCAPS 标 记 qGL3-F/qGL3-R , SSR 标 记GS5-F/GS5-R,dCAPS 标记 GW8-F2/GW8-R2 和CAPS 标记TGW6-F/TGW6-R。其次,除了可根据基因旁侧序列设计易于操作的SSR 或InDel 连锁标记,根据已发表的序列变异可以有目的性、准确地设计基因功能标记。qSW5/GW5/GSE5基因可根据粳稻1 212 bp 或籼稻950 bp 缺失设计InDel 功能标记,GW6a基因ORF 在日本晴与Kasalath 之间存在9 个SNP 的差异,可用于设计CAPS 或dCAPS 标记,GL7基因可根据拷贝数变异设计显性标记,OsLG3基因可根据启动子5 bp 缺失设计InDel 标记,GL4则可根据大粒基因型第1 外显子6 bp 插入设计InDel 标记,TGW3/qTGW3/GL3.3可根据大粒基因型中第3 和第4 外显子的大片段缺失设计InDel 标记,GS2可根据 SNP(TC487-488AA)设计 CAPS 或dCAPS 标记,qLGY3/OsLG3b/GW3p6基因可利用紧密连锁标记XP22、XP23 或第7 内含子的碱基缺失设计InDel功能标记,GLW7可根据基因ORF区SNP设计 CAPS 或 dCAPS 功能标记。GL6可根据第 1内含子 15 bp 的缺失设计 InDel 功能标记。TGW2基因可根据启动子上游1818 bp 的单碱基突变(G-A)设计CAPS 或dCAPS 标记。
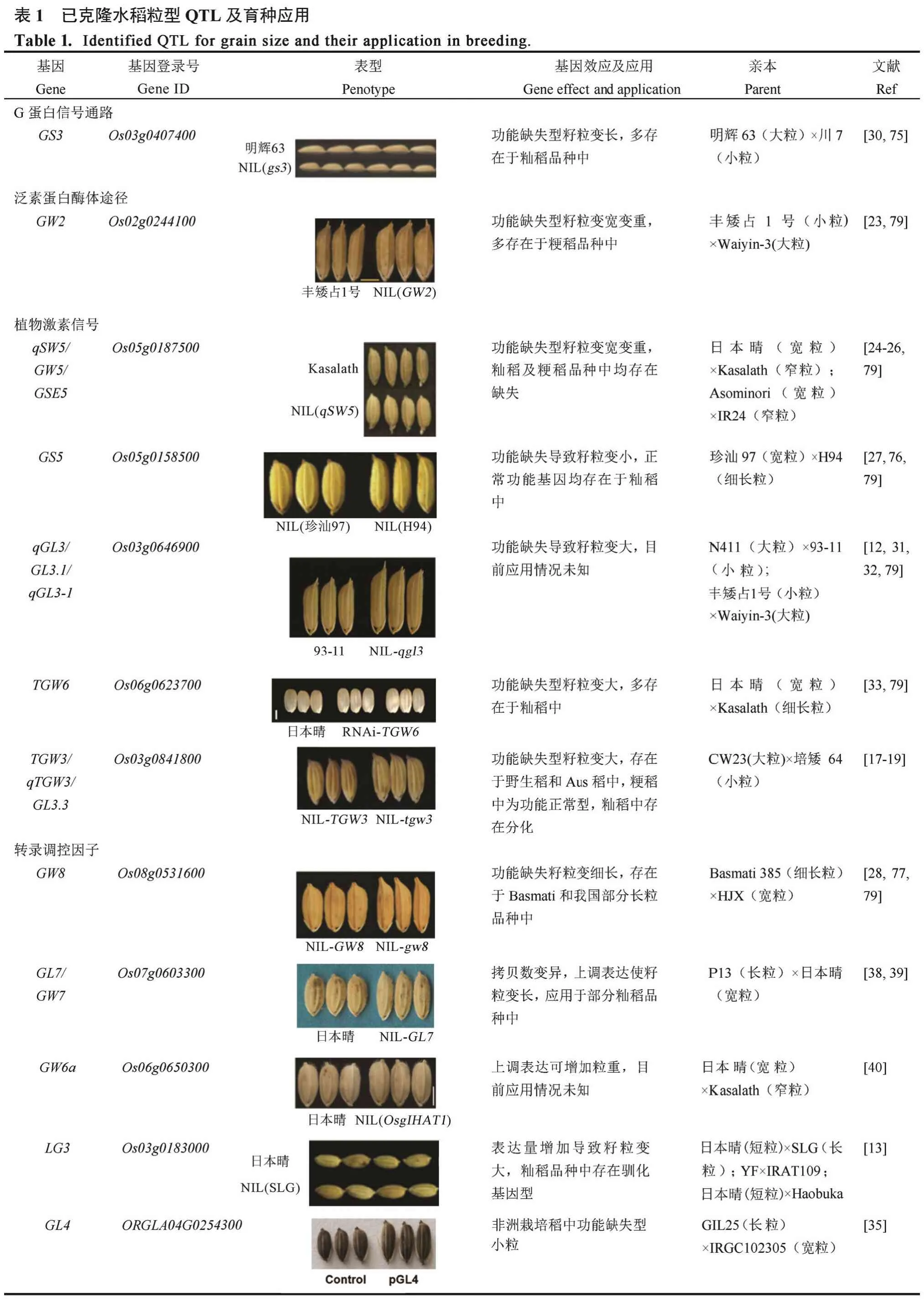
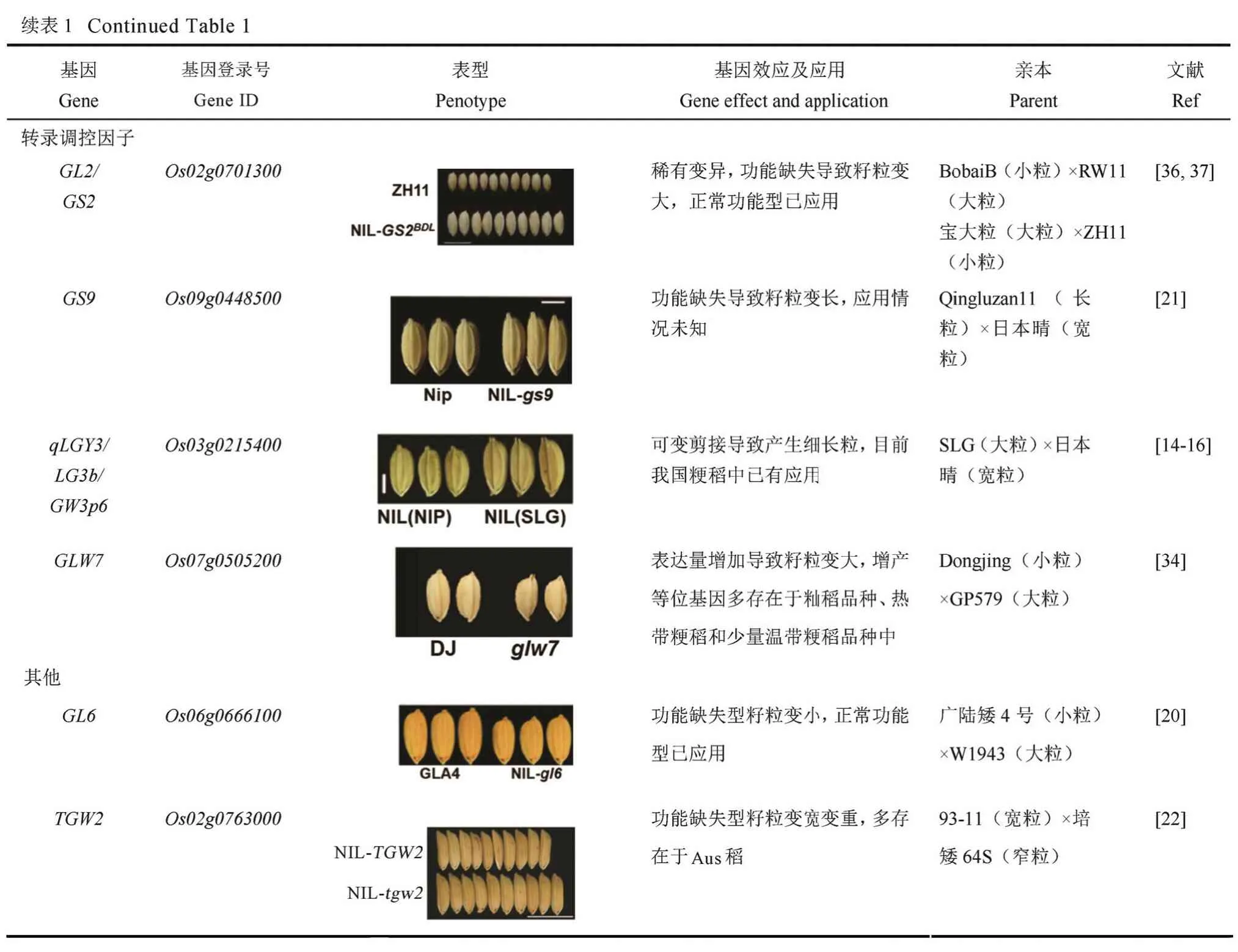
4 已克隆粒型基因的育种应用
?
?
评估18 个已克隆的水稻粒型QTL,并开发相关基因的功能标记,可以帮助育种家们合理选择并聚合有利等位基因,加快育种进程。在已有的报道中,对于粒宽和粒重基因GW2,与轮回亲本FAZ1,NIL(WY3)粒宽增加0.6 mm,粒厚增加0.3 mm,千粒重增加9 g,单株产量增加5 g[23]。籽粒大小调控基因GS5:与NIL(H94)相比,珍汕97 粒宽在增加0.3 mm, 粒重增加1.6 g,单产增加1.4 g[80]。粒重基因GW8:轮回亲本 HJX749 粒宽和粒重较GW8表达量低的 NIL(Basmati 385)分别增加了0.3 mm 和 3 g,单产增加 5 g[28]。粒宽粒重基因TGW2:93-11 的tgw2等位基因导入培矮 64S,单产提高13.6 %[22]。如表 1 所示,在这些 QTL 中,GS3、TGW6、GL6、OsLG3和GS5增产等位基因目前已广泛应用于籼稻品种中,qSW5/GW5/GSE5增产等位基因已在粳稻品种中得到广泛应用;GLW7增产等位基因多存在于籼稻品种、热带粳稻和少量温带粳稻品种中;GW8、GL7/GW7和qLGY3/OsLG3b的优质或增产等位基因可能在我国部分水稻品种中存在;GL2/GS2增产突变和GS9优质突变基因型为稀有突变,TGW3/qTGW3/GL3.3和TGW2增产突变型主要存在于野生稻及Aus 稻中;GW2、GW6a、GL4和qGL3/GL3.1/qGL3-1增产或优质基因型在生产中情况未知,需要更多的相关研究结果才能进行评估。综合上述信息,qLGY3/OsLG3b/GW3p6、GL2/GS2、TGW3/qTGW3/GL3.3和TGW2的增产等位基因在增加粒重的育种目标中具有较大的应用潜力,qLGY3/OsLG3b/GW3p、GW8、GL7/GW7和GS9的优质等位基因在优质稻育种方向中具有很好的应用前景,GW6a、GL4和qGL3/GL3.1/qGL3-1增产或优质等位基因可能具有较大的应用潜力。另一方面,在籼稻中广泛应用的高产或优质等位基因可能在粳稻品种改良中具有较大的潜力,如GLW7、GS3、TGW6、GL6、OsLG3和GS5;GW2和qSW5/GW5/GSE5在籼稻中多表现细长粒和较低的千粒重,鉴于目前籼稻长粒优质稻的育种趋势,育种家在籼稻改良中是否利用GW2及qSW5/GW5/GSE5增产等位基因时可能存在一定的分歧,是追求更高的千粒重还是追求长粒型的外观品质取决于育种目标。
在分子标记辅助育种中应注意以下问题:1)目标基因及供体亲本的选择。研究者首先应通过测序鉴定或功能标记分析确定待改良亲本主要粒型基因的基因型,然后根据育种目标选择合适的目标基因及供体亲本用于分子设计改良。2)基因效应的可变性。一方面不同的遗传背景可能导致基因效应的改变,例如同一基因在籼稻与粳稻品种背景中的表型可能存在较大差异;另一方面,不同粒型基因之间的互作可能导致基因效应的变化,如qSW5对粒长的效应受到GS3的影响,GS3对粒宽的作用受到qSW5的影响[81]。因此,在进行分子标记辅助选择进行基因聚合的过程中,基因效应的评估是一项不可或缺的工作。3)连锁累赘的打破。由于连锁累赘现象的存在,在回交导入有利基因的同时往往伴随其连锁不利育种性状基因的导入。如第7 染色体上两个粒型基因GL7/GW7和GLW7遗传上连锁,另外它们还与水稻直立型密穗基因DEP2[82]、种子休眠和驯化调节子基因Sdr-4[83]连锁,分子标记辅助选择应用时应根据育种目标和亲本基因型进行育种设计,在回交选择过程中利用大样本量的分离群体打破不利连锁,选择保留有利等位基因。4)导入粒型基因可能改变株叶形态或其他性状。水稻粒型的改变是由于颖壳细胞大小或细胞数目的改变引起[84,85],往往伴随水稻株高、穗长、叶长或叶宽等性状的改变,进而导致株叶形态、光合效率或抗倒伏等其他性状发生变异,如GS3除了可以调控粒型,对株高和穗长均有影响[30],而GL4除了可以控制粒长,还能影响水稻的落粒性[35]。因此,综合农艺性状的考查是评估目标基因分子育种应用中必不可少的环节。
5 展望
5.1 加速水稻粒型相关基因的克隆
水稻籽粒大小的调控机理还有许多未解之谜,而基因克隆是研究水稻籽粒大小调控机理的基础,因此有必要克隆更多的粒型调控基因。相比传统的图位克隆方法,近年多个新兴的基因定位方法为粒型基因的挖掘提供了更加简便快捷的途径。基于二代测序的集群分离分析法(Bulk Segregant Analysis,BSA)是近年来兴起的一种快速简单的性状定位的方法,通过在群体中挑选极端或能代表目标性状的个体构建 DNA 混池进行全基因组重测序及差异分析,从而快速实现对单一性状的初定位[86,87]。MutMap[88]将BSA 与全基因组重测序结合寻找候选基因,适合对化学诱变产生的隐性基因进行分析,例如可利用BC1F2分离的野生型表型群体和突变体表型群体构建两个 DNA 混池,将两个混池分别进行DNA 测序,计算SNP 在突变体混池和野生型混池出现的频率,可得到和突变表型连锁的染色体区段和可能的突变位点。应对不同的分离群体衍生出MutMap+[89]、MutMap-Gap[90]和 QTL-seq[91]等,它们不需要复杂的定位群体,可大大缩短基因定位周期。全基因组关联分析(Genome-Wide Association Study, GWAS)是利用自然群体或者人工群体在全基因组范围内找出存在的SNP、InDel、CNV 等变异类型,从中筛选出与目标性状相关的变异位点的方法[34,92-94]。如韩斌团队[34]通过对381 份粳稻品种籽粒大小性状的GWAS,快速将目标区间定位到第7 染色体上,SNP 峰值注释到11 个基因,结合表达量分析快速得到候选基因OsSPL13并通过了功能验证。虽然上述方法在基因挖掘方面存在快速高效的优点,但在涉及 QTL 的克隆时往往仍需结合传统图位克隆方法才能最终确定候选基因。
5.2 提高粒型分子设计育种的效率
随着多个水稻基因组测序的完成,如今数以百万计的遗传标记可以轻易获得,大量功能基因正被逐渐揭示和利用,精准基因组设计育种时代已逐渐来临。目前,大量基于芯片或测序技术的水稻分子育种产品被研发并进入应用阶段,主效基因相关标记的应用已逐渐形成规模[95]。若针对粒型性状,可特异性地设计包含多个粒型基因基于多重 PCR 或液相探针的基因型分型体系,可快速有效地分析目标基因的基因型,用于分子育种[96,97]。但目前无论是基于芯片还是二代测序技术,其分子育种应用最大的限制因素在于高昂的价格。另外,不同基因间的上位性效应评估不完善也限制了其分子设计特别是聚合育种的应用。随着技术的进步及水稻分子育种产业化的进程,我们可以乐观地期盼基于芯片或重测序的基因分型产品价格的大幅降低,直至可以大规模应用于水稻育种。
5.3 应用新兴技术改良粒型
此外,基因编辑(gene editing)是近年来兴起的一种精确打靶技术,在水稻中能快速有效地创制功能缺失型突变体[98]。CRISPR-Cas9 多基因编辑的功能具有快速高效地特点,如王春等利用该系统同时编辑REC8[99]、PAIR1[100]、OSD1[101]和MTL[101]四个基因,获得了可以通过种子进行无性繁殖的植株从而可以使优良F1杂交作物实现杂种优势固定,这一研究成果为基因编辑在水稻分子育种中的应用描绘了光明前景[102]。水稻粒型基因中的负调控因子适用于进行基因编辑以创制增产或优质的基因敲除后代,例如敲除GS3、GS2、TGW6、qTGW3、GSE5,籽粒千粒重增加,而敲除GW8可获得细长籽粒(表 1)。最近科学家开发的全新精准基因编辑工具PE(Primer Editors),无需额外的DNA 模板,从只能实现C-T/G-A 转换[103]到可实现12 种单碱基的转换和多碱基的精准插入与删除[104],这为基因编辑领域带来了重大变革,也为基因编辑在水稻分子育种的应用展示了光明的前景。值得注意的是,由于基因编辑经历转基因过程,应用时需首先去除转基因成分,需谨慎选用此方法;其次基因编辑存在基因组突变和脱靶现象,一般需将获得的阳性植株与背景亲本进行回交纯化后才能进行应用。
猜你喜欢
环球时报(2022-09-20)2022-09-20
麦类作物学报(2022年6期)2022-08-09
中国农学通报(2022年13期)2022-05-31
现代畜牧科技(2021年4期)2021-12-05
中国农业气象(2021年7期)2021-07-28
麦类作物学报(2021年3期)2021-05-25
今日农业(2020年24期)2020-12-15
山东建筑大学学报(2020年5期)2020-09-16
现代农业研究(2019年3期)2019-06-05
科学种养(2017年6期)2017-06-13
