
湿地松树木形成层恢复活动期、活动期和休眠期原始细胞超微结构变化
2020-11-30 00:36徐有明周彩霞陶吉云张菊花
林业科学 2020年10期
徐有明 周彩霞 林 汉 陶吉云 张菊花
(华中农业大学园艺林学学院 武汉 430070)
维管形成层活动是木材形成机制和木材基因调控研究的焦点(Oliveiraetal.,2009;Huangetal.,2014)。近年来,国内外学者在植物细胞壁与生物质能源利用方面开展了大量研究,其中树木形成层细胞的季节性分生分裂活动和木材形成规律得到广泛重视,如杉木(Cunninghamialanceolata)形成层季节性活动研究表明,维管形成层细胞数量与形成层区宽度正相关,形成层细胞数量和形成层细胞径向直径与木质部形成密切相关(Wuetal.,2016),形成层纺锤形细胞径向壁和弦向壁厚度差异显著,射线细胞壁为典型的复层结构(Songetal.,2011)。目前,在超微水平上研究针叶材树木管胞具缘纹孔和细胞壁结构方面的文献较多(Kwon,2014;Jelonek,2015;Sivakumaretal.,2016),但有关形成层原始细胞在不同时期超微结构的动态变化、活动方式以及与物候关系方面的研究较少(Songetal.,2011;Raoetal.,2001;Antonovaetal.,2006)。
湿地松(Pinuselliottii)是我国南方引种栽培较为成功的速生用材树种(黄齐等,2010;Zhongetal.,2001)。Harley等(2012)采用光学显微镜结合NIS元素图像分析系统研究美国佛罗里达湿地松不同阶段形成层细胞分生分裂活动和木材形成规律,结果发现湿地松生长季较长,从2月至12月连续11个月生长,但次年1月都会经历一段时间的短暂休眠(可以解释其生长轮具有明确边界的现象);短暂休眠状态下,湿地松个体间差异很小,太阳辐射是影响个体间生长的最主要气候因子,特别是形成层活动周期季节性变化与物候之间的关系密切。我国对湿地松的研究主要集中在人工林栽培管理、苗木培育、松脂、木材质量评价等方面(徐有明等,2002;肖兴翠等,2015),其树脂道形成过程中的超微结构变化已有报道(Guoetal.,2002),但有关形成层不同生长时期细胞发育分生分化过程中的超微结构变化及其管胞壁超微结构未见报道。
透射电镜技术是在纳米水平上研究维管形成层原始细胞和成熟木材细胞壁超微结构的良好工具(Hafrénetal.,2000;Singhetal.,2001;Frommetal.,2003)。本研究采用透射电镜观察湿地松整个年活动期内形成层原始细胞的超微结构变化,获取更多关于形成层季节性活动与次生木质部变化之间的信息,旨在丰富树木生长发育生物学知识,为湿地松人工林培育、材性材质改良和森林资源利用提供科学依据,也为其他树种的分子生物学研究提供有价值的细胞学参考。
1 材料与方法
试验材料取自华中农业大学校园湿地松人工林(113°41′—115°05′E,29°58′—31°22′N)。2016年3月—2017年3月,在湿地松人工林内选取5株生长健康的湿地松作为样株,其中3—8月每周采样1次,9—11月毎半个月采样1次,12月至次年2月每月采样1次。采样时,选取样株树冠中上部生长良好的2~3年生枝,截取其中部,切成若干0.5~1 cm小段,放入2.5%戊二醛固定液中,抽真空30 min后,置于4 ℃冰箱保存。
样品用氢氟酸、酒精甘油、乙二胺等化学药剂软化,使用双面刀片切成1 mm小块,放入新鲜固定液中,再抽一次真空,常压下固定2~3 h,用磷酸缓冲液冲洗4次,每次约30 min。洗涤后在由0.2 mol·L-1磷酸缓冲液配制的1%锇酸溶液中4 ℃下固定过夜。用丙酮系列脱水,纯丙酮脱水3次。按照丙酮与Spurr树脂3∶1、2∶1、1∶1、1∶2、1∶3渗透,再用纯树脂渗透,每次12 h。最后用树脂包埋,置于70 ℃烘箱中聚合2天。修块后用LKB-2188型超薄切片机切取连续超薄切片,100目铜网捞片,采用透射电镜观察并拍照。
2 结果与分析
2.1 形成层细胞恢复活动期超微结构变化
图1为湿地松形成层原始细胞恢复活动期细胞横切面超微结构。湿地松形成层原始细胞一般在3月初恢复活动,形成层细胞与木质部细胞具有明显界限,细胞层数为2~3层,3月底细胞明显液泡化,基本呈椭圆形,形成层纺锤形细胞弦向壁和径向壁厚度无明显差异,为0.46~0.66 μm(图1A、B)。形成层细胞内逐渐形成大液泡,油滴和淀粉颗粒含量明显减少,淀粉颗粒出现电子透明区(图1B)。含细胞器的细胞质被大液泡挤到细胞边缘地带,可见明显的质壁分离现象(图1C、D)。在径向壁,特别是与弦向壁相连的角隅处有部分自溶现象(图1A、D),成熟射线细胞中含有分散的小液泡,细胞质浓度明显减少,细胞内含物较丰富(图1E)。上一年形成的管胞壁角隅处也存在部分自溶现象(图1F)。从生理发育角度考虑,自溶现象主要是形成层活动提供营养物质的同时,未木质化的细胞壁柔软,细胞形状易变化,细胞壁间更好接触,增强木材内接强度,适应木材新细胞和木材形成的需要。
2.2 形成层细胞活动期超微结构变化
图2(4月18日)、图3(5月11日)、图4(6月22日)、图5(9月23日)为湿地松形成层细胞活动期不同时间的细胞超微结构。4月初形成层细胞进入活动期,整个形成层带高度液泡化,径向壁和弦向壁均变薄,且形状很不规则,径向壁和弦向壁厚度差异明显,径向壁厚度为040~0.71 μm,弦向壁厚度为0.20~0.42 μm(图2A、B)。形成层细胞中出现许多大小不一的小泡,同时质膜向内折叠形成较大凹陷(图2C)。与恢复活动期(图1C、D、E)相比,在与木质部细胞相邻的木质部射线细胞中,细胞质浓度明显降低,细胞内含物减少,但油滴含量相比其他细胞仍然较高(图2D)。5月中旬,在径切面可观察到发生径向膨大的木质部细胞,且开始出现次生壁加厚,2个正在分化的木质部细胞间存在丰富的胞间连丝,细胞质基本贴壁分布(图3A、B)。6月中旬,形成层细胞活动达到旺盛期,此时形成层带包括形成层原始细胞以及大量正在分裂的木质部和韧皮部母细胞,细胞层数明显增加,初生壁周围有丰富的颗粒状物质沉积,正在进行次生壁加厚,韧皮部母细胞弦向壁明显加厚,但整个形成层带细胞的弦向壁和径向壁仍很薄,且径向壁比弦向壁厚,径向壁厚度为1.29~2.00 μm,弦向壁厚度为0.11~0.40 μm(图4A、B、D)。形成层细胞中深色的蛋白质物质、淀粉颗粒和油滴数量迅速减少。上一年形成的射线细胞中内含物急剧减少,大量物质被消耗(图4C)。在径切面可明显观察到形成层细胞径向壁和弦向壁加厚,刚形成的细胞壁排列较疏松(图4D)。形成层带细胞层数增加的同时,未成熟木质部细胞不断发育,次生壁厚度也逐渐增加,细胞壁出现分层现象。到9月底,管胞壁的次生壁加厚基本完成,形成层细胞层数明显减少,为2~3层,径向壁和弦向壁明显增厚,弦向壁厚度为0.29~0.44 μm,径向壁厚度为0.71~1.89 μm(图5A、B)。随着木质部细胞逐渐增加,管胞壁次生壁上S2、S3层不断积累,层次结构逐渐明显(图5A、B)。

图1 湿地松形成层细胞恢复活动期(3月22日)细胞超微结构Fig.1 The ultrastructure of cambium cells during the reactivation period (March 22th) of P.elliottiiA—D为形成层带细胞的横切面,E—F为次生木质部细胞的横切面。A.形成层细胞的细胞质浓度降低,形成层带与木质部之间界限明显,细胞壁角隅处有自溶现象;B.淀粉颗粒出现电子透明区,直径开始减小,内含物减少;C.原生质体凝缩,发生显著的质壁分离现象,细胞质呈条带状分布;D.径向壁与弦向壁相连的角隅处有自溶现象(箭头所示),油滴直径减小,内含物分布不均匀;E.木质部射线细胞中含丰富的内含物和分散的小液泡;F.上一年形成的管胞分子胞间层间出现自溶现象。A—D:The cross-section of the cambium cells;E—F:The cross-section of the secondary xylem cells.A.The cytoplasmic concentration of the fusiform cells was reduced,the boundary between the cambium zone and the xylem was obvious,and there was autolysis at the corners of the cell wall.B.The starch particles appeared electron transparent zone and the diameter began to decrease,and its the content also decreased.C.The protoplasts condense and showing the significant plasmolysis,the cytoplasm was a strip distribution adherent cell wall.D.The autolysis occurred at the corner where the radial wall was connected to the tangential wall (arrow),the diameter of the oil droplets decreased and inclusions were distributed unevenly.E.Xylem ray cells contained abundant contents and scattered small vacuoles.F.The tracheid formed in the previous year occurred autolysis in the intercellular layers.
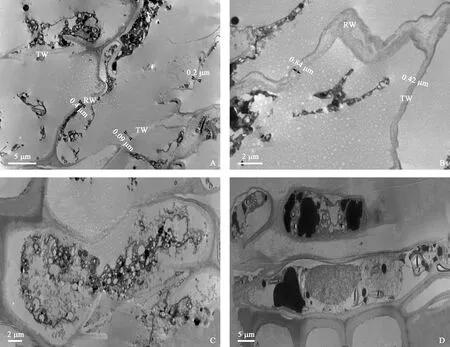
图2 湿地松形成层细胞活动期(4月18日)细胞超微结构Fig.2 The ultrastructure of cambium cells during the active period (April 18th) of P.elliottiiA.整个形成层带细胞均已高度液泡化,径向壁和弦向壁明显变薄,且形状很不规则;B.细胞弦向壁更薄,径向壁和弦向壁厚度差异明显;C.靠近木质部的形成层纺锤形细胞,细胞内含物较丰富;D.上一年形成的射线细胞中含有大油滴,细胞核位于中部,淀粉颗粒出现电子透明区。A.The entire cambial cells had been highly vacuolated,the radial wall and tangential wall were significantly thinned,and the shape of cambial cells was very irregular.B.The tangential wall of the cambium cell was thinner,and the thickness of the radial wall and the tangential wall was obviously different.C.The cambium cells near the xylem was abundant in contents.D.The ray cells formed in the previous year contained large oil droplets,the nucleus was located in the middle and the starch granules showed the electron transparent zone.

图3 湿地松形成层细胞活动期(5月11日)细胞超微结构Fig.3 The ultrastructure of cambium cells during the active period(May 11th) of P.elliottii A.紧邻次生加厚木质部细胞的形成层纺锤形细胞,箭头所示为纹孔口;B.细胞壁上有丰富的胞间连丝,细胞质贴壁分布,含有少量内含物。A.The fusiform cells near the secondary thickened xylem cells,the arrow showed the pit aperture.B.The cell wall had abundant in plasmodesmata,with cytoplasm adhering to the wall and containing a small amount of inclusions.

图4 湿地松形成层细胞活动期(6月22日)细胞超微结构Fig.4 The ultrastructure of cambium cells during the active period(June 22th) of P.elliottiiA.细胞高度液泡化,径向壁与弦向壁厚度差异显著,初生壁附近有丰富的颗粒状物质;B.靠近韧皮部的形成层细胞,弦向壁明显变厚,但径向壁仍比弦向壁厚;C.上一年形成的射线细胞中含少量内含物,细胞核呈不规则状;D.形成层细胞径向壁明显加厚。A.The fusiform cells were highly vacuolated,with significant differences between the radial wall and the tangential wall,and there were abundant granular materials near the primary wall.B.The cambium cells near the phloem cells,the tangential wall was thick,but the radial wall was still thicker than the tangential wall.C.The ray cells formed in the previous year contained a small amount of inclusions and the nucleus was irregular.D.The radial walls of cambium cells was obviously thickened.

图5 湿地松形成层细胞活动期(9月23日)细胞超微结构Fig.5 The ultrastructure of cambium cells during the active period(September 23th) of P.elliottiiA.形成层细胞仅有2~3层,径向壁和弦向壁差异显著;B.靠近形成层的木质部细胞已经完成次生壁加厚过程发育为成熟细胞,具有明显的层次结构,形成层纺锤形细胞中含有很多颗粒状物质。A.There were only 2-3 layers of cambium cells,and the radial wall and the tangential wall was significantly different.B.The xylem cells near the cambium cells had completed the secondary wall thickening and developed into mature cells with the distinct layered structure.The fusiform cells contained many granular substances.
2.3 形成层细胞休眠期超微结构变化
图6(10月26日)、图7(1月17日)为湿地松形成层细胞休眠期不同时间的细胞超微结构。10月底至次年2月底,形成层细胞处于休眠期,形成层带通常只包含1~2层细胞,形成层纺锤形细胞扁平且细胞质较稠密,弦向壁和径向壁均明显比活动期增厚,且径向壁比弦向壁厚,径向壁厚度为1.18~2.01 μm,弦向壁厚度为0.18~0.36 μm(图7A)。木射线细胞中,小液泡逐渐融合成大液泡,大液泡中含有深色的蛋白类物质和大量油滴(图6A、7B)。在相邻的射线细胞水平壁上可观察到丰富的胞间连丝,可能有利于物质运输(图6B)。在进入休眠期前,靠近形成层的木质部细胞次生壁沉积过程已经结束,除了射线细胞外,其他类型细胞的细胞质在休眠末期完全消失(图6A、6B、7B)。在整个休眠期,没有观察到有丝分裂现象,也没有新形成的细胞壁出现。

图6 湿地松形成层细胞休眠期(10月26日)细胞超微结构Fig.6 The ultrastructure of cambium cells during the dormant period (October 26th) of P.elliottiiA.木质部射线细胞中内含物较多,小液泡融合为大液泡,大部分液泡被深色蛋白质物质所填充;B.射线细胞水平壁上有丰富的胞间连丝。A.The xylem ray cells contained more inclusions,small vacuoles fused into large vacuoles,most of the vacuoles were filled with dark protein material.B.There are abundant plasmodesmata on the horizontal wall of ray cells.

图7 湿地松形成层细胞休眠期(1月17日)细胞超微结构Fig.7 The ultrastructure of cambium cells during the dormant period (January 17th) of P.elliottiiA.形成层细胞仅有1~2层厚壁细胞,形成层细胞中含很多深色物质,形成层细胞的弦向壁和径向壁都较厚,径向壁明显比弦向壁厚;B.木质部射线细胞中小液泡开始融合成大液泡,发生轻微的质壁分离现象。A.The cambium zone consisted of only 1-2 layers of thick-walled cells and contained many dark substances.The tangential walls and radial walls of the cambial cells were thicker,and the thickness of the radial walls was significantly thicker than the tangential wall.B.In the xylem ray cells,small vacuoles began to fuse into large vacuoles,and showing slight plasmolysis.
3 讨论
温带地区树木形成层活动具有明显周期性,但不同树种之间差异较大(Oladietal.,2011),同一树种生长在不同环境条件下,其形成层活动周期也不完全相同。Rao等(2001)研究发现,印度不同林地印楝(Azadirachtaindica)树维管形成层活动方式不同,其在灌木丛林中可全年处于活跃期。Singh等(2011)研究表明,印度亚热带雨林思茅松(Pinuskesiya)2月中旬新针出现后,就会重新激活休眠的形成层,12月底至次年1月底形成层活动停止。但Pumijumnong(2006)发现,泰国热带雨林思茅松3、4月处于休眠阶段,5月雨季来临时形成层开始活动直到10月,然后在干燥季节进入休眠。
在武汉,湿地松形成层细胞活动期从3月初一直持续到10月底,4月中旬进入活动期;而在原产地(美国佛罗里达),湿地松生长季从2月底一直延续到12月(Harleyetal.,2012),这可能与光照、温度和降雨量有关,冬季日照缩短、低温和少量降雨导致形成层细胞提早进入休眠期(Singhetal.,2011;Begumetal.,2012;2015;Carreretal.,2017)。湿地松在6月中旬左右达到活动旺盛期,形成层带细胞层数明显增加,形成层带细胞包括形成层原始细胞以及大量正在分裂的木质部和韧皮部母细胞,木质部细胞开始进行初生壁加厚和次生壁沉积,进一步证明7月左右湿地松开始产生晚材的结论(Harleyetal.,2012)。湿地松形成层带旺盛期可达到的最大细胞层数明显少于阔叶树种(Minetal.,2016;Liuetal.,2014;Chenetal.,2010),进一步解释了针叶树年生长缓慢的原因。10月底至次年2月湿地松处于休眠期,通常只包含1~2层形成层带细胞,细胞形状扁平,细胞弦向壁和径向壁均明显比活动期增厚,且径向壁和弦向壁厚度差异更加显著,细胞内含物少量增加。形成层带细胞经历一段时间休眠,也正好可以解释其明显的年轮界限。
湿地松形成层细胞恢复活动期,径向壁特别是与弦向壁连接的角隅处出现部分自溶现象,该现象可能在一定程度上破坏了细胞壁结构,提高了细胞壁的弹性和可塑性,有利于活动期形成层细胞的径向生长(Event,2006)。Wu等(2016)发现,杉木在休眠期,形成层径向壁比弦向壁含有更多果胶和少量木质素,果胶积累促进了细胞壁的扩张和形成层的活化。湿地松形成层活动期时,细胞形状变得不规则,整个形成层带细胞高度液泡化,淀粉颗粒出现电子透明区,油滴直径减小。纺锤形细胞壁尤其是径向壁变薄是形成层恢复活动的开始,且整个活动期和休眠期内,形成层细胞径向壁明显比弦向壁厚。这些超微结构特征均与温带地区其他阔叶树种类似,如柚木(Tectonagrandis)(Raoetal.,2010)、山毛榉(Fagussylvatica)(Prislanetal.,2011)、毛白杨(Populustomentosa)(Yinetal.,2002;Chenetal.,2010;Arendetal.,2003)等;唯一不同的是,湿地松形成层在恢复活动期径向壁和弦向壁厚度差异不是很明显。无论是纺锤形细胞还是射线细胞,均可观察到丰富的胞间连丝,可能有利于细胞间的物质运输和细胞壁的建成。根据细胞超微结构变化,比以往主要通过光学显微镜(Harleyetal.,2012)能更清楚、更准确地观察到形成层休眠期向活动期转变的分化特点以及整个活动期内形成层细胞的分裂分化。
湿地松在整个生长季均会出现松针脱落现象,休眠期间也有新的松针产生,表明松针的产生和脱落与形成层细胞的休眠和激活无明显关联。研究还发现湿地松次生木质部中成熟的射线细胞一直含有较多内含物,其原生质体没有出现衰退现象,说明次生木质部中的射线薄壁细胞多年内仍保持较高生理活性,以备于形成层细胞受伤后射线薄壁细胞转化为形成层细胞进行新的分裂分生活动。
4 结论
1) 湿地松形成层细胞层数基本为1~2层,其年活动期内具有明显的休眠期和活动期变化。2月底至3月底为形成层细胞恢复活动期,径向壁变薄是形成层恢复活动的主要特征。部分自溶现象发生在形成层细胞径向壁,尤其在细胞壁连接处。打破休眠期的标志是形成层纺锤形细胞的出现,纺锤形细胞径向壁和弦向壁厚度差异不显著,呈椭圆形或近圆形。形成层纺锤形细胞出现少许质壁分离现象,淀粉颗粒出现明显电子透明区。3月底细胞明显液泡化,基本呈椭圆形,形成层纺锤形细胞弦向壁和径向壁厚度无明显差异。
2) 4月初至10月底为湿地松形成层细胞活动期,是湿地松木材形成的主要阶段。活动期内,整个形成层带细胞高度液泡化,细胞形状变得不规则,径向壁和弦向壁均变薄,弦向壁更薄且变得不明显,油滴和淀粉颗粒急剧减少。
3) 6月中旬为湿地松形成层细胞活动旺盛期。旺盛期形成层带细胞层数明显增加,次生壁开始加厚,原生质体基本瓦解并消失。
4) 10月底至次年2月底为湿地松形成层细胞休眠期。休眠期开始后,形成层带细胞层数明显减少,只含有1~2层扁平的厚壁纺锤形细胞,其径向壁和弦向壁明显增厚,细胞内含物开始积累。整个活动期内形成层细胞径向壁明显比弦向壁厚,休眠期这种差异更明显。
猜你喜欢
智慧农业导刊(2022年22期)2022-11-17
农业与技术(2021年6期)2021-12-12
西藏人文地理(2020年4期)2020-11-19
农业工程技术·综合版(2020年1期)2020-04-23
林业与生态(2020年2期)2020-04-16
中国农业科技导报(2020年1期)2020-03-15
科学咨询(2020年36期)2020-01-06
中国真菌学杂志(2019年1期)2019-01-23
湖南林业科技(2017年4期)2017-11-24
江苏农业科学(2016年5期)2016-07-23
