
可见-近红外光谱结合机器视觉动态检测花生黄曲霉毒素B1污染
2020-12-04 13:25蒋雪松何学明周宏平刘兴泉
光谱学与光谱分析 2020年12期
严 晨,蒋雪松*, 沈 飞, 何学明,方 勇,刘 琴,周宏平, 刘兴泉
1. 南京林业大学机械电子工程学院,江苏 南京 210037 2. 南京财经大学食品科学与工程学院,江苏 南京 210023 3. 浙江农林大学农业与食品科学学院, 浙江 杭州 311300
引 言
花生作为一种重要的油料作物,在世界各地均有大规模种植。 在生长和储藏过程中,花生易受温度、湿度的影响而发生霉变。 感染的有害霉菌会产生一种毒性很强的次生代谢产物——黄曲霉毒素[1]。 黄曲霉毒素是一种高毒性和致癌性的化合物,会严重影响花生的品质和安全。 其中以黄曲霉毒素B1(Aflatoxin B1,AFB1)毒性最大,对动物和人体有较高的健康威胁,被国际癌症研究机构划定为Ⅰ类致癌物[2]。 依照《GB2761 2017食品安全国家标准 食品中真菌毒素限量》, 我国对花生及其制品中黄曲霉毒素B1限量指标为20 μg·kg-1 [3]。
传统的黄曲霉毒素检测方法如薄层色谱法,高效液相色谱法以及酶联免疫吸附法等[4]虽然精确,但操作较复杂、成本较高且不适合实时控制,不能满足花生储运及交易时快速检测要求。 因此需要一种快速、便捷且有效检出花生中黄曲霉毒素B1超标的方法。 由于近红外光谱技术以及机器视觉技术的无损、快速等特性,国内外已有学者将该技术应用于粮食安全的检测中[5-8]。 从大多研究结果来看,近红外光谱技术和图像技术有能力对霉菌侵染程度准确判别和预测; 但是大多研究以不同浓度毒素浸泡的样品作为研究对象,且以静态检测为主,动态检测自然霉变花生中黄曲霉毒素B1的报道较为罕见。
本研究以自然霉变的花生籽粒为对象,动态获取其可见-近红外光谱和图像信息,结合化学计量法建立花生中黄曲霉毒素B1超标与否的检测方法,实现AFB1超标花生的动态筛选,为最终实现花生质量安全的在线监测提供技术参考。
1 实验部分
1.1 材料
花生籽粒通过市场选购,品种为大白沙,产地山东青岛,挑选表面没有破损、发霉及发芽且大小相似的花生籽粒。
1.2 方法
利用分析天平称取150份,每份100 g的花生,将150份花生样品放置在带有透气孔的方形容器中。 转移至人工气候箱中,在28 ℃,85%相对湿度(RH)的条件下储藏8 d。 以样品放入人工气候箱时刻作为第0天,选取储藏节点0, 4, 6, 7和8 d取样,每次相同时刻取出30份用于分析,将第0 d花生样品作为对照组,并将样品放置于密封袋中于-4 ℃冰柜冷藏,待后续分析。
1.3 花生样品可见-近红外光谱和机器视觉动态采集
图1为可见-近红外光谱与机器视觉动态检测系统,该系统由可见-近红外光谱仪(德国Zeiss公司MCS 600型)、OMK500-H/NIR漫反射探头(德国Zeiss公司)、MV-
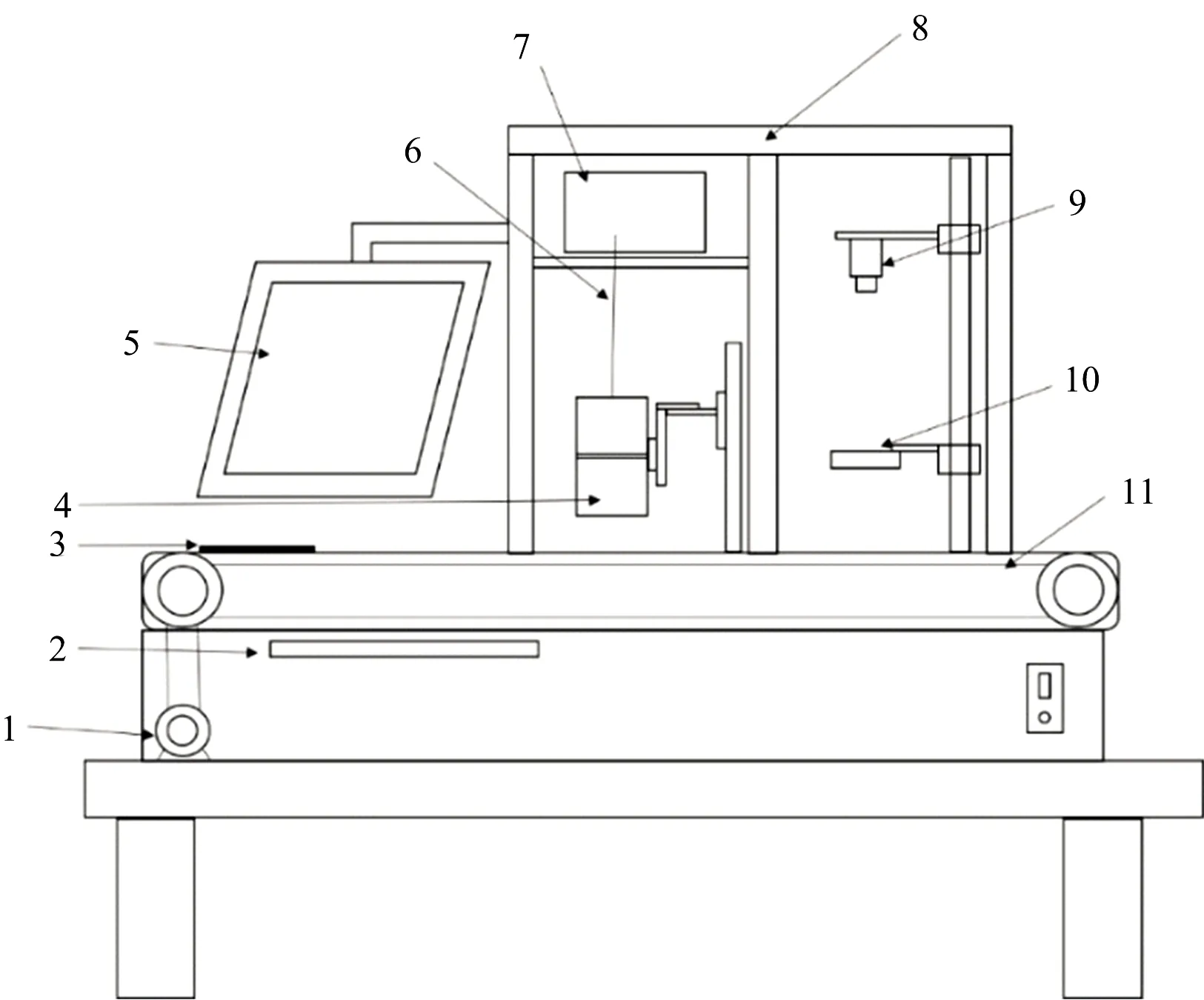
图1 可见-近红外光谱和机器视觉检测系统
1: 驱动电机; 2: 键鼠; 3: 载物盘; 4: 漫反射探头; 5: 显示器; 6: 光纤; 7: 光谱仪; 8: 暗箱; 9: 数字相机; 10: 环形光源; 11: 传送带
Fig.1 Visible/near infrared spectroscopy and
machinevisioninspectionsystem
1: Drive motor; 2: Keyboard; 3: Slide plate; 4: Diffuse reflection probe; 5: displayer; 6: Optical fiber; 7: Spectrograph; 8: Camera obscura; 9: Digital camera; 10: Ring light source; 11: Conveyor
EM120C/M型相机(陕西维视图像公司)、LED环形光源、暗箱、驱动电机、传送带、电脑显示器、光纤以及载物盘等部分构成。
采集前将冷藏(-4 ℃)的花生样品置于室温下(23±1) ℃下2 h以上,直至样品平衡到室温。 为使光谱系统稳定,将红外光谱仪预热30 min。 首先对光谱进行黑白校正,之后将花生样品放置于直径90 mm高15 mm的石英培养皿中,将培养皿放置于载物盘上,样品上表面距探头约20 mm,样品表面光斑约3.14 cm2。 样品随传送带以0.15 m·s-1速度运动至漫反射探头下方时,采集样品光谱(560~1 700 nm),积分时间设置为100 ms; 当样品随传送带以0.15 m·s-1速度运动至数字相机正下方时,采集样品图像,此时样品上表面距数字相机镜头约310 mm,图片分辨率为1 280×960像素,以JPEG格式存储; 每个样品重复装样并采集光谱和图像各三次,取平均分析。
显然,只有当扫描到待充电设备的两端建立连接,发射端读取接收端反馈并写入当前待充电设备的型号、电量等信息后开始充电且充电判断完成,才为一次一对一的无线充电成功。
1.4 样品AFB1含量
参照GB 5009.22—2016《食品中黄曲霉毒素B族和G族的测定》中酶联免疫吸附筛查法,在工程师的指导下,利用黄曲霉毒素B1ELISA试剂盒(北京华安麦科生物技术有限公司)测量样品AFB1含量。
样品前处理: 粉碎并取5.0 g有代表性的花生样本置入100 mL具塞三角瓶中,加入40%甲醇25 mL与其混合; 于振荡器上剧烈振荡10 min,转速为200 r·min-1; 取液体于4 000 r·min-1离心5 min; 取上清液1 mL,加入40%甲醇2 mL,振荡10 s; 取液体1 mL,再加入4 mL去离子水; 振荡5 s,取50 μL进行分析。
操作步骤: 加入标准品或样本50 μL到对应的微孔中,加入AFB1酶标物50 μL·孔-1,再加入AFB1抗试剂50 μL·孔-1,轻轻振荡混匀,用盖板膜盖板后置25 ℃避光环境中反应30 min; 小心揭开盖板膜,将孔内液体甩干,用洗涤液300 μL·孔-1; 充分洗涤4~5次,每次间隔10 s,用吸水纸拍干; 加入底物液A液50 μL·孔-1,再加底物液B液50 μL·孔-1,轻轻振荡混匀,用盖板膜盖板后置25 ℃避光环境反应15 min。 加入终止液50 μL·孔-1,轻轻振荡混匀,设定酶标仪于双波长450/630 nm处,测定每孔OD值。
利用试剂盒配套的专业分析软件计算每个样本的AFB1含量。 结果显示,随储藏时间的增加,4 d已有5份样品AFB1超标,6 d及之后的大部分花生样品AFB1含量开始逐渐超过20 μg·kg-1,超标率为58%。
1.5 数据分析
采用MATLAB 2015b软件(美国Matheorks公司)和Unscrambler X10.4软件(Camo Analytics公司)对花生样品的光谱和图像信息进行处理和建模。 为消除基线漂移、噪声等带来的误差,采用多元散射校正(multiplicative scatter correction,MSC),Savitzky-Golay(S-G)平滑和标准正态变量校正(standaedized normal variate,SNV)等方法对光谱进行预处理,并基于主成分分析(principal component analysis,PCA)权重系数优选特征波长作为后续模型输入。 利用灰度化、形态学运算等方法对图像信息进行处理,剔除无效信息,以图像的RGB(红色、绿色、蓝色)和HIS(色调、饱和度、亮度)的各分量均值以及方差作为图像特征参数。 将获取的光谱特征波长和图像特征参数融合作为样品的总特征信息。 在建立模型前,先运用主成分分析提取主要数据并进行聚类分析; 再通过线性判别分析(linear discriminant analysis,LDA)与支持向量机(support vector machine,SVM)建立花生黄曲霉毒素B1侵染程度的判别模型。 模型建立时,将150份样品按2∶1随机分为建模集和预测集。 按照国家标准,将高于20 μg·kg-1的样本作为超标样本,低于20 μg·kg-1的样本作为未超标样本。 在模型中将未超标样本记为类N,超标样本记为类Y。
2 结果与讨论
2.1 可见-近红外光谱分析
利用可见-近红外系统动态采集花生样品光谱,选取光谱范围600~1 600 nm进行分析。 图2为五份不同黄曲霉毒素B1含量的样品原始光谱和二阶导数光谱图。 原始吸收光谱图(图2a)与其他研究报道的相符,不同AFB1含量的样品光谱整体相似,在962,1 150以及1 415 nm三个波长附近有吸收峰。 有相关研究表明,这分别与蛋白质中N—H的二级振动、淀粉或脂质中C—H的二级振动、水中O—H的一级基频振动等有关[9]。 为了进一步分析,对光谱进行二阶导数处理以消除大量噪音并放大部分波段。 图2(b)观察到在920,1 080,1 180和1 320 nm等波峰附近,随着霉变程度的加深,AFB1侵染水平的增加,吸光度在逐渐降低,这种现象在一些小麦和玉米的真菌感染研究中也有所描述[10]。 真菌污染过程中,花生中的成分发生改变,如脂肪作为主要碳源被消耗,并被分解为游离脂肪酸和甘油脂,而脂肪对于1 180 nm处的近红外光吸收较为敏感[11]。 此外,在近红外波长范围内,光谱差异总体上是由散射现象决定的,可能真菌污染会破坏花生籽粒的细胞壁并使籽粒变得多孔,从而增加近红外波长范围内的散射光,导致吸光度有所降低[12]。
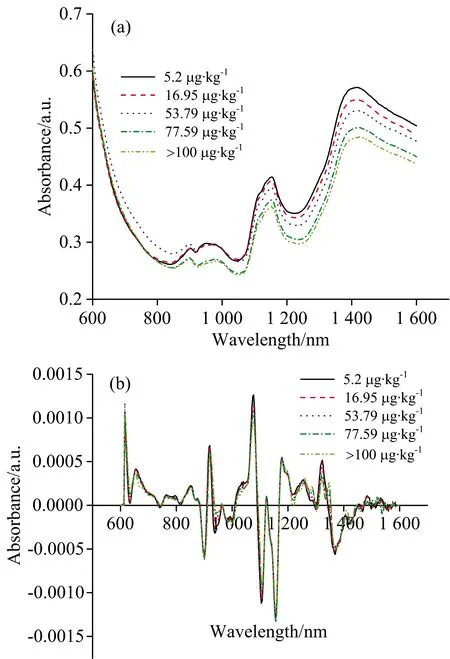
图2 五种不同AFB1含量的花生样品原始(a)与二阶导数(b)光谱图
2.2 花生图像信息分析
利用MATLAB 2015b对花生样品图像进行处理。 首先对图像灰度化处理,将彩色图像转化为灰度图像,以减少计算量,加快处理速度。 再将灰度图像二值化处理,利用最大类间方差法将图像分割为目标和背景两部分。 最后提取图像的RGB和HIS颜色模型中的各颜色分量均值和方差作为图像颜色特征参数。 从图3和图4中观察发现,随着毒素含量的增加,花生样品RGB值总体呈下降趋势,表面颜色变化较为明显,低于20 μg·kg-1的花生样品表面干净有光泽,20~50 μg·kg-1样品颜色则相对暗淡,已有少量菌丝附着。 当毒素含量超过100 μg·kg-1时,花生样品表面暗淡并且有大量菌丝覆盖,霉变严重。

图3 不同AFB1含量的花生样品原始图像和对应灰度图
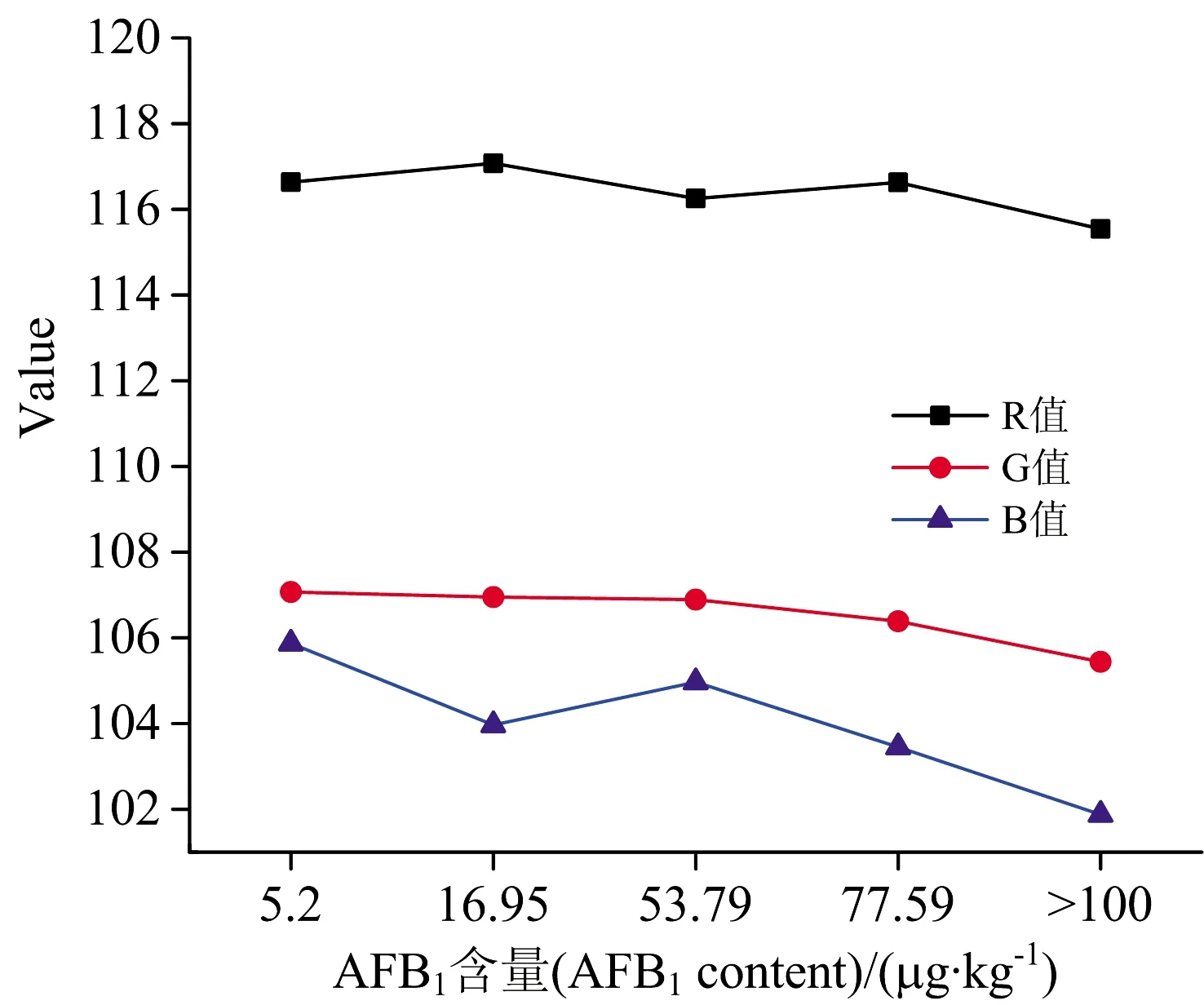
图4 不同AFB1含量的花生样品RGB值变化图Fig.4 RGB value of peanut samples contaminatedwith different levels of AFB1
2.3 主成分分析
为适应动态检测的需要,需提取特征波长作为模型输入以提高模型运算速度。 PCA权重系数α可用来选取特征波长,权重系数曲线中的局部最高峰和最低峰都可认为是一个特征波长[13]。 在图5中,PC1的加载曲线较为平滑,最佳波长不明显,而PC2和PC3有较为明显的峰值,选择630,1 067,1 150,1 227,1 390和1 415 nm作为最佳波长。

图5 花生样品的前3主成分权重系数图Fig.5 Weighting coefficients for PC1—PC3 of peanut samples
利用PCA根据毒素水平可以探寻样品分组的可能性。 图6是花生样品的光谱、图像和数据融合信息的PCA得分图,前两个主成分贡献之和在71%~99%之间,体现了原始数据中的大多数变化。 从图6(a)和(b)中可以观察出虽然有小部分样品重叠,但是整体上仍存在一定的聚类趋势。 这表明,随着霉变程度的加深,毒素含量的变化,花生的可见-近红外光谱也随之发生一定的改变,为后续定性判别提供了基础。 而图6(c)和6(d)中观察到第一第二聚类趋势不明显,大部分样品重叠,无法实现超标样品和未超标样品的区分。 这可能由于毒素的含量较低以及非产毒菌落的影响,因此需要进一步建立判别分析模型来识别超标和未超标样本。
2.4 花生样品AFB1超标与否的定性判别分析
PCA虽然可探寻出不同样品之间的聚类趋势,但是并不能表达出具体的分类结果。 因此采用LDA和SVM判别分析方法分别基于全谱段、最佳波长、图像和数据融合建立花生AFB1超标与否的定性分析模型。

图6 花生样品第一、第二主成分得分图(a): 可见-近红外全谱段; (b): 特征波长; (c): 图像; (d): 特征波长与图像融合特征参数Fig.6 Score plot of PC1 and PC2 of peanut samples(a): Visible/near-infrared full-band; (b): Characteristic wavelength; (c): Image; (d): Spectral and image fusion characteristic parameters
通过对光谱数据使用不同预处理方法开发了多种LDA判别模型; 通过对全谱段光谱平滑去噪以及消除散射处理,经S-G和MSC处理的全谱段光谱分类准确性相对原始光谱略有提升,预测集提升至92%,而原始光谱的特征波长模型分类准确性优于经处理的光谱。 表1和表2分别是根据样品全谱段、特征波长、图像以及特征波长和图像融合信息建立的LDA和SVM模型结果。 可以看出全谱段下,SVM模型效果低于LDA模型,最佳建模集准确率分别为89%和94%,最佳预测集准确率分别为82%和92%; Tao[14]等利用可见-近红外光谱对侵染6个AFB1浓度梯度的花生进行鉴别研究,以20 μg·kg-1作为分类阈值,基于全谱段建立的PLS-DA和LS-SVM模型准确率分别达到88.6%和90%,与本研究结果类似。
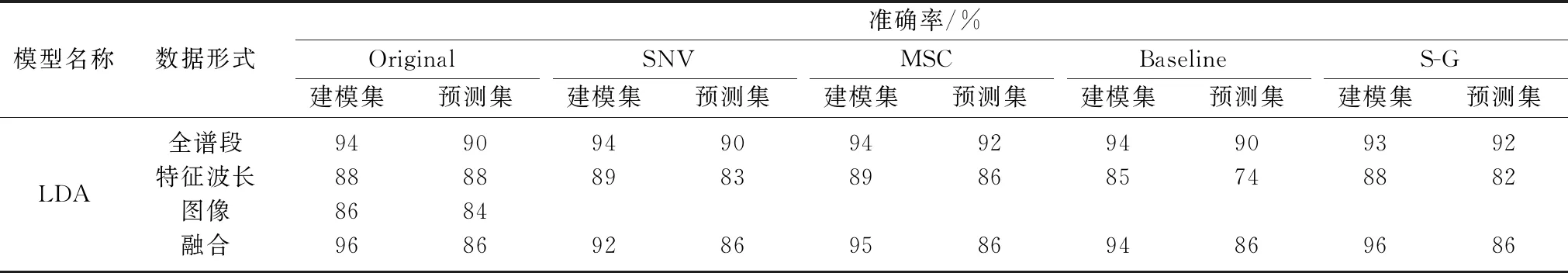
表1 基于不同数据形式的花生样品LDA分类模型结果Table 1 Results of LDA classification model of peanut samples based on different data forms

表2 基于不同数据形式的花生样品SVM分类模型结果Table 2 Results of SVM classification model of peanut samples based on different data forms
为了满足在线应用的需要,探讨了基于特征波长建立的LDA和SVM模型。 从表1和表2观察到以特征波长作为输入的LDA和SVM模型效果相对全谱段有所降低,最优模型为88%,这可能是由于提取特征波长时将一些相关信息遗失,但在一定程度上提高了运算速度,降低了模型的复杂程度。 随着储藏时间的延长,有害霉菌逐渐繁殖并排放毒素,导致花生样品的外观特征产生变化,如花生图像颜色逐渐灰暗,亮度降低,籽粒多孔等,使得图像特征参数产生一定的变化。 为了进一步提高模型的准确性,引入花生样品图像颜色特征参数建立判别模型。 不同核函数的SVM模型分类准确率不同,以径向基函数核(RBF kernel)为核函数的Nu-SVM模型效果最优,预测集准确率为90%。 LDA在利用光谱信息建立判别模型中表现相对较优,而非线性模型SVM则更适合于基于图像信息建模。
黄曲霉毒素B1的侵染会引起花生内部品质如蛋白质和脂肪的变化,这些变化能引起近红外光谱的改变,从而能够间接指示花生中毒素的含量。 将来自图像的外观特征和近红外光谱的内部化学键信息进行融合处理,综合两者的优势,能在一定程度上提升分类模型的效果。 在此基础上分别建立LDA和SVM融合模型。 表1观察到LDA融合模型相对于单独应用特征波长和图像信息时的分类准确度均有所提升,其中以Baseline-LDA融合模型效果提升最大,预测集分类准确度提高12%。 表2的6种SVM融合模型中,除Sigmoid内核的SVM融合模型效果略微下降,其余模型均有不同程度提升。 纵观所有融合模型,以RBF为核函数的C-SVM融合模型效果最优,分类准确度达到92%,相较于利用特征波长建模效果提升18%。 结果表明,可见-近红外光谱技术和机器视觉技术获得花生样品的内部和表面信息并结合化学计量法,能够实现花生样品黄曲霉毒素B1的动态判别,且表现出可观的精度。
3 结 论
以自然霉变花生为研究对象,对可见-近红外光谱结合机器视觉技术动态分类黄曲霉毒素B1超标和未超标花生的可能性进行了探索。 结果表明不同AFB1侵染程度的花生光谱和图像呈现一定的差异; 进一步采用LDA和SVM建立的花生黄曲霉毒素B1超标判别模型拥有良好的准确性,并且通过优选特征波长建模能够简化模型,提高运行速度,有利于动态应用。 利用近红外信息结合表面图像信息建立LDA和SVM判别模型,相较单独利用特征波长建立模型准确率有所提高,模型最佳识别率提升至92%。 因此利用可见-近红外光谱结合机器视觉技术,实现花生AFB1含量超标与否的动态判别具有一定可行性。
猜你喜欢
考试与评价·高二版(2021年3期)2021-09-10
现代畜牧科技(2021年2期)2021-03-19
数学物理学报(2020年5期)2020-11-26
天然产物研究与开发(2018年9期)2018-10-08
天然产物研究与开发(2018年8期)2018-09-10
中南大学学报(自然科学版)(2016年2期)2017-01-19
中国照明(2016年4期)2016-05-17
公民与法治(2016年14期)2016-05-17
中国当代医药(2015年26期)2015-03-01
云南中医学院学报(2014年5期)2014-07-31
