
基于傅里叶光谱技术对Pb胁迫下斜生栅藻物质成分变化的研究
2020-12-04 13:25刘群伟邵咏妮陈小婉蒋林华
光谱学与光谱分析 2020年12期
刘群伟,邵咏妮,陈小婉,蒋林华
上海理工大学上海市现代光学系统重点实验室,光电信息与计算机工程学院,上海 200093
引 言
重金属是自然界中危害较重的一类污染物,一般指密度≥4 g·cm-3的金属。 绝大多数的重金属具有致癌性,会破坏人体器官,对人体造成极大的危害。 在重金属中,铅(Pb)是三种最常见的重金属之一。 重金属危害的持久性、不易降解以及随着食物链的富集危害性逐渐增强的特点,使得我们不得不重视重金属污染的问题。 藻类植物作为水生生态系统的初级生产者,最先受到重金属的影响。 有研究表明Pb对氮和硫具有很高的亲和力,Pb离子可以替代微藻细胞内化合物分子中的其他金属离子。 因此,Pb可能会改变代谢过程中的酶或其他成分,从而最终影响细胞的分子组成。 另外,Pb可以置换金属离子,如光合机构中的Mg2+,Fe2+,Ca2+或与光合作用中的酶相互作用,导致该过程的抑制,最终致使初级生产力下降,间接改变细胞内部物质成分。
研究铅对藻类的影响,为风险评估、水质管理、金属生物修复和生物燃料生产提供了重要的数据。 但是,大多数用于这些评估的方法都是耗时和破坏性的,而且需要相对较多的细胞。 此外,评估还涉及繁琐的操作,如细胞的采集和破坏以及生物化学提取,这可能会带来更多随机和系统的误差。 光谱技术在微藻细胞方面的应用弥补了以上的不足,其中拉曼、近红外技术等在该领域已经取得了不少的成果。 如,Wu等[1]通过单细胞激光拉曼光谱技术实现对微藻体内油脂的直接定量分析。 Collins等[2]采用共焦拉曼显微镜技术观察雨生红球藻在不同生长状态下细胞内叶绿素、类胡萝卜素以及虾青素的组成变化。 此外,Brown等[3]采用近红外反射光谱技术对微藻体内的脂质进行快速检测。 Wagner等[4]利用中红外光谱技术对微藻体内蛋白质、脂质以及碳水化合物的含量进行了定量检测。 尽管拉曼、红外光谱技术在微藻物质成分研究方面取得了一定的进展,但仍然存在一些问题,如拉曼信号灵敏度低,易受到荧光干扰[5]; 而近红外光谱的信噪比低,测量灵敏度较低[6]。
在基于光谱技术的微藻成分检测方面,拉曼技术是通过谱线的强度反映分子内部的振动情况,主要用于鉴定分子中存在的官能团。 太赫兹技术则可以通过检测分子间和骨架间的振动直接反映分子的结构[7]。 与传统光谱检测相比,太赫兹具有较高的分辨率,更为重要的是许多生物大分子的振动频率处在太赫兹波段,诸多研究已经实现通过太赫兹检测多肽、DNA、蛋白质等生物大分子[7]。 傅里叶变换红外光谱中引入原子的振动和分子的化学键,产生包含不同大分子特征带的吸收谱。 每个分子都有自己的光谱特征对波段位置、宽度和强度的分析提供了有关细胞组成和生理状态的信息。 然而,该技术较少应用于包括Pb在内的重金属对藻类物质成分的影响研究,并且在使用有限的单变量比较的光谱分析方面进行的研究很少[8]。 因此太赫兹、远红外光谱技术在研究微藻体内大分子物质变化上存在一定的优势。
本研究中我们采用太赫兹、远红外光谱技术对Pb胁迫下斜生栅藻体内淀粉、类胡萝卜素含量变化进行研究。 采用傅里叶红外光谱仪采集微藻的光谱信号,同时结合主成分分析(PCA)方法,对Pb胁迫不同时间点的光谱数据进行聚类分析。 同时比较了以重金属浓度与时间为变量的条件下,斜生栅藻细胞内淀粉、类胡萝卜素含量变化趋势。 探究了傅里叶光谱技术的微藻细胞内生物大分子检测的可行性。
1 实验部分
1.1 材料
仪器与仪器设备: 紫外分光光度计(uv1902pc)、真空傅里叶变换红外光谱仪(BRUKER Vertex 70v/80v)、高速冷冻离心机(GL-20G-Ⅱ)。 斜生栅藻(FACHB-12)购于中科院武汉水生生物研究所、BG11培养基、调节pH至7.4的去离子水1 L、标准硝酸铅溶液、电热鼓风干燥箱(BGZ-30)。
1.2 方法
1.2.1 藻的培养
使用BG11培养基将斜生栅藻以1∶3的比例扩大培养。 无菌条件下,在洗净且经压力蒸汽灭菌器(XFS-280MB)高温高压灭菌的2 000 mL的三角锥形瓶中,按照3.4 g·L-1的比例加入BG11培养基,接入斜生栅藻藻种,摇匀,置于光照培养箱中,培养箱中光照强度在4 000 Lux,光周期为12L∶12D。 每日固定时间点摇匀锥形瓶4次,通气培养, CO2浓度为2%, 培养温度为(25±1) ℃,pH值为7.0。
1.2.2 实验设计
取处于生长稳定期的斜生栅藻,使用血球计数板(北京索莱宝科技有限公司)获取微藻生长稳定期的生长状态,初始藻密度为3.6×106个·mL-1。 实验使用8个洗净且经压力蒸汽灭菌器(XFS-280MB)高温高压灭菌的2 000 mL的三角锥形瓶,设置三个实验组A,B和C,一个对照组CK,另外每组设置两个平行组,向实验组和对照组中加入相同密度的斜生栅藻以及相同的标准BG11培养基。 其中实验组A,B和C中分别加入0.5,5和20 mg·L-1的Pb(NO3)2溶液,对照组CK不做处理,将实验组和对照组置于光照培养箱中,培养箱中光照强度在4 000 Lux,光周期为12L∶12D。 每日固定时间点摇匀锥形瓶4次,通气培养, CO2浓度为2%, 培养温度为(25±1) ℃,pH值为7.0。
1.2.3 斜生栅藻细胞生长的测定
根据斜生栅藻的光学特性,参照文献[8], 用UV-分光光度计(uv1902pc)分别测定斜生栅藻650 nm处的吸光度(D650)。 然后再利用显微镜计数藻细胞,得出藻细胞和吸光度之间的线性关系(图1)。 根据该关系可在后续试验中利用藻液吸光度来表示斜生栅藻的生长情况。

图1 藻细胞密度与吸光度(D650)之间的线性关系Fig.1 Linear relationship between algal cell densityand absorbance (D650)
1.2.4 红外光谱的采集与处理
每间隔3 h分别取各实验组A,B,C和CK对照组摇匀后的60 mL样品,使用高速冷冻离心机(GL-20G-Ⅱ),在环境温度下以4 000 r·min-1的转速离心5 min收获30 mL细胞悬浮液,再用30 mL蒸馏水将沉淀洗涤、离心,此操作重复两次。 然后将浓缩至1 mL的藻类溶液加入到直径为2 cm的聚乙烯模具中,并使用电热鼓风干燥箱(BGZ-30)在40 ℃下干燥2.5 h,用于生物组成测定。 此时,将样品制成厚度为20 μm的薄膜。
采集光谱时,为了消除大气水蒸气的吸收贡献,在真空腔内以4 cm-1(0.12 THz)的分辨率采集THz光谱,并且每次测量是64次样品扫描和64次背景扫描的平均值,使用Bruker 66v/s傅里叶变换光谱仪在1~20 THz的范围内。 实验每3h进行一次,在每个样品的不同点收集8组数据。
1.2.5 数据处理与统计分析
所采集的试验数据使用Excel2018,The Unscrambler X和origin2018数据分析软件进行数据统计和相关性分析。
2 结果与讨论
2.1 不同浓度的Pb2+处理对斜生栅藻生长的影响
图2为使用UV-分光光度计(uv1902pc)在不同时间段采集实验组A,B,C与对照组CK中斜生栅藻的650 nm处的吸光度(D650)值,直接反应了各个时间点斜生栅藻的生长状态。 主要表现在低质量浓度生长较快,高质量浓度使斜生栅藻生长减慢。 在胁迫的24 h内(图2),从整体上看CK对照组生长最快,0.5和5 mg·L-1低质量浓度胁迫的斜生栅藻生长状况区别不大。 说明低质量浓度的Pb离子对斜生栅藻的生长抑制程度比较小。 而在20 mg·L-1质量浓度的Pb离子胁迫的情况下,相对于对照组斜生栅藻生长状况是抑制程度较大的。 斜生栅藻在不同重金属离子影响下生长呈现差异性,很大程度上是由于在高质量浓度下,重金属在细胞膜上反应,影响细胞的分裂与渗透性。 在细胞内部,重金属离子替换活性金属或键合巯基、氨基和羰基基团而使必需酶失去活性,从而制约了细胞的生长代谢[9]。
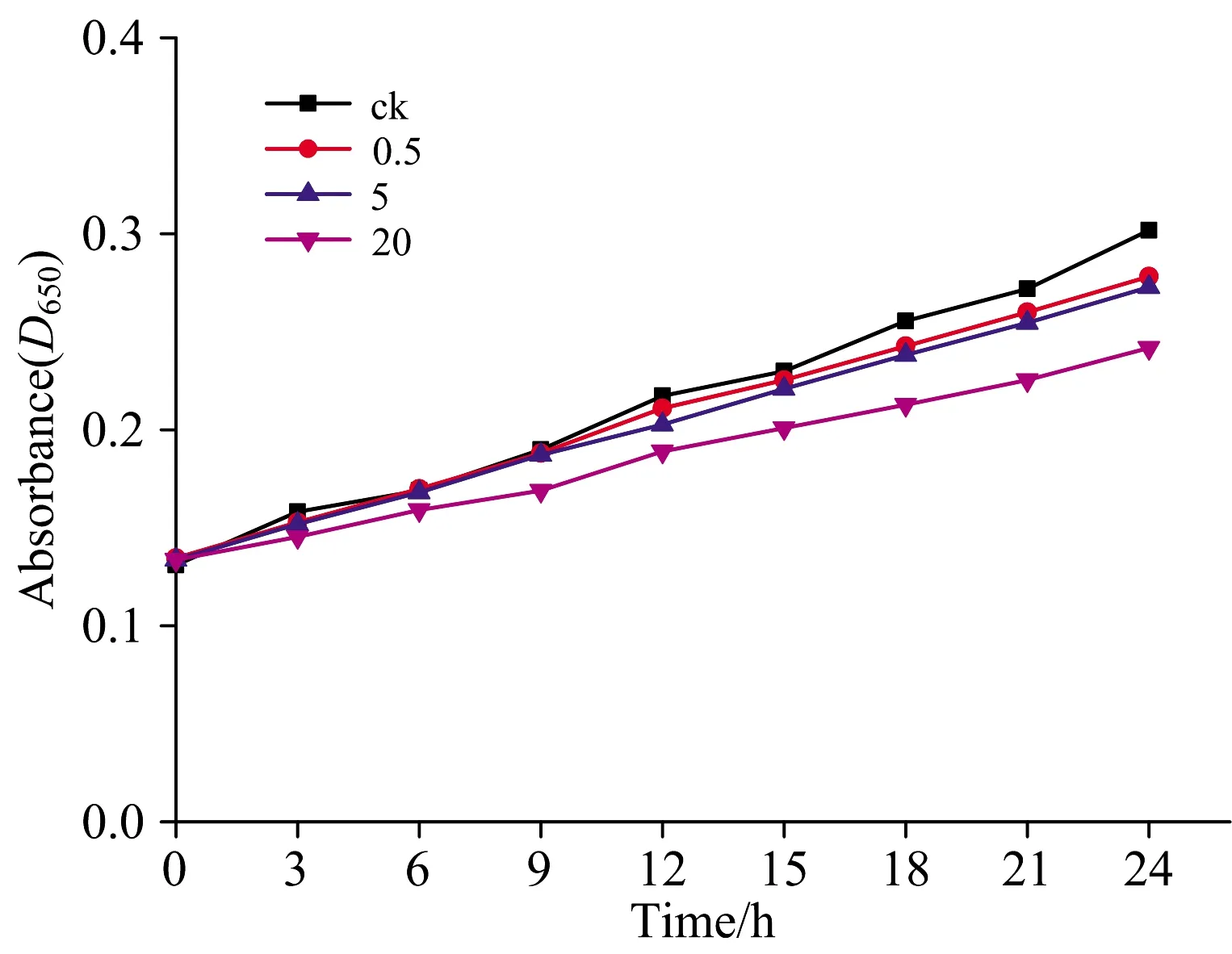
图2 重金属离子对栅藻生长的影响Fig.2 Effect of heavy metal ions on the growth of Scenedesmus
2.2 斜生栅藻的太赫兹光谱的光学特性
2.2.1 斜生栅藻在不同浓度Pb2+胁迫下同一时间点淀粉与类胡萝卜素的含量变化
光谱可以反映生物化学成分的变化,对于不同胁迫条件下的斜生栅藻,微藻内碳水化合物和类胡萝卜素含量发生变化时所对应的傅里叶光谱一般呈现出不同的趋势。 在图3显示1~20 THz的斜生栅藻几种大分子物质的吸收峰位置中,主要有碳水化合物和类胡萝卜素,其类别可通过振动光谱法鉴定。 以下是它们在光谱的太赫兹和远红外部分的不同频率区域的吸光度: 9.1,10.5和17.3 THz用于碳水化合物(淀粉)[8, 10],16.7 THz用于类胡萝卜素(β-胡萝卜素)[11-12]。

图3 Pb胁迫斜生栅藻24 h时实验组与对照组的对比
如图3所示,对照组为不含重金属的BG-11培养基,另外还有三个重金属浓度的实验组: 0.5,5和20 mg·L-1。 培养周期为24h时,每胁迫三小时检测一次FTIR,然后构建坐标系,将淀粉的特征峰(9.1,10.5和17.3 THz)与类胡萝卜素的特征峰(16.7 THz)所对应的峰值作为y值,时间点作为x轴绘制二维坐标图4(a,b,c和d)。 由图可知,在胁迫12 h内,各浓度胁迫下的淀粉含量变化程度差异不大,随着胁迫时间增加至24 h,实验组中斜生栅藻细胞内的淀粉与类胡萝卜素在不断的积累,20 mg·L-1胁迫下的斜生栅藻累积的淀粉与类胡萝卜素始终多于0.5和5 mg·L-1的实验组,另外0.5和5 mg·L-1实验组之间淀粉与类胡萝卜素含量变化程度差异不大。
实验表明,Pb胁迫下实验组A,B和C中类胡萝卜素和碳水化合物增加,Pb离子对碳水化合物和类胡萝卜素的累积有促进作用。 并且A,B和C实验组中类胡萝卜素和碳水化合物的累积关系也与其胁迫Pb离子浓度存在正相关性。 这与解毒机制有关,光合作用的变化可能是铅直接抑制光合机制引起的,或间接通过增加活性氧ROS产生的,或两者结合而成。 大分子组分的变化与铅对光合效率的抑制是一致的,因为当光合作用受到损害时,细胞优先合成,与蛋白质相比,碳水化合物和类胡萝卜素等是不那么复杂的分子。 综上所述一定浓度的重金属Pb促进了斜生栅藻中类胡萝卜素和碳水化合物的累积,而且促进效果20,5和0.5 mg·L-1依次递减。
2.2.2 基于太赫兹光谱的PCA分析
对两个时间点(12和24 h)全光谱数据进行PCA处理,图5(a)代表12 h时样本数据PCA处理结果,样本中PC-1和PC-2分别为85%和12%,其中5与20 mg·L-1实验组聚类明显,对照组、0.5 mg·L-1聚类效果相对较差。 图5(b)代表24 h时样本数据PCA处理结果,样本中PC-1和PC-2分别为87%和9%; 24 h实验组样本整体聚类明显。
PCA图中,12与24 h各胁迫条件下的样本数据都存在一定差异,另外整体上24 h的样本数据聚类效果更好,这与胁迫时间有关,另外 24 h胁迫过程中有12 h的无光合作用阶段不能忽略,在无光照情况下,微藻体内色素、碳水化合物等多种物质的合成均会受到影响[13],如暗环境下影响脂肪酸合成,叶绿素a、核酸等含量减少。 另外铅浓度的增加,导致细胞内碳水化合物和脂类含量增加,蛋白质和磷酸化水平下降。 因此,由于多种物质的复杂变化,24 h的PCA图与12 h的PCA图存在一定的差异。
3 结 论
利用太赫兹、远红外技术对Pb胁迫下斜生栅藻物质含量变化进行了研究。 通过对比各个实验组样本在淀粉(9.1,10.5和17.3 THz)与类胡萝卜素(16.7 THz)特征吸收峰位处的吸光度值,对微藻内淀粉与类胡萝卜素含量的趋势变化进行了研究。 实验结果显示,在胁迫24 h内,0~9 h时间点实验组A,B,C与对照组的特征吸收峰值差异不大,主要是因为实验胁迫时间短,且微藻细胞内物质合成需一定时间。 9 h开始A,B,C与对照组中微藻细胞物质变化得差异逐渐显现,随胁迫时间增长至24 h,实验组A,B,C特征吸收峰逐渐递增,并且实验组C,B和A依次增大,这与随着铅胁迫浓度的增加,碳水化合物和脂类含量增加相一致。 研究中分别使用0.5,5和20 mg·L-1浓度的重金属胁迫微藻,实验中测定出的微藻光合效率表明,铅胁迫抑制了藻类的光合作用,细胞优先合成碳水化合物和类胡萝卜素等分子。 随着铅浓度的增加,碳水化合物和类胡萝卜素增加,这个结果与微藻得到的光谱图中淀粉(9.1,10.5和17.3 THz)与类胡萝卜素(16.7 THz)的特征吸收峰值随时间变化的趋势一致。 通过主成分分析,其中12和24 h下数据的聚类效果明显,说明可以利用太赫兹、远红外光谱识别铅胁迫下藻类内部物质成分的变化。 太赫兹、远红外光谱可作为一种快速,无损方法,用于对铅胁迫下藻体细胞成分的变化研究,从而实现水体污染程度的判定。

图4 重金属胁迫下(a,b,c,d)9.1,10.5,16.7和17.3 THz吸收峰值Fig.4 9.1,10.5,16.7 and 17.3 THz absorption peak under heavy metal stress (a, b, c, d)

图5 不同时间[12 h (a)和24 h (b)]样本PCA对比Fig.5 Comparison of sample PCA at the same time [12 h (a), 24 h(b)]
猜你喜欢
山西大学学报(自然科学版)(2022年2期)2022-06-07
古今农业(2022年1期)2022-05-05
中国油脂(2020年8期)2020-08-12
新农村(浙江)(2019年5期)2019-05-29
吉林农业(2018年24期)2018-12-18
环境保护与循环经济(2017年3期)2017-09-26
环境污染与防治(2016年9期)2016-03-13
中国酿造(2016年12期)2016-03-01
中国果菜(2016年9期)2016-03-01
食品工业科技(2014年23期)2014-03-11
