
水稻地方品种高抗性淀粉含量QTL挖掘与定位
2021-01-02 09:31林静张云辉张所兵陈海元朱晓妹唐伟杰方先文
江苏农业科学 2021年23期
关键词:水稻
林静 张云辉 张所兵 陈海元 朱晓妹 唐伟杰 方先文

摘要:以高抗性淀粉籼稻品种扎西玛和江苏省著名优质粳稻南粳46为亲本构建的重组自交系为材料,运用AOAC法测定了群体家系的抗性淀粉含量,用于QTL分析的分子连锁图谱包含202个SSR分子标记。本研究共检测到2个加性QTL:qRS-6和qRS-8,分别位于第6和第8染色体上,共解释58.38%的表型变异。其中,来源于高值亲本扎西玛的qRS-6贡献率高达51.38%,是一个主效的QTL位点;来源于低值亲本南粳46的qRS-8贡献率只有7%。结果可为高抗性淀粉水稻育种提供资源,进一步挖掘紧密连锁分子标记奠定基础。
关键词:水稻;重组自交系;抗性淀粉;QTLs
中图分类号: S511.01 文献标志码: A
文章编号:1002-1302(2021)23-0058-03
收稿日期:2021-03-16
基金项目:江苏省农业科技自主创新资金[编号:CX(18)1001]。
作者简介:林 静(1981—),女,江苏海安人,硕士,副研究员,主要从事水稻资源研究。E-mail:534663739@qq.com。
水稻大米是亚太地区主食,大量的淀粉存在于大米胚乳中,但其中的抗性淀粉含量很低,热米饭中抗性淀粉含量一般低于1%,冷米饭中也仅为10%~2.1%[1]。随着社会的发展,人们物质生活水平的提高,饮食结构也发生了巨大的改变。体内血糖含量超标人群以及肥胖人群不断增加。稻米中的抗性淀粉具有与水溶性膳食纤维相似的生理功能。高抗性淀粉大米既可以满足饱腹的需求,又能够防止糖尿病患者餐后血糖的快速升高,避免减肥人士摄入过多能量[2-5]。前期抗性淀粉的研究主要集中于功能性试验以及含量测定[6-9],目前,主要集中在稻米抗性淀粉的形成、理化特性及其与直链淀粉的关系等方面[10-16],而对于高抗性淀粉水稻的遗传改良研究相关报道不多[17-18]。
地方品种资源中筛选出的高抗性淀粉水稻存在农艺性状较差、产量不高、品质差、适口性不好等缺点,无法直接用于生产与推广。水稻育种工作者希望通过配置抗性淀粉含量差异较大的遗传群体,对控制抗性淀粉含量的性状进行数量性状基因座(quantitative trait locus,QTL)定位,找到該性状连锁的分子标记,通过分子标记辅助选择,将高抗性淀粉QTL导入到生产上大面积推广的品种中,可缩短育种进程,及早培育出适应生产和市场的高抗性淀粉水稻新品种。笔者所在研究室利用AOAC测定方法筛选出一个高抗性淀粉水稻品种扎西玛,与优良食味水稻品种南粳46配置重组自交系,运用已经构建的遗传图谱对获得的表型数据进行分析,对水稻高抗性淀粉性状进行QTL定位,以期为高抗性淀粉水稻育种提供标记和利用依据。
1 材料与方法
1.1 试验材料
以高抗性淀粉籼稻品种扎西玛为母本,优良食味粳稻品种南粳46为父本,通过单粒传法,获得143个重组自交系(F8)家系。所有试验材料于2017年种植于江苏省农业科学院试验田内,按常规种植,收获晾干后收集存放于江苏省农业科学院种质资源研究中心备用。
1.2 抗性淀粉含量测定
测定米粉中抗性淀粉含量参照罗曦等的方法[19],精确称取0.1 g精米粉,加入胰腺α-淀粉酶,37 ℃水浴振荡消化16 h后,检测样品中抗性淀粉含量。每份样品3次重复。
1.3 QTL分析
采用Wang等开发的QTL检测软件[20]QTL IciMapping V 3.2进行QTL分析,以LOD值2.5为阈值。QTL命名遵循McCouch原则[21]。
2 结果与分析
2.1 亲本与群体抗性淀粉(RS)含量分析
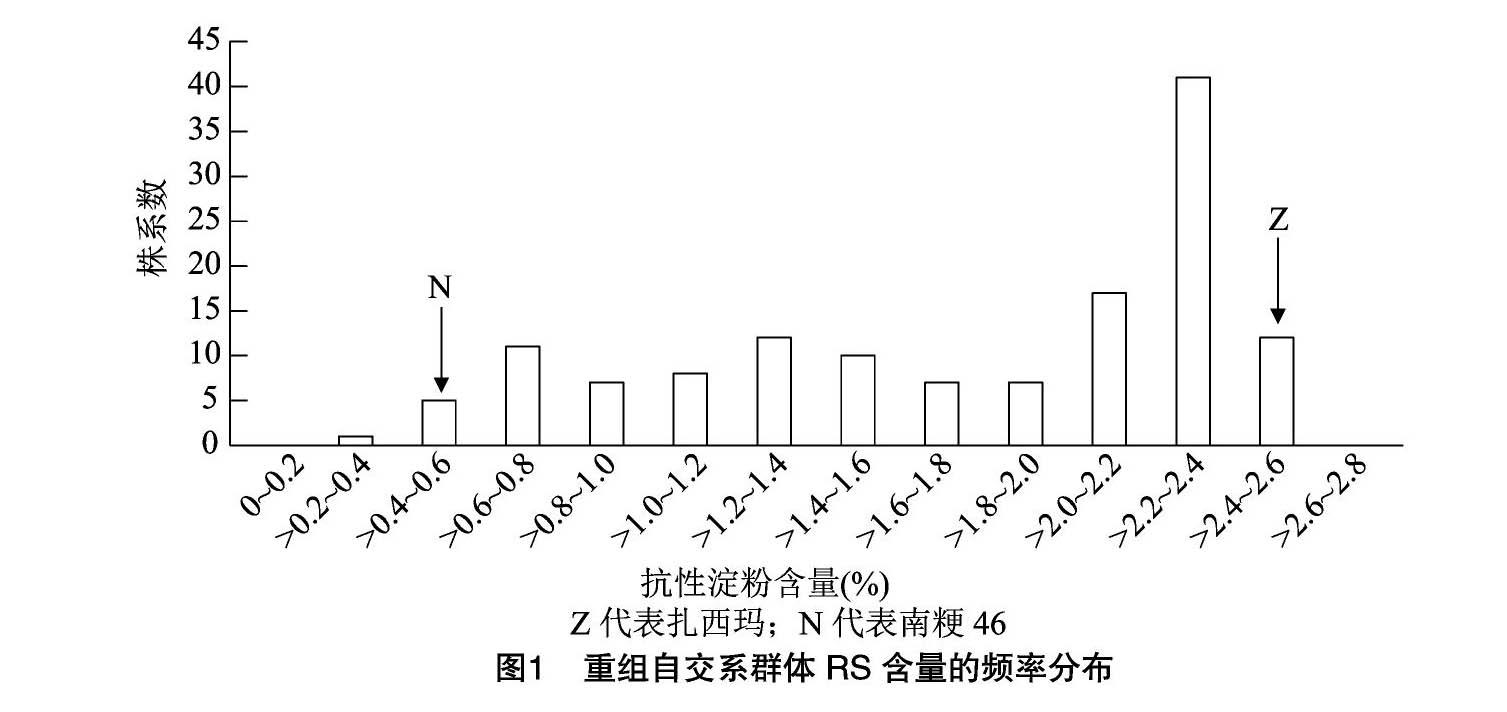
高抗性淀粉品种扎西玛的RS含量达2.42%,南粳46为0.58%,差异有统计学意义。用于QTL检测的重组自交系群体RS含量为0.33%~249%,其变化表现为连续偏正态分布(图1),偏度为-0.659,峰度为-0.970。有一定数量的株系抗性淀粉含量超过亲本,表明该性状是由多基因控制的数量性状。
2.2 抗性淀粉含量的QTL分析
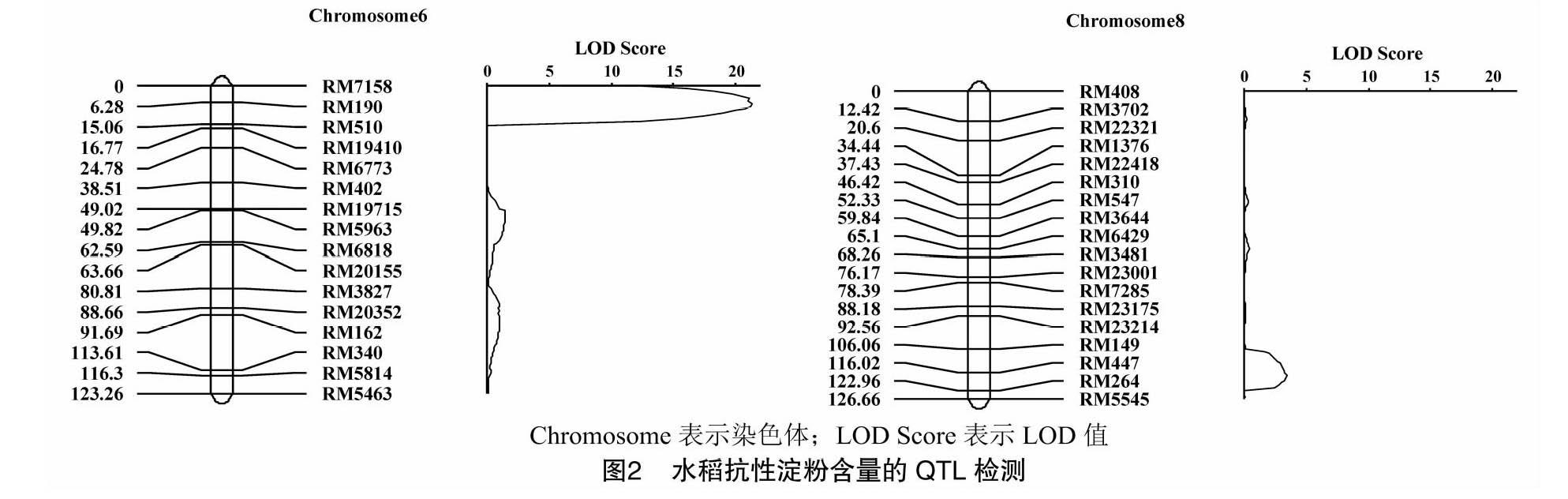
本研究前期运用较均匀分布于水稻12条染色体上的202对SSR多态性分子标记,构建了该重组自交系的分子连锁图谱,全长1 437.3 cM,平均图距8.1 cM,可以用于QTL定位。结合抗性淀粉测定数据,利用QTL IciMapping 3.2分析QTL的位置及遗传效应。共检测到2个控制抗性淀粉含量的加性QTL(qRS-6,qRS-8),分别位于第6和第8染色体上(图2、表1),共解释了58.38%的表型变异。qRS-6 的贡献率高达51.38%,是一个主效的QTL位点,加性效应为正值,表明抗性淀粉含量的增效位点来源于高抗性淀粉亲本扎西玛。qRS-8的贡献率较低,为7.00%,但加性效应为负值,表明增效位点来源于低抗性淀粉亲本南粳46。
3 结论与讨论
大面积推广品种正面临遗传基础单一的问题。充分挖掘地方品种中蕴含的优异基因资源,对推动现代育种具有积极的意义。相关研究表明,水稻大部分性状,如穗长、粒形、抗病、耐盐等,都属于多基因控制的数量性状,遗传基础复杂[22-25]。克隆数量性状基因时,首先要通过构建合适的作图群体鉴定出数量性状相关QTL,分析每个QTL的位置和效应;再通过不断回交的方式将不同的QTL分开,获得相同背景的近等基因系,复杂性状才能被分解成单个孟德尔因子,以便开展下一步的工作。水稻的很多重要性状QTL就是通过这种方法得到了克隆[26-29]。
大米作为我国的传统主食,其淀粉适口性好。而抗性淀粉因其突出的生理作用,成为功能性食品的研究热点。水稻育种工作者希望能培育出高抗性淀粉水稻品种,以满足糖尿病人、减肥人士等特殊人群的需求。稻米抗性淀粉含量测定工作量非常大。因此,水稻育种者希望能找到与高抗性淀粉性状紧密连锁的分子标记,通过标记辅助选择,大大减少测定工作量。对抗性淀粉的遗传学进行研究,也有助于在分子层面解释水稻抗性淀粉的形成和发育机制,对培育高抗性淀粉水稻品种具有重要的意义。
本試验中,扎西玛/南粳46的重组自交系群体中抗性淀粉含量表现为连续的单峰偏正态分布。位于第6染色体上的qRS-6为主效QTL,效应值达51.38%,表明水稻抗性淀粉含量是由少数主效基因和多个微效基因以及非等位基因间互作所控制的数量性状(QTL)。与以往的遗传研究结论[19,30]一致。并且位于第6染色体上的qRS-6与罗曦等利用不同遗传群体检测到的位点基本一致,均位于标记RM510附近,可能是同一位点[19]。后续的研究将集中在与该主效位点紧密连锁的分子标记的开发,为高抗性淀粉水稻育种提供分子标记。
参考文献:
[1]Englyst H,Wiggins H S,Cummings J H.Determination of the non-starch polysaccharides in plant foods by gas-liquid chromatography of constituent sugars as alditol acetates[J]. The Analyst,1982,107(1272):307-318.
[2]Kwak J H,Paik J K,Kim H I,et al. Dietary treatment with rice containing resistant starch improves markers of endothelial function with reduction of postprandial blood glucose and oxidative stress in patients with prediabetes or newly diagnosed type 2 diabetes[J]. Atherosclerosis,2012,224(2):457-464.
[3]Wang H S,Pang G C.Effect of resistant and digestible rice starches on human cytokine and lactate metabolic networks in serum[J]. Cytokine,2017,93:57-65.
[4]Ananda D,Zuhairini Y,Sutadipura N.Resistant starch in cooled white rice reduce glycaemic index[J]. Obesity Research & Clinical Practice,2013,7(1):38.
[5]吴 伟,刘 鑫,杨朝柱,等. 抗性淀粉及预防糖尿病和肥胖症功能稻米研究进展[J]. 核农学报,2006,20(1):60-63,22.
[6]Eggum B O,Juliano B O,Perez C M,et al. The resistant starch,undigestible energy and undigestible protein contents of raw and cooked milled rice[J]. Journal of Cereal Science,1993,18(2):159-170.
[7]Rosin P M,Lajolo F M,Menezes E W.Measurement and characterization of dietary starches[J]. Journal of Food Composition and Analysis,2002,15(4):367-377.
[8]Crosby G A. Resistant starch makes better carbs[J]. Funct Foods & Nutrace,2003(6):34-36.
[9]Kim W K,Chung M I K,Kang N,et al. Effect of resistant starch from corn or rice on glucose control,colonic events,and blood lipid concentrations in streptozotocin-induced diabetic rats[J]. The Journal of Nutritional Biochemistry,2003,14(3):166-172.
[10]Ashwar B A,Gani A,Wani I A,et al. Production of resistant starch from rice by dual autoclaving-retrogradation treatment:invitro digestibility,thermal and structural characterization[J]. Food Hydrocolloids,2016,56:108-117.
[11]Sun J A,Wu D X,Xu J Y,et al. Characterisation of starch during germination and seedling development of a rice mutant with a high content of resistant starch[J]. Journal of Cereal Science,2015,62:94-101.
[12]Itoh Y,Crofts N,Abe M,et al. Characterization of the endosperm starch and the pleiotropic effects of biosynthetic enzymes on their properties in novel mutant rice lines with high resistant starch and amylose content[J]. Plant Science,2017,258:52-60.
[13]Kiatponglarp W,Tongta S,Rolland-Sabaté A,et al. Crystallization and chain reorganization of debranched rice starches in relation to resistant starch formation[J]. Carbohydrate Polymers,2015,122:108-114.
[14]Pongjanta J,Utaipattanaceep A,Naivikul O,et al. Debranching enzyme concentration effected on physicochemical properties and α-amylase hydrolysis rate of resistant starch type III from amylose rice starch[J]. Carbohydrate Polymers,2009,78(1):5-9.
[15]Hung P V,Vien N L,Phi N T L.Resistant starch improvement of rice starches under a combination of acid and heat-moisture treatments[J]. Food Chemistry,2016,191:67-73.
[16]Chen M H,Bergman C J,McClung A M,et al. Resistant starch:variation among high amylose rice varieties and its relationship with apparent amylose content,pasting properties and cooking methods[J]. Food Chemistry,2017,234:180-189.
[17]牟方贵,闫宗武,冉瑞林,等. 水稻抗性淀粉相关SSR标记的初步研究[J]. 分子植物育种,2008,6(3):432-438.
[18]罗 曦,黄锦峰,朱永生,等. 水稻功米3号高抗性淀粉性状的遗传分析[J]. 农业生物技术学报,2014,22(1):10-16.
[19]罗 曦,黄锦峰,朱永生,等. 稻米抗性淀粉含量测定方法的比较分析[J]. 福建农业学报,2011,26(5):822-826.
[20]Wang J K,Wan X Y,Li H H,et al. Application of identified QTL-marker associations in rice quality improvement through a design-breeding approach[J]. Theoretical and Applied Genetics,2007,115(1):87-100.
[21]Mccouch S R,Cho Y G,Panl E,et al. Report on QTL nomenclature[J]. Rice Genet Newsl,1997,14:11-13.
[22]张云辉,张所兵,林 静,等. 长穗型水稻地方品种中穗长QTL的挖掘与定位[J]. 分子植物育种,2018,16(12):3967-3972.
[23]游 佳,谷 晗,朱 泽,等. 水稻粒质量和粒形QTL定位及粒长位点qGL3.2的鉴定[J]. 南京农业大学学报,2019,42(4):612-621.
[24]林 静,张所兵,张云辉,等. 利用染色体片段置换系定位水稻抗纹枯病QTLs[J]. 江苏农业学报,2013,29(4):691-695.
[25]林 静,张云辉,陈海元,等. 水稻地方品种苗期耐盐QTL的定位[J]. 华北农学报,2019,34(增刊1):1-5.
[26]Weng J F,Gu S H,Wan X Y,et al. Isolation and initial characterization of GW5,a major QTL associated with rice grain width and weight[J]. Cell Research,2008,18(12):1199-1209.
[27]Zhang X J,Wang J F,Huang J,et al. Rare allele of OsPPKL1 associated with grain length causes extra-large grain and a significant yield increase in rice[J]. Proceedings of the National Academy of Sciences of the United States of America,2012,109(52):21534-21539.
[28]Ishimaru K,Hirotsu N,Madoka Y,et al. Loss of function of the IAA-glucose hydrolase gene TGW6 enhances rice grain weight and increases yield[J]. Nature Genetics,2013,45(6):707-711.
[29]Li Y B,Fan C C,Xing Y Z,et al. Chalk5 encodes a vacuolar H+-translocating pyrophosphatase influencing grain chalkiness in rice[J]. Nature Genetics,2014,46(4):398-404.
[30]孫春龙. 水稻抗性淀粉含量性状的QTL定位及遗传分析[D]. 长春:吉林大学,2013.
猜你喜欢
幼儿100(2023年39期)2023-10-23
中国土壤与肥料(2021年5期)2021-12-12
今日农业(2021年21期)2021-11-26
军事文摘(2021年22期)2021-11-26
今日农业(2021年14期)2021-10-14
金桥(2021年7期)2021-07-22
今日农业(2020年20期)2020-11-26
时代农机(2016年6期)2016-12-01
中国化肥信息(2016年45期)2016-06-15
