
孕穗期剪叶对稻田甲烷排放的影响
2021-01-02 10:29王澜赵步洪张亚军蒋红亮李国明刘立军陈云
江苏农业科学 2021年23期
王澜 赵步洪 张亚军 蒋红亮 李国明 刘立军 陈云

摘要:水稻根系对稻田甲烷排放有重要影响,剪叶可以调节水稻源库关系和根冠生长,但其对稻田甲烷排放的调控效应及机制尚不清楚。以扬稻6号(籼稻)和日本晴(粳稻)为材料,在大田栽培条件下对孕穗期水稻植株进行剪叶处理,研究植株与土壤的相关性状及其对稻田甲烷排放的影响。结果表明,与对照相比(未剪叶),剪叶处理显著降低了稻田甲烷排放通量,扬稻6号和日本晴2个品种的甲烷排放通量在剪叶36 h后分别比对照降低了23.0%和22.1%。剪叶极显著降低了水稻叶面积指数,但其对根系形态特征、根系泌氧能力和氧化力无显著影响;剪叶处理使根中非结构性碳水化合物(NSC)和根系分泌物总有机碳(TOC)含量显著降低,但使2个品种根中NSC/TOC分别增加了33.1%和28.9%。Mantel检验分析表明,根中NSC/TOC是影响稻田甲烷通量和产甲烷菌最主要的因素。这些结果表明,剪叶处理可以调节光合产物在根系和根际土壤中的分配,提高NSC/TOC,导致土壤中产甲烷菌数量下降进而减少稻田甲烷排放。
关键词:水稻;剪叶;甲烷排放;根系;非结构性碳水化合物;NSC/TOC
中图分类号: S511.01 文献标志码: A
文章编号:1002-1302(2021)23-0071-06
收稿日期:2021-06-15
基金項目:江苏省农业科技自主创新资金[编号:CX(17)3042];江苏省自然科学基金(编号:BK20181214);国家自然科学基金(编号:31871557、32071947)。
作者简介:王 澜(2001—),女,江苏镇江人,主要从事水稻生理生态研究。E-mail:3465393772@qq.com。
通信作者:陈 云,博士,副教授,主要从事水稻生理生态研究。E-mail:yunchen@yzu.edu.cn。
全球气候变暖一直备受关注。甲烷(CH4)是第二大温室气体,增温潜势约为二氧化碳(CO2)的28倍,其温室效应仅次于CO2[1]。水稻是全球最重要的粮食作物之一。然而,稻田也是甲烷的重要排放源,约20%的人为甲烷排放来自稻田[2]。因此,控制稻田甲烷排放对缓解全球温室效应具有重要作用。
水稻的植株脱落物、根系分泌物以及土壤中的有机质等可以为土壤中的产甲烷菌提供甲烷产生的底物[3-4]。叶片光合作用固定的碳是水稻植株生物碳积累及籽粒充实的最主要来源。剪叶或疏花处理可以调节水稻的源库关系,影响植株的干物质积累及光合同化物向根系的转运[5]。有研究表明,在苗期或孕穗期剪叶后水稻的根质量及根系活力都有所增加[6]。虽然水稻叶片的光合作用产物不直接作为甲烷产生的底物,但植物-土壤系统中光合碳的分配与周转非常迅速,使用同位素标记的方法发现光合作用固定的碳在几小时内即可分配到根际土壤,进而在稻田排放的甲烷中检测到标记的甲烷[7-8]。孕穗期是稻田甲烷排放的高峰期之一,此期甲烷排放量可占全生育期总甲烷排放的40%以上[9]。但孕穗期光合产物在根系中的分配及其与土壤性状的关系的研究较少,生理机制也尚不明确。
本研究选用籼稻扬稻6号和粳稻日本晴为材料,通过在孕穗期剪叶处理减少水稻叶片光合源,研究剪叶对光合同化碳在根系和土壤中分配的影响及其对土壤中甲烷相关菌群的调节。旨在进一步揭示水稻植株对稻田甲烷排放的影响机制,为稻田甲烷减排提供理论与实践依据。
1 材料与方法
1.1 试验材料与试验设计
试验于2019年和2020年在扬州大学江苏省作物栽培生理重点实验室试验农场(119°25′E、32°30′N)进行。供试品种为扬稻6号(YD 6)和日本晴(NIP)。试验田前茬作物为小麦,土壤为沙壤土,有机质含量为23.0~25.8 g/kg,速效钾含量为60.8~65.4 mg/kg,有效氮含量为111~123 mg/kg,速效磷含量为30.2~34.3 mg/kg。水稻播种期为5月15日,移栽期为6月10日,收获期为10月8—12日。株行距分别为0.16、0.25 m,单本种植。全生育期施用氮肥(折合纯氮)270 kg/hm2,其中基肥占60%,分蘖肥占10%,穗肥占30%。每小区在移栽前一次性基施过磷酸钙(含P2O5 13.5%) 300 kg/hm2 和氯化钾(含K2O 52%)195 kg/hm2。整个水稻生育期按照常规灌溉进行水分管理,中期搁田1周。定期除草、施用农药并严格控制杂草和病虫害。
水稻生长至孕穗期时,将每株水稻茎蘖上倒1叶至倒3叶共3张叶片沿叶脉纵向剪去1/2(leaf-cutting),以不剪叶为对照(control)。每个处理各 15 m2,3次重复。
1.2 测定项目与方法
1.2.1 稻田甲烷的收集与分析 使用静态箱-气相色谱法采集并测定稻田排放的甲烷。剪叶处理后的48 h内,每隔12 h取气1次。采集的气体用气相色谱仪分析甲烷浓度。甲烷通量计算公式参照Chen等的方法[9],具体如下:
F=MV1×PP1×T1T×H×dCdt。
式中:F为气体排放通量;M为气体摩尔质量;V1、P1和T1分别为标准状态下气体的摩尔体积、空气压强及绝对温度;P和T分别为采样点的空气气压和绝对温度;H为箱体的高度;dCdt为单位时间内箱子内甲烷浓度变化率。
1.2.2 植株取样与测定 剪叶处理36 h时,每小区选取有代表性的植株3株,用便携式叶面积仪测定叶面积指数,用叶绿素仪测量剑叶叶绿素的相对含量(SPAD值)。用便携式光合仪测量光合指标。在对上述性状进行测量的同一天,每个小区采集12株水稻,取样时尽量减少边际效应。将其中3株用于根系性状测定,并测定根中非结构碳水化合物(nonstructural carbohydrates,NSC)含量,3株用于根系氧化活力测定,3株用于根系泌氧能力测定,剩余3株用于收集和分析根分泌物。
1.2.3 根中NSC含量测定含量 参照Wang等的方法[10]提取并测定根中可溶性糖和淀粉含量。NSC浓度是指可溶性糖和淀粉的含量之和。每株水稻根中NSC含量以根生物量与单位质量根中NSC含量的乘积表示。
1.2.4 根系性状 用液压气动冲洗装置将根洗净,并从根基部分离。用扫描仪扫描完根系图像后,用WinRHIZO根系分析系统(Regent Instruments Inc.,Canada)分析,计算单株根数、根长、根直径和根表面积[9]。之后将根系放入烘箱中在105 ℃下烘 30 min,然后在75 ℃下烘干至质量恒定,测定根干质量。用α-萘胺法测定根系氧化活力[11],用柠檬酸比色法测定根系泌氧能力[12]。
1.2.5 根系分泌物的收集和分析 根系分泌物的收集和稻田取气同时进行,按照Chen等的描述[9]进行,使用总有机碳分析仪分析样品中的有机碳含量。
1.2.6 土壤样品采集与分析 使用直径为3 cm的土壤采样器从水稻根系中心2~10 cm深处收集新鲜土壤。样本用于实时荧光定量PCR测定土壤甲烷相关菌群数量。以提取的土壤DNA为模板,使用ABI 7500定量分析土壤产甲烷菌和甲烷氧化菌的数量。PCR反应体系包括5 μL qPCR Master Mix、0.5 μL primer mix和4.5 μL模板。扩增条件为 95 ℃ 预变性30 s;95 ℃变性10 s,55 ℃退火30 s,72 ℃延伸30 s,40个循环10 s;最后72 ℃延伸 1 min。引物[9]为产甲烷菌(mcrA):MLF(GGTATGGAGCAGTACGAGGAGTTC)和MLR(GTAGCCGAAGAAGCCGAGACG),甲烷氧化菌(pmoA):A189F(GGTGACTGGGACTTCTGG)和MB661R(CCGTGCAACGTCYTTACC)。以1 g干质量土壤中的基因拷贝数表示土壤菌群的数量。
1.3 数据处理与分析
统计分析软件SPSS 18.0 用于独立样品t检验分析。R语言中的ggplot2程序包用于绘制折线图和箱形图,fmsb程序包用于绘制雷达图,ggcor程序包用于Mantel test检验统计植物性状变量矩阵与甲烷排放通量之间的相关性。
试验重复2年,除箱形图呈现所有数据点外,其余均以2年数据的平均结果表示。
2 结果与分析
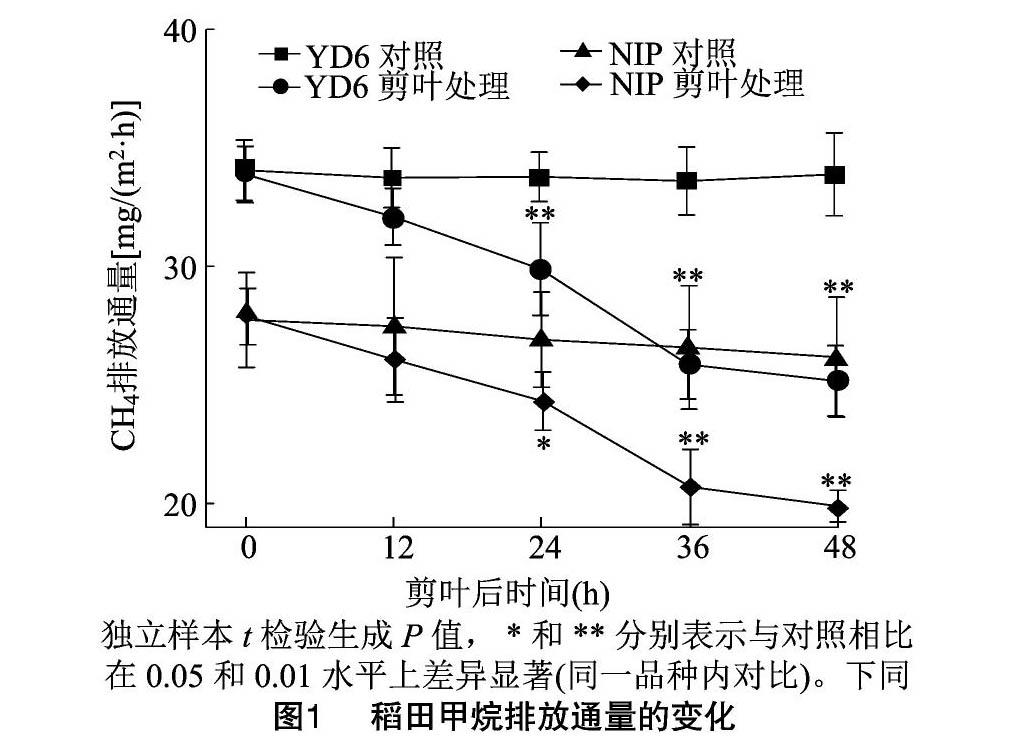
2.1 剪叶处理对稻田甲烷排放通量的影响
由图1可知,剪叶处理显著降低了YD 6和NIP的稻田甲烷排放通量。2个品种的稻田甲烷排放通量在剪叶处理后逐渐降低。与对照不剪叶相比,剪叶后36 h差异均达极显著水平。此时,YD 6 和NIP分别比对照组极显著降低了23.0%和22.1%。36 h 之后甲烷排放通量趋于平稳。
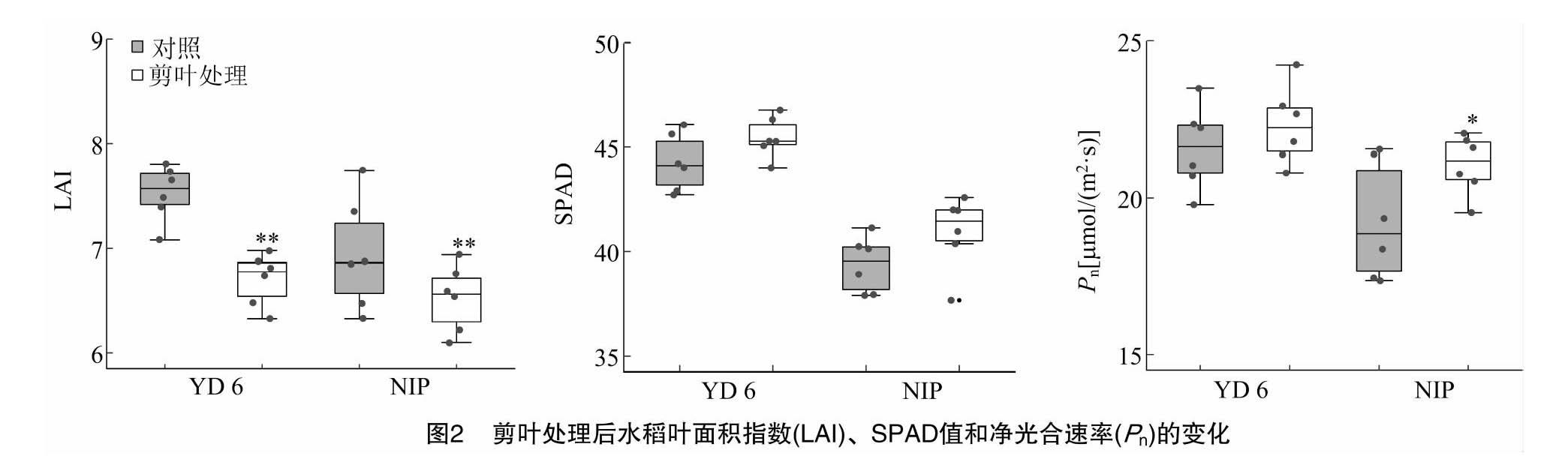
2.2 剪叶处理对水稻叶片光合指标的影响
由图2可知,YD 6 的叶面积指数(LAI)和NIP差异不显著。与对照相比,剪叶处理使2个品种的LAI分别下降了11.0%和6.0%。剪叶处理对叶片SPAD值和净光合速率(Pn)的影响一致,二者均呈现增加趋势,t检验表明仅有NIP的Pn变化呈显著水平。双因素分析结果表明,SPAD值和Pn在品种和处理间均有差异,但上述光合指标在品种和处理之间无互作效应(表1)。
2.3 剪叶处理对水稻根系形态生理的影响
由图3可知,处理36 h后,与对照相比,剪叶处理对YD 6 和NIP 的主要根系形态指标(根长、根质量、根直径等)均无显著影响。根系氧化力及根系泌氧能力的变化也未达显著水平。表明剪叶处理后36 h并不影响水稻根系的主要形态特征,对根系活力的影响也较小。
根系非结构性碳水化合物及根系分泌物中总有机碳(TOC)含量测定结果表明,与对照相比,剪叶处理后YD 6和NIP根中NSC含量分别降低了19.6% 和 25.6%, TOC含量分别降低了40.1% 和42.2%(图3)。2个品种NSC/TOC的比值则分别增加了33.1%和28.9%,且与对照相比差异达极显著水平(图4)。
2.4 剪叶处理对甲烷相关菌群数量的影响
如图5所示,剪叶处理后YD 6 和NIP根际土壤中产甲烷菌数量均极显著下降,分别下降31.6%和43.2%;而甲烷氧化菌数量无显著变化。
甲烷通量与甲烷相关菌群的相关性分析结果表明,剪叶处理下甲烷通量与产甲烷菌显著相关,与甲烷氧化菌相关性不显著(图6)。
2.5 植株性状与甲烷通量和产甲烷菌的Mantel检验分析
通过Mantel 检验分析甲烷排放通量和产甲烷菌数量与水稻植株性状的相关关系。由图7可以看出,在所观测的水稻植株的10个性状中,根系中NSC/TOC比值是影响甲烷通量和产甲烷菌数量的主要因素。NSC和TOC为影响甲烷通量和产甲烷菌数量的次要因素。其他植株性状与甲烷排放通量和产甲烷菌数量均无显著相关性。表明水稻根系NSC/TOC的比值可以调控产甲烷菌数量,从而影响稻田甲烷排放。
3 討论与结论
3.1 剪叶对稻田甲烷排放的影响
前人通过同位素标记等手段研究发现稻田产甲烷菌所需碳源的60%来自以根分泌形式分配到根际的光合碳,其余部分来自水稻植株脱落物以及沉积有机物的降解[3,8]。剪叶处理会降低植株的总光合产物量[13]。也有报道认为,减源处理使水稻剑叶的SPAD值和光合速率增加,表现为反馈促进效应[14]。通过人工剪穗和转基因的方式减少或增加光合产物向穗部运输,可改变根系分泌的光合产物量从而增加或减少稻田甲烷排放[4,15-16]。本研究观察到剪叶虽然增加了水稻单叶的SPAD值和净光合能力,但由于LAI的极显著降低,根系分泌物中有机碳含量也显著降低,土壤中产甲烷菌的数量降低,最终导致甲烷排放减少。
吴降星等研究表明,剪叶面积为30%~50%时,剪叶处理后水稻根系干物质量和活力显著增加[6]。一定程度的剪叶率激发的根系活力增加对于延长植株叶片后期功能、延缓叶片叶绿素的降解、维持较大的光合作用面积、延长叶片寿命和功能期及增加灌浆物质的制造均有积极的作用[17-18]。本研究中剪叶处理未显著影响根系形态和生理指标,这可能与本研究处理时间较短(0~48 h)有关。
3.2 剪叶影响稻田甲烷排放的原因分析
在稻田甲烷的循环系统中,水稻植株、产甲烷菌和甲烷氧化菌是决定稻田甲烷释放量的关键因素,而根际甲烷相关菌群又与水稻植株所营造的生境密切相关[9]。前人研究表明,水稻的地上部性状如分蘖数、株高等,地下部性状如根活力对稻田甲烷排放的影响不尽相同[19]。剪叶处理后会增大分配到根系的光合产物的利用比例,从而对根系生长和根系活力进行生理补偿[6]。NSC是光合碳分配的主要形式,根系NSC含量表明光合生产在根系生长和根系呼吸损失之间的平衡,而TOC含量则表现为根沉积能力[20-21]。在本试验条件下,剪叶处理同时降低了NSC和TOC含量,但极显著增加了NSC/TOC。Mental检验分析表明,剪叶处理甲烷排放的变化主要是源于产甲烷菌数量的变化而非甲烷氧化菌的变化,并且水稻根系特征NSC/TOC是驱动甲烷排放差异的主要因素。
综上所述,剪叶后减少了水稻光合产物向根际土壤的分泌,降低了土壤中产甲烷菌的数量,导致甲烷排放减少。水稻根系NSC/TOC是影响稻田产甲烷菌和甲烷排放通量的重要因素。该研究为深入理解稻田甲烷排放机制提供了新思路,为通过调节水稻植株生长实现甲烷减排提供了途径。
参考文献:
[1]Schaefer H. On the causes and consequences of recent trends in atmospheric methane[J]. Current Climate Change Reports,2019,5(4):259-274.
[2]Conrad R.The global methane cycle:recent advances in understanding the microbial processes involved[J]. Environmental Microbiology Reports,2009,1(5):285-292.
[3]Tokida T,Adachi M,Cheng W G,et al. Methane and soil CO2 production from current-season photosynthates in a rice paddy exposed to elevated CO2 concentration and soil temperature[J]. Global Change Biology,2011,17(11):3327-3337.
[4]Su J,Hu C,Yan X,et al. Expression of barley SUSIBA2 transcription factor yields high-starch low-methane rice[J]. Nature,2015,523(7562):602-606.
[5]He A B,Wang W Q,Jiang G L,et al. Source-sink regulation and its effects on the regeneration ability of ratoon rice[J]. Field Crops Research,2019,236(1):155-164.
[6]吳降星,郑许松,周光华,等. 不同生育期剪叶对水稻生长、产量及生理的影响[J]. 应用昆虫学报,2013,50(3):651-658.
[7]Kuzyakov Y,Gavrichkova O.Review:Time lag between photosynthesis and carbon dioxide efflux from soil:a review of mechanisms and controls[J]. Global Change Biology,2010,16(12):3386-3406.
[8]Yuan Q A,Pump J,Conrad R. Partitioning of CH4 and CO2 production originating from rice straw,soil and root organic carbon in rice microcosms[J]. PLoS One,2012,7(11):e49073.
[9]Chen Y,Li S Y,Zhang Y J,et al. Rice root morphological and physiological traits interaction with rhizosphere soil and its effect on methane emissions in paddy fields[J]. Soil Biology and Biochemistry,2019,129:191-200.
[10]Wang Z Q,Zhang W Y,Beebout S S,et al. Grain yield,water and nitrogen use efficiencies of rice as influenced by irrigation regimes and their interaction with nitrogen rates[J]. Field Crops Research,2016,193:54-69.
[11]Ota Y. Diagnostic methods for the measurement of root activity in rice plant[J]. Japan Agricultural Research Quarterly,1970,5(3):1-6.
[12]Kludze H K,DeLaune R D,Jr Patrick W H.Aerenchyma formation and methane and oxygen exchange in rice[J]. Soil Science Society of America Journal,1993,57(2):386-391.
[13]杨靖睿,曹培培,杨 凯,等. CO2浓度升高和不同氮肥水平下源库处理对粳稻茎鞘非结构性碳水化合物积累和转运的影响[J]. 生态学杂志,2021,40(3):615-626.
[14]李 杰,冯跃华,牟桂婷,等. 剪叶、粘叶处理对水稻剑叶主脉两侧SPAD值及籽粒产量的影响[J]. 中国稻米,2018,24(6):40-46.
[15]van der Gon H A C D,Kropff M J,van Breemen N,et al. Optimizing grain yields reduces CH4 emissions from rice paddy fields[J]. PNAS,2002,99(19):12021-12024.
[16]Du L,Wang Y F,Shan Z,et al. Comprehensive analysis of SUSIBA2 rice:The low-methane trait and associated changes in soil carbon and microbial communities[J]. Science of the Total Environment,2021,764:144508.
[17]梁 嫻,赵建华,罗 充.不同修剪方式对黑麦草叶绿素、根系活力的影响[J]. 贵州师范大学学报(自然科学版),2010,28(3):4-7.
[18]凌启鸿,凌 励.水稻不同层次根系的功能及对产量形成作用的研究[J]. 中国农业科学,1984,17(5):3-11.
[19]江 瑜,管大海,张卫建.水稻植株特性对稻田甲烷排放的影响及其机制的研究进展[J]. 中国生态农业学报,2018,26(2):175-181.
[20]Dietze M C,Sala A,Carbone M S,et al. Nonstructural carbon in woody plants[J]. Annual Review of Plant Biology,2014,65(1):667-687.
[21]Xiao M L,Zang H D,Ge T D,et al. Effect of nitrogen fertilizer on rice photosynthate allocation and carbon input in paddy soil[J]. European Journal of Soil Science,2019,70(4):786-795.
猜你喜欢
现代园艺(2022年7期)2022-11-19
今日农业(2021年21期)2021-11-26
军事文摘(2021年22期)2021-11-26
今日农业(2021年14期)2021-10-14
四川劳动保障(2021年3期)2021-06-09
现代园艺(2017年13期)2018-01-19
西南农业学报(2016年5期)2016-05-17
中国农业文摘-农业工程(2016年5期)2016-04-12
水土保持通报(2014年5期)2014-06-09
