
不同种植年限对凤丹牡丹根际真菌群落组成和多样性的影响
2021-01-02 18:58耿晓东周英汪成忠钱剑林
江苏农业科学 2021年23期
耿晓东 周英 汪成忠 钱剑林
摘要:连作障碍对牡丹的生长发育会产生严重的影响,根际微生物环境会对这种改变做出相应的反馈,因此,对连作状态下的不同种植年限凤丹牡丹根际微生物环境的研究有重要意义。采用Illumina 高通量测序技术,以苏州地区的3、6、10年生凤丹牡丹根际土壤为研究对象,对根际真菌群落进行测序分析,研究真菌ITS rDNA可变区的丰富度和多样性指数。测序后不同种植年限根际土中共得到1 020 520条原始序列,序列过滤拼接后可得613 250 条序列(tags);3组土壤样品共含有891个分类操作单元(OTUs),涵盖了5 门、115纲、15目、175 科、230属、320种菌群;3年生凤丹牡丹根际优势菌群为球囊菌门(Glomeromycota)、木耳科(Auriculariales)、盘菌科(Pezizaceae)、球腔菌科(Mycosphaerellaceae);6年生凤丹牡丹的优势真菌群落主要有肉座菌目(Hypocreales)、鬼伞属(Coprinopsis)、巢菌科(Nidulariaceae)、黑蛋巢菌(Cyathus)、银耳目(Tremellales);10年生凤丹牡丹的优势真菌群落主要有丝膜菌属(Cortinarius)、蜡皮马鞍蜡壳菌(Helvellosebacina)、革菌属(Thelephora)。α多样性分析和β多样性分析均表明,不同种植年限凤丹牡丹根际土菌群群落多样性所呈现的规律为6年生>3年生>10年生。此外还发现,在连作状态下不同种植年限的凤丹牡丹根际真菌群落多样性之间有显著差异,菌群群落多样性呈先升高后降低的趋势。
关键词:凤丹牡丹;多样性;根际微生物;连作;群落结构
中图分类号: S685.110.4;S154.3 文献标志码: A
文章编号:1002-1302(2021)23-0145-07
收稿日期:2021-09-27
基金项目:江苏省现代农业产业技术体系建设专项(编号:JATS[2020]340);江苏省苏州市农业技术应用基础研究项目(编号:SNG2018052);苏州农业职业技术学院科技发展专项(编号:19QN1010、PY2003)。
作者简介:耿晓东(1979—),男,江苏扬中人,副教授,主要从事观赏园艺新品种繁育及产业化推广研究。E-mail:7587633@qq.com。
通信作者:钱剑林,教授,主要从事花卉新品种选育及产业化示范推广。E-mail:qianjl03@126.com。
凤丹牡丹(Paeonia ostii)即杨山牡丹,属于林下小灌木,是当前油用牡丹的重要品种。目前,凤丹牡丹和紫斑牡丹(P. rockii)在我国牡丹油用方面表现突出。牡丹籽油中含有丰富的亚麻酸等不饱和脂肪酸[1-2],以及白藜芦醇等药用成分[3-4],具有抑制癌细胞、降低血脂、改善神经功能等功效[5-6]。油用牡丹种植区域的扩增与栽植时期的延长,使得植株的生长活力逐年减弱,此外,还会导致植株死亡。如果在种植过牡丹植株的土壤中再次种植牡丹,所种植的牡丹会出现生长缓慢、根系不发达等现象[7],可见连作障碍已对油用牡丹产业发展产生了严重的限制作用。
根际土壤微生物在植物生长发育阶段具有重要作用,可对植物生长和土壤状况产生影响[8]。其中,植物体内的内生菌在植物生长过程中具有促进作用,同时也改善了某些天然产物的品质[9-10]。内生菌与植株长时间共同进化后,对生态学功能有所保留,或对子代赋予了某些功能特性[11]。有研究表明,微生物群落结构的改变会影响牡丹的生长和土壤酶活性。例如,郭丽丽等发现,根际土壤的微生物结构功能的不平衡会引起植物的连作障碍,产生不利的影响[12]。韩继刚等的研究显示,根际的多黏类芽孢杆菌可以对凤丹牡丹的生根发芽和幼苗的生产发育起到促进的作用[13]。郑艳等的研究结果表明,牡丹中的某些药用成分或许与微生物之间有一定的关联[14]。史冬燕等的研究表明,根际微生物活性明显比非根际土壤微生物的活性高[15]。这些学者的研究大部分集中在药用、油用以及其他方面,而在自然环境中,研究凤丹牡丹微生物的特点相对困难。因此,本试验利用Illumina高通量测序技术对3个不同种植年限的凤丹牡丹根际微生物进行研究,探讨其组成特征,以根际微生态角度分析不同种植年限凤丹牡丹根际状态下真菌群落结构及其多样性变化,从而阐明连作后凤丹牡丹根际真菌群落结构的变化规律,为解决凤丹牡丹连作障碍提供科学依据,并为推动中国油用牡丹产业化发展提供理论依据。
1 材料与方法
1.1 研究区概况
本试验于苏州农业职业技术学院(30°52′14. 28″N,118°1′13.39″E)进行,该区域属亚热带季风型大陆气候,海拔为 210~230 m,年均气温在 15.6 ℃ 左右,年均降水量为 620 mm,主要的土壤为褐土。分别设置不同时期栽培的油用牡丹,长期观察栽培时间对凤丹牡丹产生的影响。
1.2 试验材料
供试土壤取自苏州农业职业技术学院凤丹牡丹栽培基地。选取种植年限分别为3、6、10年的油用凤丹牡丹根际土壤作为本试验的研究对象,在种子收获期(2020年8月)取样。依照五点取样法,以随机挑选的方式在3个种植年限中各选取3株生长状况相同的植株(每处理3个重复)。每株植株都以主茎为中心,在半径长度为30~40 cm的范围内进行取样,取样深度为0~20 cm,用四分法将取出的土壤混匀,去除土壤表面的杂质后,再将该土壤样品进行分装,放入标记好的相应无菌牛皮纸袋中,置于冰盒中保存,并迅速帶回实验室进行后续处理。
1.3 试验方法
1.3.1 土壤微生物基因组DNA提取及质量检测 用FastDNA SPIN Kitfor Soil(USA)试剂盒对3个种植年限的15个土壤样本提取总DNA;2%琼脂糖电泳和 Agilent 2100 Bioanalyzer 用于检测 DNA 样品的降解程度和样品中存在的杂质;利用NanoPhotometer( IMPLEN,德国) 分光光度计对DNA 样品纯度进行测定;Qubit 2.0 Flurometer 用于检测 DNA 样品浓度;DNA 样品置于-20 ℃ 保存,供后续使用。
1.3.2 PCR 扩增细菌 在ITS rRNA 基因ITS1-ITS2可变区中选取 20 ng 土壤基因组 DNA 作为模板,使用TaKaRa EX-tag酶,ITS rRNA 基因ITS1可变区ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA-3′)和ITS2 (5′-GCTGCGTTCTTCATCGATGC-3′)作为引物,扩增ITS rRNA基因序列ITS1高变区,富集目的片段。
1.3.3 ITS rRNA 文库质检及测序 文库构建结束后,Qubit 2. 0被用来对完成的文库进行初步的定量分析;文库被稀释到1 ng/μL,然后用Agilent 2100检测库的片段大小。如果片段大小与预期结果一致,则用Bio-RAD CFX 96荧光定量PCR仪进行实时荧光定量(qPCR),以准确量化文库的有效浓度,从而确保文库的质量。检测合格的文库运用 Illumina MiSeq 对 ITS rRNA 基因序列的ITS1区进行高通量测序。
1.3.4 信息分析流程 在测序序列中,低质量的碱基和拼接污染序列被清除,并对数据进行过滤以获得可信的目标序列;针对会出现末尾重叠的状况,运用算法PEAR对双端测序序列拼合连接;得到的标签和分类操作单元(OTUs)序列进行比较,把相似度高于97% 的序列当作同一种类型的 OTUs,随后进行物种识别。运用软件QIIME 1. 8. 0对拼接后的序列进行 OTUs 交叠分析、聚类分析、系统发生树构建、α多样性、β多样性等相关分析。
2 结果与分析
2.1 土壤样品测序深度评估
油用凤丹牡丹根际真菌ITS rRNA 测序共获得5 102 600条序列,最终转化为1 020 520 条原始测序序列,共 230.20 Mb 原始序列片段,对原始序列片段进行过滤,即除去接头污染、劣质序列和N含量高于5%的序列,过滤结束后得到180 Mb有效序列(Clean reads),其中各个样品中平均有40 200条,数量范围为36 825~42 663 条。运用Q30 碱基百分比的统计方式对有效序列的质量进行统计,统计结果的指标越高则表明测序碱基中错误率低于0.1%的碱基所占的比例越大。全部样本的统计Q30碱基百分比的指标结果都在90%附近,即测序有效序列错误率都低于0.1%,用以建库质量较高。此外,Q30碱基在每个样本中所占的比例都相差不大,说明过滤后的样本所具有的均一性效果好。对序列双末端重叠连接后进行测序,测序后出现 331 252 条序列(tags),平均大小为425 bp。
2.2 不同种植年限油用凤丹牡丹根际土壤真菌OTUs
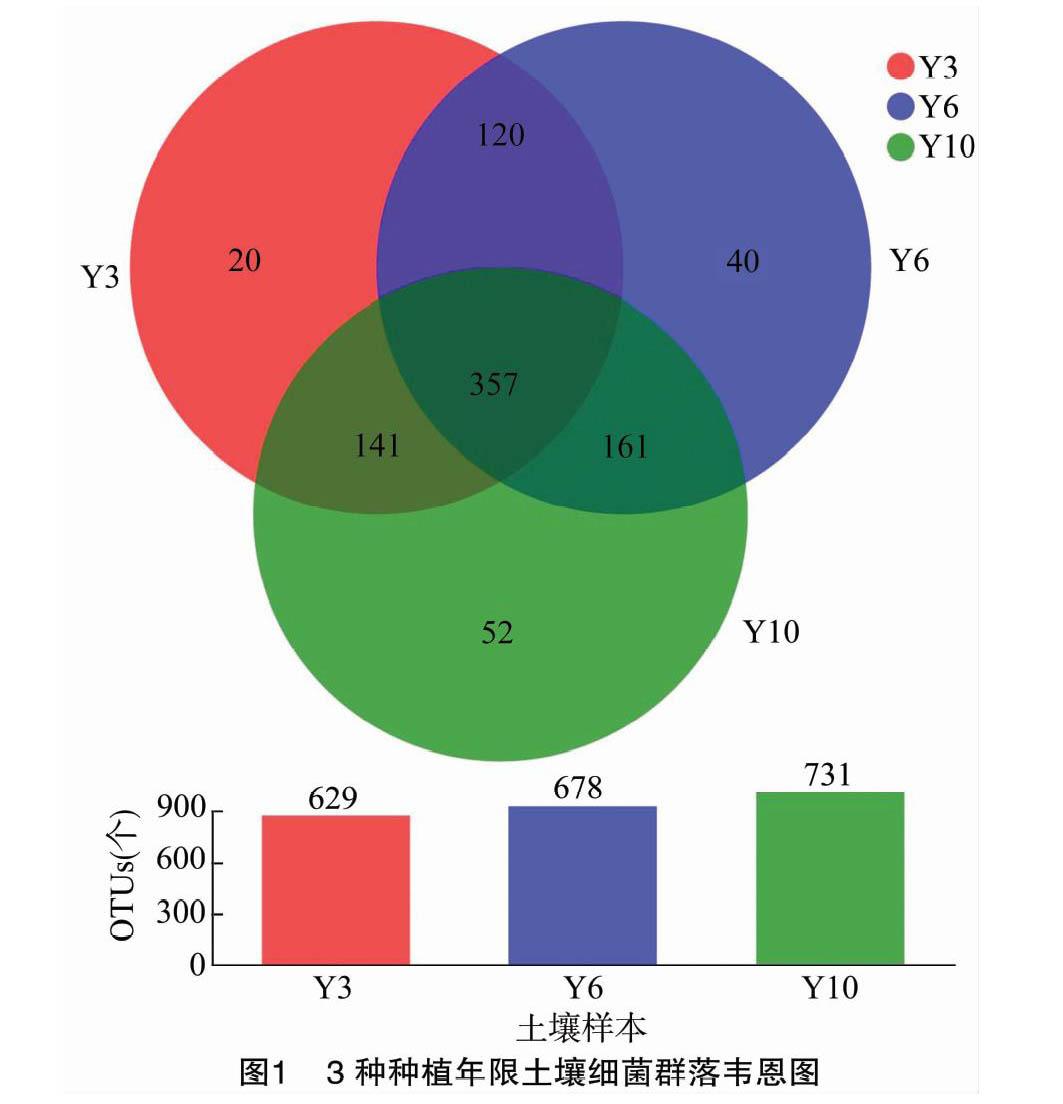
OTU 是便于分析的假定分类单位,本试验将相似性高于97%的有效序列归为一类,9个样本共获得891个OTUs,其中移栽后3年生牡丹根际土壤样本中有629 个OTUs,6年生牡丹根际土壤样本中有678 个OTUs,10 年生牡丹根际土壤样本中有731个OTUs (图1)。从韦恩图(图1)可以看出,不同年限共有OTU 357个,占总数的40.1%;Y3独有的OTU数量为20个,占总数的2.2%;Y6独有的OTU数量为40个,占总数的4.5%;Y10独有的OTU数量为52个,占总数的5.8%。
2.3 不同种植年限油用凤丹牡丹根际土壤真菌物种丰度聚类
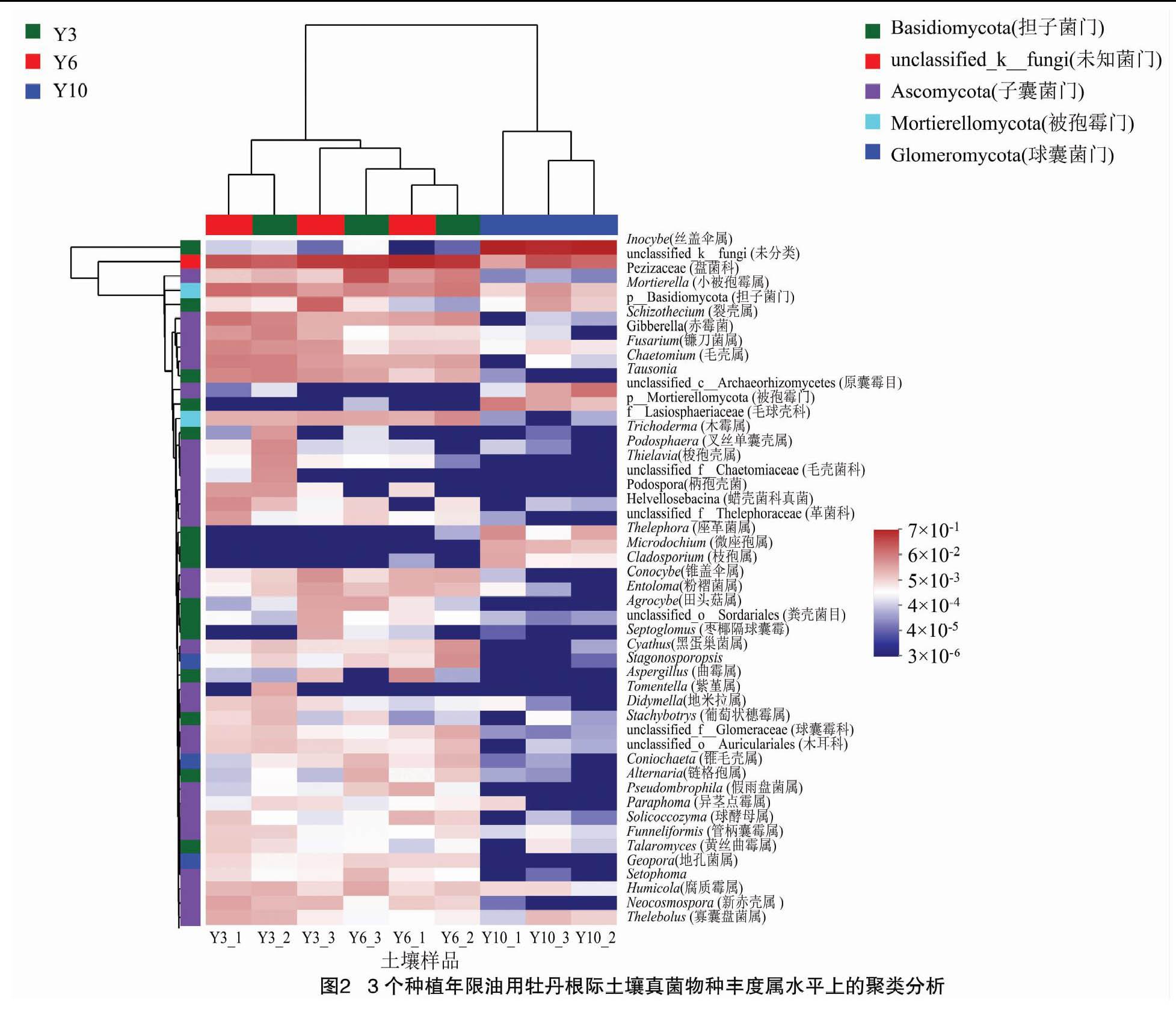
依照所有样本在科级的物种注释和丰度信息,选取每个样本中丰度最高的25科及其丰度信息来绘制热图。依据分类信息以及各样本间的差异程度来聚类,并找出样本中聚集性更强的物种(图2)。研究结果显示,在同一种植年限内,凤丹和牡丹的根际真菌样品几乎都汇集一块,由此可见样品间的重复性较好。其中,10年生凤牡丹的3个样本集中到一个分支,与其他年份的分支距离较远,表明它们之间的菌群丰度存在显著差异。此外,从整体上来看,土壤真菌菌群丰度随种植年限的增加而减少,可见不同种植年限的凤丹牡丹根际真菌菌群多样性之间存在差异。
2.4 不同种植年限油用凤丹牡丹根际土壤真菌群落结构组成
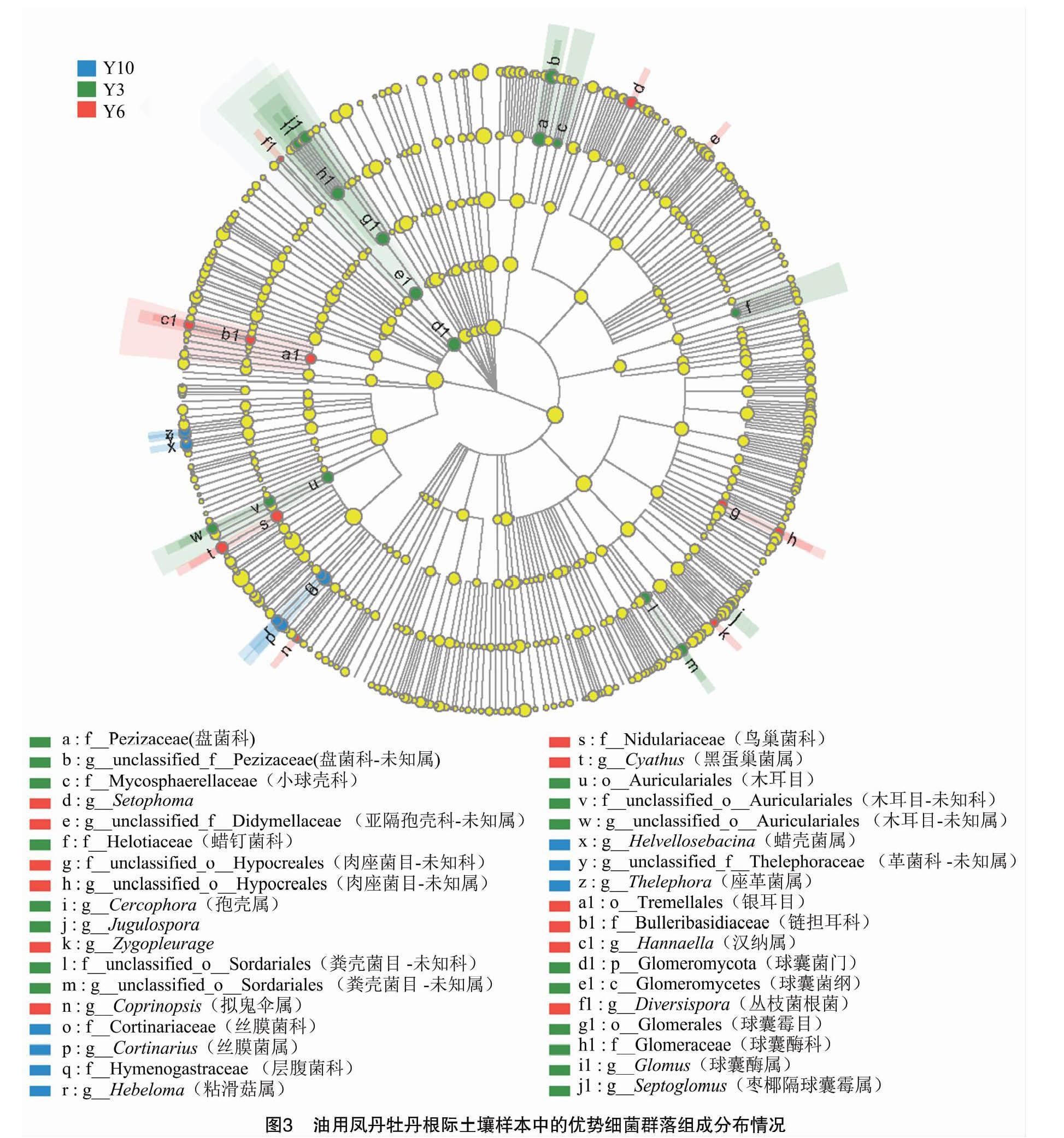
15 个不同种植年限油用牡丹根际土壤样本的真菌群落微生物多样性的整体分布结果显示: 3年生凤丹牡丹的优势真菌群落主要由球囊菌门(Glomeromycota)、木耳科(Auriculariales)、盘菌科(Pezizaceae)、球腔菌科(Mycosphaerellaceae) 等门类构成(图3);6年生凤丹牡丹的优势真菌群落主要有肉座菌目(Hypocreales)、鬼伞属(Coprinopsis)、巢菌科(Nidulariaceae)、黑蛋巢菌属(Cyathus)、银耳目(Tremellales);10年生凤丹牡丹的优势真菌群落主要有丝膜菌属(Cortinarius)、蜡壳菌(Helvellosebacina)、座革菌属(Thelephora)。这说明不同栽培年限的凤丹牡丹真菌的优势群落差异显著,长期栽培会导致土壤真菌组成发生显著改变。
由图4可知,中心优势菌种以担子菌门(45.8%)、子囊菌门(35.5%)、被孢霉门(12.4%)、球囊菌门(9.5%)为主。其中,3、6年生凤丹土壤真菌门主要以子囊菌门和提子菌门为主,而10年生土壤真菌门主要以担子菌门為主,另外3年生土壤真菌的球囊菌门要显著高于种植6、10年生期凤丹土壤。
3个种植年限凤丹牡丹根际真菌群落在属水平上的组成和优势菌属所占比例差异显著,去除未分类的真菌属,所占比例较高的主要隶属于10个真菌属,所占比例达到40%以上。其中丝盖伞属(Inocybe)、被孢霉属(Mortierella)、裂壳属(Schizothecium)、Tausonia、镰刀菌属(Fusarium)、赤霉菌属(Gibberella)为属水平上的优势菌(图5)。
2.5 不同种植年限油用凤丹牡丹根际土壤真菌群落多样性
α多样性是反映丰富度和均匀度的综合指标,其中,香农指数(Shannon指数)可对群落物种组成的均匀度进行评价,物种数(Sobs)和赵氏指数 (Chao1指数)可以很好地反映群落物种的丰富度。3个种植年限的油用凤丹牡丹根际土壤样本中的α多样性指数均出现显著差异。其中,从图6可以看出,土壤真菌Ace、Chao1、Sobs丰富度指数均表现为6年生与10年生之间差异显著,而土壤真菌Shannon多样性指数表现为3年生与10年生、6年生与10年生差异显著。10年生的凤丹土壤真菌Ace、Chao1、Sobs丰富度指数最小,而6年生最高;Shannon多样性指数则表现为3年生最高、10年生最低(图6)。
由图7的主成分分析可看出,PC1和PC2分别解释方差为土壤真菌群落组成的52.65%和12.54%,共解释65.19%。Adonis分析显示,不同种植年限土壤真菌群落结构差异显著(R2=0.882 5,P<0.05)。在PC1轴上可以看出,3年生和6年生土壤真菌群落结构相似度较高,而与10年生的土壤真菌差异显著。
3 讨论
连作障碍通常指某一植物或近缘植物在相同的地域上连续种植多年后,表现为生长状况差、病虫害严重、减产、品质差的现象[16]。连作障碍引起产量下降的重要原因是随着种植年限的增加,植物根际分泌物和植物残体逐年积累,导致土壤微生物区系紊乱[17]。根际微生态系统失衡会刺激土传病原菌大量繁殖,还会对有益拮抗剂的生长起抑制作用,导致植物生长状况差,从而对产量和品质的形成产生影响[18]。
根际是指植物根系周围的土壤区域,该部分土壤受到根系分泌物的施加作用较小,其理化性质和生物活性与原土体有显著差异[19]。在根系分泌物的影响下,该区域微生物的功能结构也会有所变化,引起牡丹连作障碍形成的主要是由于根际微生物群落的结构失调[20]。牡丹根系所分泌出的物质会影响微生物的生长代谢,从而影响根系土壤的理化性质,对根系微生物种群结构产生影响[15]。
在種植多年的牡丹区域中再栽培牡丹植株时,牡丹种苗在生长发育阶段会出现生长效率逐年减慢,根系和植株生长状况不好、病虫害严重等连作障碍所引起的现象。如今连作障碍对牡丹产业的成长和进化影响重大,具有极其复杂的形成过程和加重机制,尤其在根际微生物群落结构方面最为重要[21]。
随着分子生态学技术的蓬勃发展,对植物根际真菌区系结构的研究技术也随之发展,如克隆测序技术、变性梯度电泳、温度梯度电泳、高通量测序技术和扩增18S rDNA 限制性分析。在本研究中共得到1 020 520条优质序列,这些序列足以代表样本中的凤丹牡丹根际土壤菌群,从而有效地反映出其微生物种类和结构。对本试验中的数据分析后可得到891个OTUs,涵盖了5 门、115纲、15目、175 科、230属、320种菌群。对不同种植年限凤丹牡丹根际菌群群落丰度对比分析可看出,根际效应在不同的真菌中有明显的差异。随着土壤中的有益微生物数量的不断减少,某些病原菌数量也因此逐年增加,则导致连作状态下的凤丹牡丹生长发育速度逐渐减慢。
物种多样性为群落的一个重要特征,而对比2个群落中的某物种的丰富度指数是比较群落物种多样性最便捷的方法。但是,物种多样性具有二元性,物种丰富度可能会产生误导,因此常用综合指标——多样性指数来衡量物种多样性。多样性指数常由多种计算方式所得出,其中物种丰富度、香农指数、辛普森多样性均是计算多样性指数最常使用的方法。目前,牡丹根际微生物的相关研究结果均显示,牡丹根际真菌多样性会由于种植年限的过长而降低。在本研究中,4种指标的结果都呈现抛物线的变化趋势,即随着种植年限的增加,根际菌群多样性先升高后降低,其中在连作10年时的多样性水平最高 (图7),4种指标的结果趋势与OTUs 一致。由此可见,连作会影响凤丹牡丹植株和其根际微生物之间的作用效果,改变有益菌和病原菌在土壤中占比,从而引起失衡。
本试验运用Illumina 高通量测序技术,对3个种植年限的凤丹牡丹根际真菌群落结构及相对丰度的变化规律进行探讨。结果表明,在连作状态下,凤丹牡丹根际土壤中菌群群落会发生改变,其中真菌数量和群落多样性指数均随着种植年限的延长呈现先升高后降低的趋势;根际土壤中的菌群结构出现变化,不同种植年限凤丹牡丹土壤中的微生物菌群都拥有高特异性,但关键优质菌群几乎没有变化;在不同分类群中发现了不同种植年限的优势菌群,但这些菌株的大量存在引起的根际微环境的变化仍需进一步探讨。
4 结论
凤丹牡丹长期的连作种植会导致土壤真菌丰富度和均匀度发生显著改变,特别是在种植10年的土壤中真菌丰富度和均匀度均显著下降,这说明凤丹的长期连作会对土壤真菌α多样性产生不利的影响。而连作10年以上会造成土壤真菌β多样性也发生显著改变。
油用牡丹根际土壤样本的真菌群落主要包括球囊菌门(Glomeromycota)、木耳科(Auriculariales)、盘菌科(Pezizaceae)、球腔菌科(Mycosphaerellaceae)、肉座菌目(Hypocreales)、鬼伞属(Coprinopsis)、巢菌科(Nidulariaceae)、黑蛋巢菌(Cyathus)、银耳目(Tremellales)、丝膜菌属(Cortinarius)、蜡皮马鞍蜡壳菌(Helvellosebacina)、革菌属(Thelephora)。这说明不同栽培年限的凤丹牡丹真菌的优势群落差异显著,长期的栽培会导致土壤真菌组成发生显著改变。
可见,油用凤丹牡丹根际微生物对土壤根际微环境十分重要。根际真菌的多样性和根际分泌物对根际微生物所产生的影响有待探讨。本研究结果可为解决牡丹连作障碍提供一定的数据支撑。
参考文献:
[1]廉小芳,李昱莹,张婉青,等. 凤丹牡丹胚培养体系建立及初代畸形苗甲基化变异分析[J]. 河南农业科学,2020,49 (11):110-119.
[2]李超琼,王雪芹,刘红占,等. 凤丹牡丹PoFAD7基因的克隆及表达分析[J]. 中国油脂,2019,44(9):114-118,127.
[3]王崑仑,赵修华,祖元刚,等. 白藜芦醇在几种植物中各部位的分布[J]. 植物研究,2015,35(4):638-640.
[4]李 娟,潘 婷,张志浩,等. 凤丹牡丹光合特性年变化及其与生理生态因子的关系[J]. 江苏农业科学,2019,47(2):135-137.
[5]戚军超,周海梅,马锦琦,等. 牡丹籽油化学成分GC-MS分析[J]. 粮食与油脂,2005,18(11):22-23.
[6]李子璇,秦公伟,何建华,等. 紫斑牡丹种仁种皮中脂肪酸组成比较分析[J]. 种子,2010,29(1):34-36.
[7]史冬燕,彭恒辰,孙 迅. 牡丹连作下不同种植模式对土壤微生物活性的影响[J]. 菏泽学院学报,2018,40(5):82-85.
[8]Jumpponen A,Jones K L. Vertical distribution of fungal communities in tallgrass prairie soil[J]. Mycologia,2010,102(5):1027-1041.
[9]Schmalenberger A,Tebbe C C. Bacterial diversity in maize rhizospheres:Conclusions on the use of genetic profiles based on PCR-amplified partial small subunit rRNA genes in ecological studies[J]. Molecular Ecology,2002,12(1):251-262.
[10]Akyol ,Ince O,Ince B. Crop-based composting of lignocellulosic digestates:Focus on bacterial and fungal diversity[J]. Bioresource Technology,2019,288:121549.
[11]Xu W,Gao Y H,Gong L F,et al. Fungal diversity in the deep-sea hadal sediments of the yap trench by cultivation and high throughput sequencing methods based on ITS rRNA gene[J]. Deep-Sea Research,2019,145(3):125-136.
[12]郭麗丽,尹伟伦,郭大龙,等. 油用凤丹牡丹不同种植时间根际细菌群落多样性变化[J]. 林业科学,2017,53(11):131-141.
[13]韩继刚,王云山,胡永红. 拮抗多种牡丹病原菌的多黏类芽孢杆菌的应用:CN105861378A[P]. 2016-08-17.
[14]郑 艳,刘 炜,黄军祥,等.基于牡丹根际土壤微生物的中药材道地性研究[J]. 药学学报,2016,51(8):1325-1333.
[15]史冬燕,王宜磊. 牡丹根际微生物区系及土壤酶活的研究[J]. 黑龙江农业科学,2013 (6):15-17.
[16]Xiao X M,Cheng Z H,Meng M A,et al. Intercropping with garlic alleviated continuous cropping obstacle of cucumber in plastic tunnel[J]. Acta Agriculturae Scandinavica Section B(Soil and Plant Science),2012,62(8):696-705.
[17]Cheng F,Ali M,Liu C,et al. Garlic allelochemical diallyl disulfide alleviates autotoxicity in the root exudates caused by long-term continuous cropping of tomato[J]. Journal of Agricultural and Food Chemistry,2020,68(42):11684-11693.
[18]Chandanie W A,Kubota M,Hyakumachi M. Interactions between the arbuscular mycorrhizal fungus Glomus mosseae and plant growth-promoting fungi and their significance for enhancing plant growth and suppressing damping-off of cucumber (Cucumis sativus L.)[J]. Applied Soil Ecology,2009,41(3):336-341.
[19]Liu H,Wu M,Liu J M,et al. Tripartite interactions between endophytic fungi,arbuscular mycorrhizal fungi,and Leymus chinensis[J]. Microbial Ecology,2020,79(1):98-109.
[20]Xiao X M,Cheng Z H,Lü J,et al. A green garlic (Allium sativum L.) based intercropping system reduces the strain of continuous monocropping in cucumber (Cucumis sativus L.) by adjusting the micro-ecological environment of soil[J]. Peer J,2019,15:e7267.
[21]马会萍,彭正锋. 牡丹重茬种植障碍产生的原因及对策[J]. 中国园艺文摘,2011,27(4):122-123.
猜你喜欢
热带农业科学(2016年11期)2017-01-21
热带农业科学(2016年11期)2017-01-21
求知导刊(2016年30期)2016-12-03
戏剧之家(2016年22期)2016-11-30
科技资讯(2016年19期)2016-11-15
人间(2016年27期)2016-11-11
农业与技术(2016年15期)2016-11-09
河北渔业(2015年10期)2015-10-15
河北渔业(2015年7期)2015-07-21
河北渔业(2015年6期)2015-06-24
