
不同种源桤木嫁接幼苗光合和叶绿素荧光特征
2021-01-18 08:27白文玉铁烈华冯茂松戴晓康高嘉翔
四川农业大学学报 2020年6期
白文玉,铁烈华,冯茂松,赖 娟,戴晓康,高嘉翔
(四川农业大学林学院/长江上游森林资源保育与生态安全国家林业和草原局重点实验室/长江上游林业生态工程四川省重点实验室,成都 611130)
桤木(Alnus cremastogyne)为桦木科(Betulaceae)桤木属(Alnus)落叶乔木,是我国西南地区的速生用材树种和四川省重要的乡土树种,其根系发达,富含根瘤,具有较强的固氮能力,能有效地改良土壤,已成为我国长江防护林工程及国家造林项目的重要树种[1]。因其适应性强,木材用途广泛,被列为优质造纸材树种,具有广阔的发展前景[2]。
光合作用是植物生长发育的物质基础[3],是决定植物生产力的重要生理过程[4]。叶绿素荧光在探测植物对光能的利用能力以及对环境的适应能力等方面具有重要作用,可以反映光合机构对光能的吸收、转换以及电子传递过程,被视为研究植物光合作用与环境关系的内在探针[5-6]。光合特征参数和叶绿素荧光动力学参数是衡量植物光合作用能力和环境适应能力的重要指标,在植物的生长发育[7-9],逆境生理[10-11]和良种选育[12-14]等研究中均有广泛的应用。通过对植物光合与叶绿素荧光的研究可以揭示植物对其生存环境的生态适应性机制。
不同种源桤木长期处于不同的生境中,由于遗传变异和自然选择的结果,其生长情况、光合生理特征和形态结构等可能发生变化。由于内在遗传因素的改变,不同种源或不同无性系之间可能存在许多差异,这些差异可能会体现在生长特征、光合特征和叶绿素荧光特征等方面。因此,本次试验通过对1年生不同种源桤木嫁接幼苗光合和叶绿素荧光特征的研究,结合嫁接幼苗的生长状况,选出适宜地区发展的优良种源,为建立桤木无性系初级种子园和国家级桤木良种基地做准备,对加快长江上游生态建设具有重要的意义。
1 材料和方法
1.1 试验地概况
本试验地位于巴中市平昌县元山镇(31°37'18.6''N,107°7'57.4''E),属于中亚热带湿润季风气候,海拔为650 m,年降雨量为1 213 mm。四季分明,夏季平均温度30℃,8月份最高可达40℃,年平均日照时数1 239.3 h。土壤为紫色土,土层深约40 cm。
1.2 试验材料
本次试验材料来源于平昌县元山镇四川省桤木良种育苗基地。基地具有36个1.2 m×6 m的苗圃小区,采用1年生长势基本一致、无病虫害的当年生桤木实生营养袋苗作为砧木,砧木株行距为20 cm×30 cm。根据前期积累的桤木选优、嫁接和田间管理等经验[15]。2016年1月初,采用随机区组设计方法,以15~20年恩阳、金堂、平昌和宣汉种源桤木优树穗条作为接穗进行嫁接,同株优树上采集的穗条分别嫁接于不同小区。
1.3 试验方法
2016年8月中旬,首先测定小区内不同种源所有嫁接幼苗的株高和基径,根据测定结果的平均值,在每小区内选出3株嫁接幼苗作为待测植株,测定其光合参数和叶绿素荧光参数。
1.3.1 光合参数的测定
于晴朗天气,选取待测植株中上部完全成熟的叶片,用便携式光合作用仪 Li-6800(Li-Cor Inc.,USA)自带红蓝光源。测定时设置光照强度为1 200 μmol/(m2·s),控制CO2浓度为400μmol/mol、温度为25℃、相对湿度为45%~65%。测定其净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间 CO2浓度(Ci)。
快速光响应曲线的测定:对光适应样品进行连续光量子能量光照射,光合有效辐射PAR梯度为0、25、50、75、100、200、400、600、800、1 000、1 200、1 400、1 600和1 800 μmol/(m2·s),从而测得电子传递速率(ETR)。测定时设定温度为25℃,空气湿度为45%~65%。
1.3.2 叶绿素荧光的测定
叶片选取同1.3.1节。
叶绿素荧光参数的测定:采用Li-6800自带荧光叶室测定植株的叶绿素荧光参数。用锡箔纸裹住待测叶片,待叶片暗适应50 min后,首先用一束小于0.1 μmol/(m2·s)的测量光(频率为50 Hz)照射,得到初始荧光F0;后打开一个饱和脉冲光(8 000 μmol/m2·s),10 Hz,1 000 ms),得到暗适应下的最大荧光Fm,然后根据环境光强设定活化光,待F稳定后,此时的F即为稳态荧光Fs;然后再打开一个同样强度的饱和脉冲光,得到Fm';同样打开远红光(25 μmol/m2·s)得到F0'。根据测定数据,可计算得出PSⅡ潜在活性(Fv/F0)、PSⅡ最大光化学量子产量(Fv/Fm)、PSⅡ实际光化学量子产量(φPSⅡ)、光化学淬灭系数(qP)和非光化学淬灭系数(qN)[16]。
1.4 数据处理
采用直角双曲线修正模型拟合快速光响应曲线,模型表达式如下[17]:
式中:ETR为电子传递速率,a快速光响应曲线的初始斜率,PAR为光合有效辐射,b和c为系数。
用 Excel 2007 和 SPSS 19.0(SPSS Inc,USA)进行数据统计与分析。用采用单因素方差分析方法(oneway ANOVA)比较不同种源桤木嫁接幼苗间光合参数和叶绿素荧光参数的差异。并用皮尔森相关系数(Pearson correlation coefficient)分析光合参数与荧光参数之间的相关性。显著性水平设定为a=0.05。
2 结果与分析
2.1 不同种源桤木光合特征参数
由表1可知,4个种源的净光合速率(Pn)和胞间 CO2浓度(Ci)均存在显著(P<0.05)差异。其中平昌种源桤木的净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr)均最大,且显著(P<0.05)高于其他种源。金堂种源的净光合速率(Pn)和气孔导度(Gs)均显著(P<0.05)高于恩阳和宣汉,同时金堂种源的蒸腾速率(Tr)与恩阳差异不显著(P>0.05),与宣汉存在显著差异(P<0.05)。恩阳、宣汉的气孔导度(Gs)和蒸腾速率(Tr)均差异不显著(P>0.05)。宣汉种源的胞间CO2浓度(Ci)最大,显著(P<0.05)高于其他种源,平昌种源的胞间CO2浓度(Ci)最小。
2.2 不同种源桤木快速光响应曲线特征
由快速光响应曲线可以拟合出最大电子传递速率。由图1可知,4个种源的最大电子传递速率(ETRmax)表现为平昌>金堂>恩阳>宣汉。其中,平昌种源的最大电子传递速率(ETRmax)显著(P<0.05)高于其他种源,金堂种源的ETRmax次之,宣汉种源的ETRmax最小,仅为平昌种源的65%。
快速光响应曲线反映了植物叶片的电子传递速率(ETR)随光合有效辐射(PAR)强度改变的变化规律。由图2可知,ETR随PAR呈又快到慢的增长特征,在PAR小于200 μmol/(m2·s)的弱光环境下,各种源桤木的ETR迅速上升;当400 μmol/(m2·s)<PAR<1 600 μmol/(m2·s)时,随光照强度的增加,桤木的ETR增长速度明显放缓,不同种源桤木嫁接幼苗的ETR随PAR的增长速率出现较大差异,其中宣汉种源桤木嫁接幼苗的ETR随PAR的增长速率最慢,平昌种源桤木嫁接幼苗的ETR随PAR的增长速率相对较快;当PAR超过1 600 μmol/(m2·s)后,各种源桤木嫁接幼苗快速光响应曲线趋近平稳。
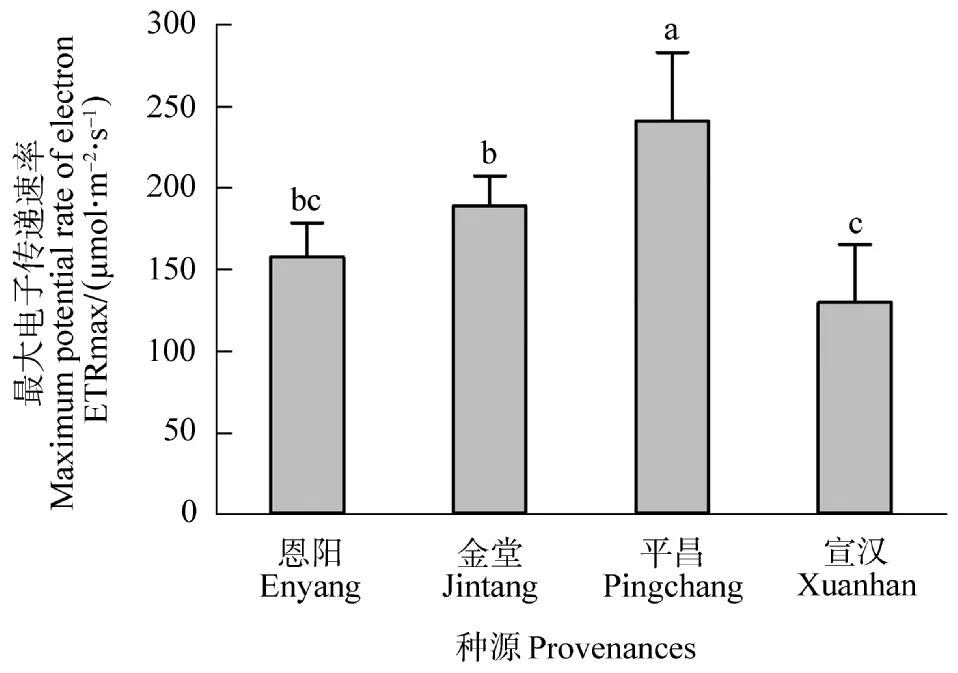
图1 不同种源桤木最大电子传递速率Figure 1 The ETRmaxof different provenances for the A.cremastogyne
2.3 不同种源桤木荧光产量特征
由图3可知,4个种源间的初始荧光产量(F0)存在显著(P<0.05)差异。其中,宣汉初始荧光产量(F0)显著(P<0.05)高于其他种源。恩阳、金堂初始荧光产量(F0)次之,平昌种源的初始荧光产量(F0)最低。4个种源间的最大荧光(Fm)产量差异不显著(P>0.05),表现为平昌>金堂>宣汉>恩阳。平昌种源的可变荧光(Fv)产量最高,且与金堂种源差异不显著(P>0.05),与恩阳、宣汉存在显著(P<0.05)差异。恩阳、宣汉种源的可变荧光(Fv)产量差异不显著(P>0.05)。可见,不同种源间的PSII反应中心活性存在一定的差异。
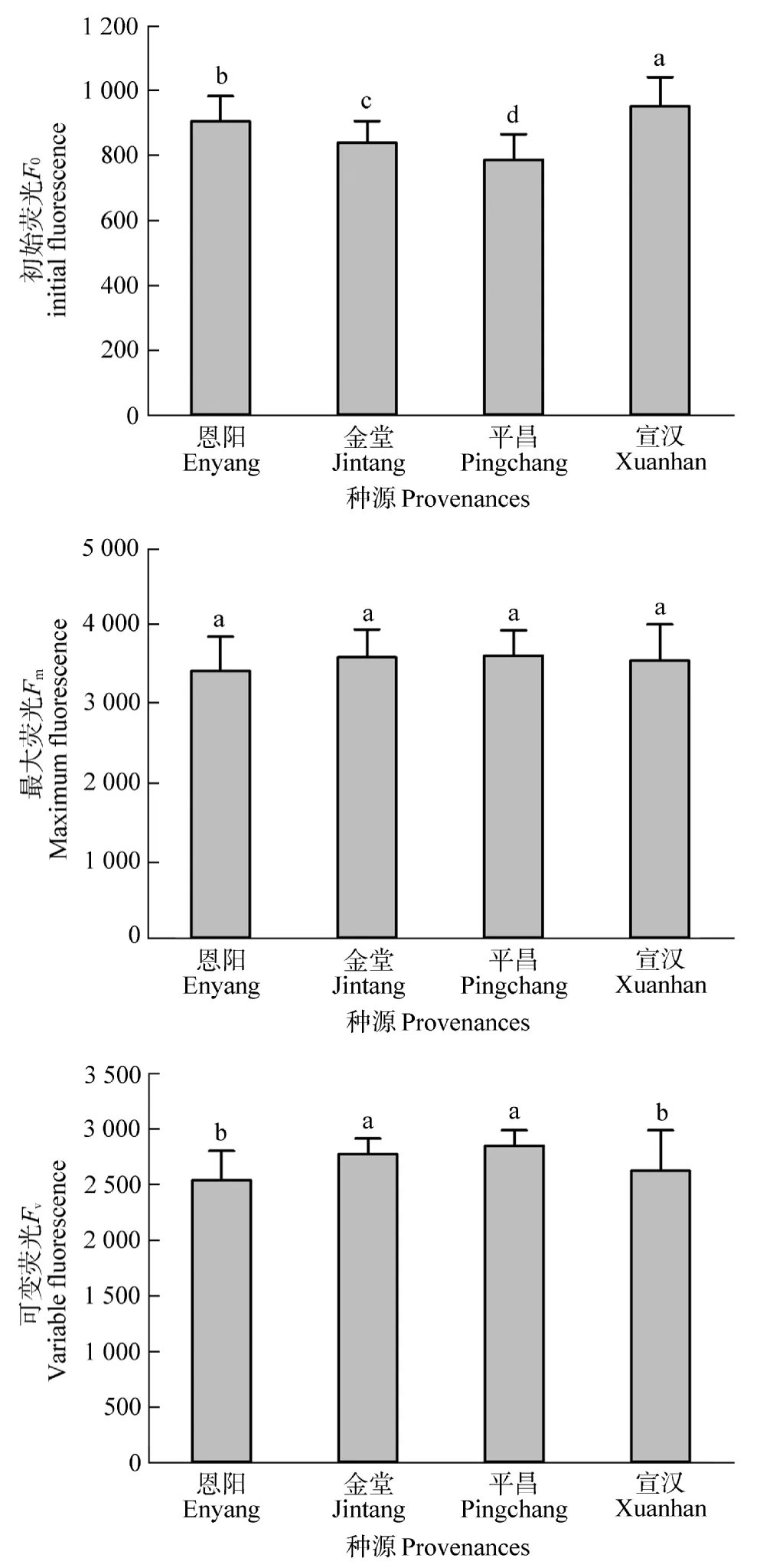
图3 不同种源桤木荧光产量特征Figure 3 The fluorescence yield characteristics of different provenances for the A.cremastogyne
2.4 不同种源桤木光能转换效率特征
由图4可知,平昌种源的最大光化学效率(Fv/Fm)值最大,与恩阳、宣汉存在显著(P<0.05)差异,与金堂种源差异不显著(P>0.05)。此外,平昌种源的潜在化学效率(Fv/F0)值也最大,与恩阳、金堂、宣汉均存在显著(P<0.05)差异,恩阳和宣汉种源的潜在化学效率(Fv/F0)差异不显著(P>0.05)。4个种源间的实际光化学效率(φPSⅡ)均存在显著差异(P<0.05),表现为平昌>金堂>恩阳>宣汉。其中,平昌种源的实际光化学效率(φPSⅡ)显著(P<0.05)高于其他种源,宣汉种源的实际光化学效率(φPSⅡ)显著(P<0.05)低于其他种源,仅为平昌种源的56.7%。可见,不同种源间的光能吸收和转换能力存在一定的差异。

图4 不同种源桤木光能转换效率特征Figure 4 The energy conversion efficiency characteristics of different provenances for the A.cremastogyne
2.5 不同种源桤木淬灭系数特征
由图5可知,4个种源间的光化学淬灭系数(qP)差异显著(P<0.05),其中平昌种源的光化学淬灭系数(qP)值最高,金堂、恩阳次之,宣汉种源的光化学淬灭系数(qP)值最低。平昌种源的非光化学淬灭系(qN)值最高,与金堂种源差异不显著(P>0.05),均显著(P<0.05)高于恩阳、宣汉。恩阳与宣汉种源的非光化学淬灭系(qN)存在显著(P<0.05)差异,且宣汉种源的非光化学淬灭系(qN)值最低。
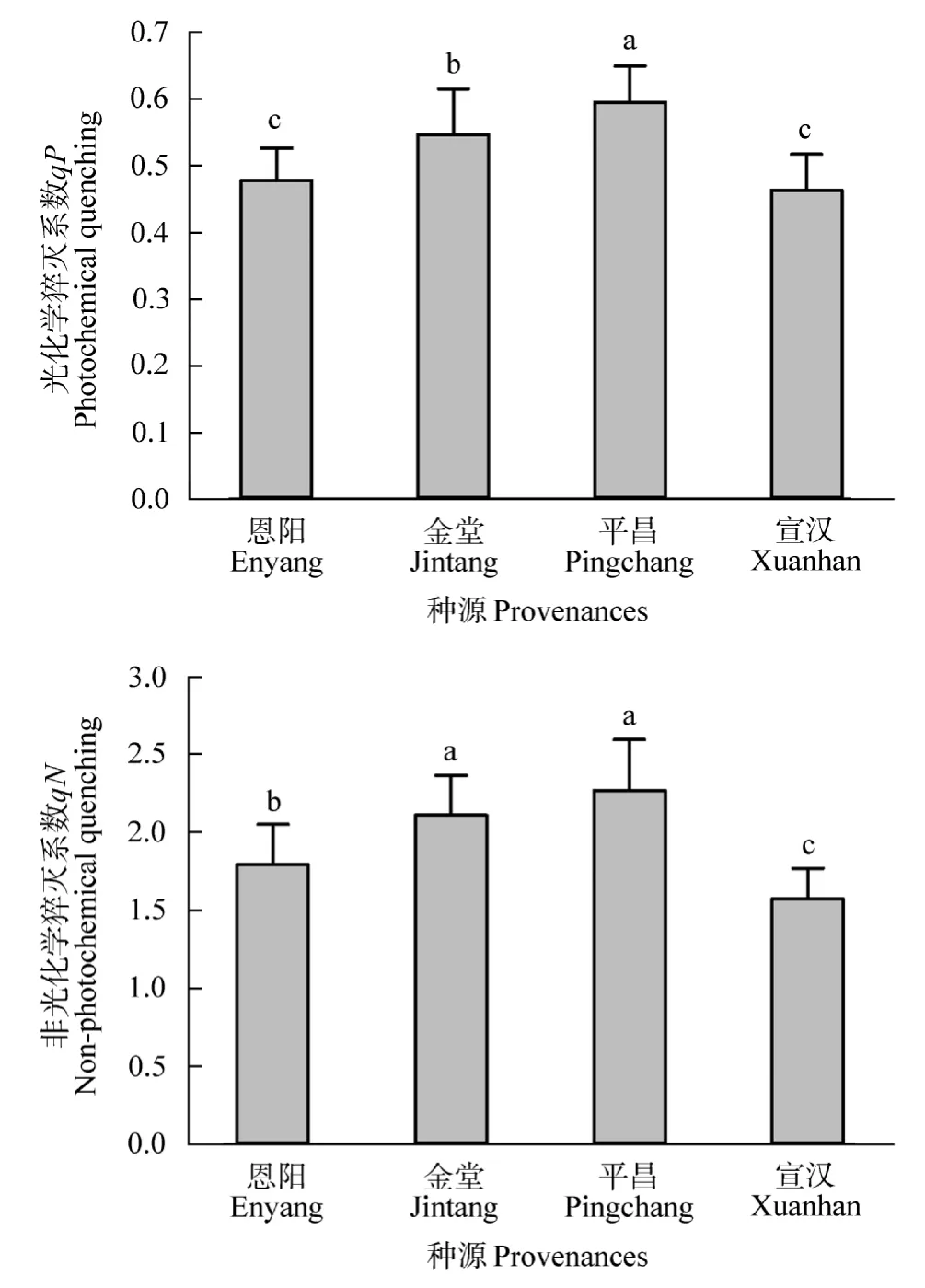
图5 不同种源桤木淬灭系数特征Figure 5 The quenching coefficient characteristics of different provenances for the A.cremastogyne
2.6 桤木光合特征参数与叶绿素荧光参数的相关关系
由表 2 可知,初始荧光(F0)与净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr)呈极显著(P<0.01)负相关关系,与胞间 CO2浓度(Ci)极显著(P<0.01)正相关关系。最大光化学效率(Fv/Fm)、潜在化学效率(Fv/F0)、实际光化学效率(φPSⅡ)、光化学淬灭系数(qP)以及非光化学淬灭系数(qN)均与净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr)呈极显著(P<0.01)正相关关系,与胞间 CO2浓度(Ci)极显著(P<0.01)负相关关系。最大荧光(Fm)与净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间 CO2浓度(Ci)均不存在显著(P>0.05)关系。
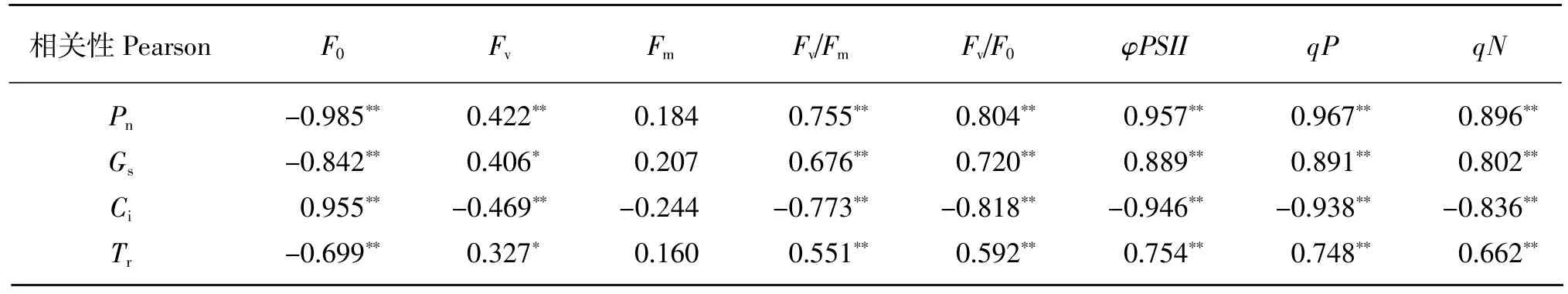
表2 桤木光合特征参数与叶绿素荧光参数之间的相关性Table 2 Correlation between photosynthetic characteristics and chlorophyll fluorescence parameters of A.cremastogyne
3 讨论
光合速率是植物生长和外界环境综合作用的象征[18]。本研究表明,平昌种源桤木嫁接幼苗的净光合速率(Pn)和蒸腾速率(Tr)最高,气孔开放程度最大,叶片中CO2的供应充足;而宣汉种源的净光合速率(Pn)和蒸腾速率(Tr)最低,气孔开放程度最小。可见,平昌种源固定环境中CO2能力较强,净光合速率最高,通过光合作用合成有机产物较多;宣汉种源光合作用能力则最弱,合成的有机产物较少。这可能与不同种源的光化学淬灭系数(qP)存在差异有关。光化学淬灭系数(qP)是PSⅡ天线色素吸收的光能用于光化学反应电子传递的份额,可以反映出PSⅡ反应中心的开放程度,其值越大,PSⅡ反应中心的电子传递活动就越活跃[19]。本研究中,平昌和金堂种源桤木嫁接幼苗的光化学淬灭系数(qP)差异不显著,且显著大于恩阳、宣汉种源。可见,平昌和金堂种源桤木嫁接幼苗PSⅡ反应中心的开放程度可能较高,植物光合电子传递的能力较强,因此净光合速率较高。而恩阳、宣汉种源桤木嫁接幼苗PSⅡ反应中心的开放程度较低,因此净光合速率较低。此外,胞间CO2浓度(Ci)的大小与植物叶片气孔开闭程度和环境因素(如光照、温度、CO2浓度等因素)有关。在本研究中4个种源桤木所处环境相同,因此,胞间CO2浓度的差异可能主要是由气孔导度(Gs)差异造成。这与刘落鱼等研究结果相似[20]。
叶绿素荧光动力学参数可以反映出环境对植物光合生理作用的影响[21]。初始荧光(F0)表示PSⅡ反应中心处于完全开放时的荧光产量。有研究表明,当环境胁迫时初始荧光(F0)一般呈现升高趋势,且伴随内囊体膜结构变化和PSⅡ反应中心可逆失活,保护PSⅡ反应中心免遭光破坏[22-24]。本研究中各种源初始荧光(F0)值偏高。出现这种现象可能是因为,本次调查在8月中旬,温度较高、光照较强,高温和强光等环境因素可能对桤木嫁接幼苗产生了一定程度的胁迫,桤木嫁接幼苗为了保护PSⅡ反应中心免遭光破坏,从而提高初始荧光(F0)。这与B.T.Nguyen等[25]对越南杉木的研究结果相似。同时,初始荧光(F0)与气孔导度(Gs)和蒸腾速率(Tr)负相关(见表2)也进一步支持了的桤木嫁接幼苗经受了环境胁迫的观点。我们的结果还表明,平昌种源桤木嫁接幼苗初始荧光(F0)显著低于其他种源,而宣汉种源桤木嫁接幼苗初始荧光(F0)显著高于其他种源。此外,最大荧光(Fm)可以有效地反映通过PSⅡ反应中心最大的电子传递情况[26]。我们发现,4个种源桤木嫁接幼苗的最大荧光(Fm)差异不显著。这表明,4个种源桤木嫁接幼苗通过PSⅡ反应中心传递电子的最大能力相当。同时,最大电子传递速率(ETRmax)对CO2的固定和同化起着重要的作用[27-29]。本研究中,平昌种源桤木嫁接幼苗最大电子传递速率(ETRmax)显著大于其他种源。这表明,虽然4个种源桤木嫁接幼苗通过PSⅡ反应中心传递电子的最大能力相当,但是由于平昌种源桤木嫁接幼苗的PSⅠ和PSⅡ反应中心色素分子数量较多,经PSⅠ和PSⅡ光系统的非环式电子传递速率较高,能较快地将光能转化为电能并传递给NADP+生成NADPH,并驱动ADP磷酸化生成ATP,加快碳同化过程。因此,平昌种源桤木嫁接幼苗固定CO2的能力较强,净光合速率较高。这也可能是因为,平昌种源桤木嫁接幼苗经PSI光系统的环式电子传递速率较高,因此4个种源桤木嫁接幼苗的最大荧光(Fm)差异不显著,但平昌种源最大电子传递速率(ETRmax)显著大于其他种源,较高的环式电子传递速率可以使光合机构免受光破坏。值得注意的是,叶绿素荧光指标测定期间(8月)的每日最高温为38~40℃,光照充足。由此可见,平昌种源桤木嫁接幼苗适应当地夏季高温、高光照的能力较强,而宣汉种源桤木接幼苗受当地夏季高温、高光照胁迫的影响可能较大。
实际光化学效率(φPSⅡ)反映了PSⅡ反应中心在部分关闭情况下的实际原初光能捕获效率,表示光合电子传递能量与吸收光能的比例[30]。本研究中,平昌种源桤木嫁接幼苗的实际光化学效率(φPSⅡ)显著高于其余种源。这表明,平昌种源桤木嫁接幼苗实际传递电子的能力更强。此外,最大光化学效率(Fv/Fm)则反映光能转换效率,是衡量植物光抑制程度的重要指标[31]。正常情况下,植物最大光化学效率(Fv/Fm)在 0.75~0.85[32]。本研究表明,平昌和金堂种源桤木嫁接幼苗的最大光化学效率(Fv/Fm)分别为0.78和0.77。因而,平昌和金堂种源桤木嫁接幼苗可能未出现光抑制现象。但是,恩阳和宣汉种源桤木嫁接幼苗的最大光化学效率(Fv/Fm)均为0.73。这表明,恩阳和宣汉种源桤木嫁接幼苗的PSⅡ反应中心可能受损,出现了光抑制现象。这与李志真等[33]对玉簪(Hosta plantaginea)叶片光抑制的研究结果一致。当植物受到环境胁迫时,植物会启动自我保护机制,将过多的光能转化为热能,并散发出去,降低对叶绿体等光合机构的损伤[34]。需要指出的是,非光化学淬灭系数(qN)表示植物PSⅡ反应中心将光能转化为热能的能力[35]。本研究还发现,平昌和金堂种源桤木嫁接幼苗的非光化学淬灭系数(qN)差异不显著,且显著大于恩阳和宣汉种源。这表明,平昌和金堂种源桤木的光合机构保护能力(抵御强光照和高温环境的能力)大于恩阳和宣汉,PSⅡ反应中心活性较高,能够将过多的光能转化为热能,因此不容易出现光抑制现象,光合速率维持在较高水平。相比之下,恩阳和宣汉种源桤木嫁接幼苗将过剩光能转化为热能的能力较弱,较难以散热形式将过多的光能消耗掉,所以其较容易出现光抑制现象,导致其光合作用能力降低。有趣的是,我们结果表明光合效率高的种源,其非光化学淬灭系数(qN)也相对较高。这与之前代大川[36]对桢楠幼苗的研究结果不一致。这一现象可能是因为,本试验指标测定期间温度较高,桤木嫁接幼苗通过提高非光化学淬灭系数(qN)尽量加快叶片散热,同时相应的增加蒸腾速率(qN与蒸腾速率正相关;表2)来加快散热,增加气孔导度(Gs),从而维持光合机构正常运行。但这还需要进一步研究。
综上所述,4个种源桤木嫁接幼苗在光合特征和叶绿素荧光特征存在显著差异。平昌种源桤木嫁接幼苗光合作用最强,且具有较强的自我保护机制。金堂种源桤木嫁接幼苗光合作用和自我调节机制次之。恩阳、宣汉种源桤木嫁接幼苗光合作用较弱,自我调节能力较差。因此,可根据经营目的选择适合地区发展的桤木嫁接幼苗,也可以根据不同种源桤木的光合和叶绿素荧光特征选择适合其发展的地区。
猜你喜欢
民间文学(2022年1期)2022-03-31
绿色科技(2021年21期)2021-11-26
现代盐化工(2021年3期)2021-08-27
种子(2021年3期)2021-04-12
先锋(2018年5期)2018-10-21
民间文学(2018年3期)2018-04-28
发明与创新·大科技(2017年7期)2017-07-17
福建林业(2016年4期)2016-08-16
新课程·下旬(2015年10期)2015-10-21
缤纷家居(2014年5期)2014-11-03
