
广东苦茶资源嘌呤生物碱含量分析与评价
2021-02-06 06:35李红建秦丹丹姜晓辉方开星王秋霜潘晨东李波王青吴华玲
茶叶科学 2021年1期
李红建,秦丹丹,姜晓辉,方开星,王秋霜,潘晨东,李波,王青,吴华玲
广东苦茶资源嘌呤生物碱含量分析与评价
李红建,秦丹丹,姜晓辉,方开星,王秋霜,潘晨东,李波,王青,吴华玲*
广东省农业科学院茶叶研究所/广东省茶树资源创新利用重点实验室,广东 广州 510640
以13份广东省境内收集的苦茶资源为研究对象,来自不同省份的28份代表性品种(系)为对照,分析苦茶资源中嘌呤生物碱含量特征。结果显示,咖啡碱、可可碱、苦茶碱及生物碱总量均表现出较大的遗传差异,遗传多样性指数()为0.78~1.94,变异系数()为34.89%~191.19%。系统聚类分析将试验材料分为6个类群,类群Ⅰ、Ⅱ、Ⅲ主要为常规品种(系),咖啡碱是其优势嘌呤碱;类群Ⅴ为低咖啡碱的南昆山毛叶茶;类群Ⅳ、Ⅵ全部为苦茶资源,苦茶碱含量较高。通过综合评价,从苦茶资源中筛选出生物碱含量特异的育种材料10份。此外,不同叶位含量分析表明,苦茶中苦茶碱随叶片成熟度增加呈缓慢降低趋势,至成熟叶期仍维持较高水平。研究结果为今后嘌呤生物碱含量特异的功能性苦茶品种培育及开发利用提供了依据。
苦茶;嘌呤生物碱;种质资源;遗传多样性;聚类分析
苦茶作为一类特殊的茶树资源,因其干茶味极苦而得名,早期多以野生茶树形式存在,具有一定的药理作用[1]。苦茶在我国主要分布在粤、湘、赣毗邻区,尤其以南岭山脉两侧居多,在云南红河、西双版纳等地也有分布[2-3]。江华苦茶是研究较多的一类苦茶,研究者认为其与云南大叶茶亲缘关系较近,是由云南大叶演变到灌木型小叶种的一类过渡类型[4-5]。王新超等[1]分析了来自广东、湖南、江西等地苦茶资源的主要生化成分,将苦茶与阿萨姆种聚为一类,并认为苦茶是比较适合制作红茶的资源。上世纪八十年代发现的生长在云南东南部红河州的野生大叶种苦茶[6],被石祥刚等[7]升格为独立的种。
茶叶中嘌呤生物碱是决定茶叶品质的关键成分之一,有多种生理功效[8-12]。常规茶叶中咖啡碱(Caffeine)为优势嘌呤生物碱,约占茶叶干重的2%~5%,可可碱(Theobromine)和茶碱(Theophylline)含量极低,约为0.05%和0.002%[13]。前人对苦茶资源生物碱也进行了较多研究。杨春等[14]对江华苦茶资源生物碱测定发现其咖啡碱和可可碱含量较高;唐琴等[15]对福建尤溪苦茶分析发现,其含有3种生物碱(咖啡碱、可可碱、茶碱),含量稍高于对照,但分布模式与常规茶基本一致;叶创兴等[16-17]报道了云南红河苦茶存在一种新的生物碱分布模式,苦茶碱(Theacrine)是其优势嘌呤碱,含量高达1.29%,其次是咖啡碱和可可碱;杨超等[18]对云南西双版纳州的苦茶特征成分进行研究,发现具有特征性成分的苦茶碱是导致该类苦茶滋味极苦的主要原因。由此可见,不同地区的苦茶在嘌呤生物碱的含量及分布模式上存在较大差异,有待对更多类型的苦茶资源进行进一步发掘。
广东地处热带、亚热带季风气候区,良好的气候和生态系统蕴含了众多独特的茶树资源,如白毛茶、凤凰单丛茶、可可茶、客家小叶种及苦茶资源[19-20]。至今,针对广东境内苦茶资源研究报道较少。本课题组前期对广东省境内苦茶资源进行了广泛调查,并收集了13份苦茶资源。本研究以国内不同茶区代表性品种(系)为对照,通过对13份苦茶资源的嘌呤生物碱进行综合评价,旨在了解不同资源的变异特征及利用潜力,为苦茶资源的开发利用提供理论参考和研究基础。
1 材料与方法
1.1 供试材料
供试材料为保存于“广东省茶树种质资源库”的41份种质资源,包括13份新收集的广东苦茶资源和28份来源于国内各省份代表性对照品种(系)(广东种质14份,浙江种质3份,湖南种质2份,湖北种质2份,云南种质2份,福建种质3份,贵州种质1份,台湾种质1份)。其基本信息如表1。
1.2 嘌呤生物碱测定
1.2.1 茶样制备
参考《茶树种质资源描述规范和数据标准》的方法[21],以一芽二叶标准对41份供试材料采样,微波蒸青5 min,80℃恒温干燥固样后粉碎,制作分析样品;同时取5份资源(K4、K22、K29、K32、K36)同一新梢下的芽头、第一至五嫩叶以及成熟叶制作不同叶位样品。
1.2.2 色谱条件
嘌呤生物碱(咖啡碱、可可碱、茶碱、苦茶碱)含量采用高效液相色谱仪(High performance liquid chromatography,HPLC)进行测定。液相色谱条件:流动相为0.1%甲酸(A相)和100%乙腈(B相),流速为1 mL·min-1,柱温35℃,检测波长为231 nm。梯度洗脱:9~15 min,A相由4%线性升至6%,B相由96%线性降至94%;15~30 min,A相由6%线性升至12%,B相由94%线性降至88%;30~55 min,A相由12%线性升至18%,B相由88%线性降至82%;55~58 min,A相由18%线性降至4%,B相由82%线性升至96%。
1.3 数据分析
2 结果与分析
2.1 嘌呤生物碱的含量与变异分析
由于茶碱仅在少数供试材料(K14、K28、K31、K35、K39、K40、K41)中检出,且含量极低(<0.08 mg·g-1),本研究只讨论咖啡碱、可可碱、苦茶碱分析结果,生物碱总量为咖啡碱、可可碱、苦茶碱含量之和。分析结果如表2所示,生物碱总量的遗传多样性指数()最高,为1.94,其次是咖啡碱、苦茶碱,分别为1.76、1.21,可可碱最低为0.78。不同材料间变异系数存在较大差异:可可碱变异系数最大,为191.19%,变幅为0.23~56.82 mg·g-1;其次是苦茶碱,其变异系数为125.54%,变幅为0.18~15.71 mg·g-1;咖啡碱变异系数为51.09%,变幅为0.66~48.72 mg·g-1;生物碱总量变异系数最小,为34.89%,变幅为6.98~61.29 mg·g-1。
2.2 聚类分析
以咖啡碱、可可碱、苦茶碱及生物碱总量为指标,选用欧氏距离、Ward法进行系统聚类分析,构建聚类树状图。由图1可知,在遗传距离阈值为5时,41份茶树资源可以分为6类,各类群的特征见表3。
类群Ⅰ包含14份种质资源,均为各省育成品种(系),其中广东种质资源8份(K16、K22、K25、K32、K34、K35、K36、K37),湖南种质资源1份(K2),云南种质资源2份(K18、K41),福建种质2份(K26、K33),台湾种质资源1份(K27)。这类种质咖啡碱含量平均值为35.90 mg·g-1,显著高于类群Ⅲ、Ⅳ、Ⅴ、Ⅵ;而可可碱(4.34 mg·g-1)、苦茶碱(0.92 mg·g-1)含量较低。
类群Ⅱ包含5份种质资源,有广东种质4份,其中育成品种(系)3份(K38、K39、K40),苦茶资源1份(K10);贵州种质资源1份(K31)。该类种质咖啡碱平均含量为41.45 mg·g-1,显著高于类群Ⅲ、Ⅳ、Ⅴ、Ⅵ。生物碱总量较高,平均值为54.43 mg·g-1,显著高于类群Ⅰ、Ⅲ、Ⅳ、Ⅵ;可可碱含量中等(9.17 mg·g-1);苦茶碱含量较低(3.81 mg·g-1)。值得注意的是,此类种质中K10苦茶碱含量较高,达12.36 mg·g-1。
类群Ⅲ包含8份种质资源,均为各省育成品种(系),其中广东种质资源1份(K1),浙江种质3份(K3、K24、K30),湖北种质2份(K19、K20),湖南种质资源1份(K15),福建种质资源1份(K21)。该类种质咖啡碱含量平均值为26.11 mg·g-1,显著低于类群Ⅰ、Ⅱ;生物碱总量平均值为28.40 mg·g-1,高于类群Ⅵ,但显著低于其他类群;可可碱含量较低,平均值为1.88 mg·g-1;苦茶碱含量为6个类群中最低,平均值仅为0.41 mg·g-1。总体而言,此类群种质嘌呤生物碱含量表现为中等(28.40 mg·g-1)。
类群Ⅳ包含5份种质资源,全部为苦茶资源(K11、K12、K13、K14、K29)。该类种质苦茶碱含量高达12.29 mg·g-1,显著高于类群Ⅰ、Ⅱ、Ⅲ、Ⅴ;咖啡碱含量表现为中等,平均值为23.57 mg·g-1,显著低于类群Ⅰ、Ⅱ;可可碱含量较低,平均值为3.13 mg·g-1。生物碱总量表现为中等,平均值为38.99 mg·g-1。此类群中,K11苦茶碱含量为15.71 mg·g-1,为所有供试材料中最高。
类群Ⅴ包含2份种质资源,均为广东特有种质南昆山毛叶茶(K4、K5)。此类种质可可碱含量平均值为56.12 mg·g-1,显著高于其他类群;咖啡碱含量平均值仅为0.78 mg·g-1,显著低于其他类群;苦茶碱含量较低,平均值为0.50 mg·g-1。此外,此类群生物碱总量为6个类群中最高,平均值为57.39 mg·g-1。
类群Ⅵ包含7份种质资源,全部是苦茶资源(K6、K7、K8、K9、K17、K23、K28)。此类种质苦茶碱含量较高,平均值为10.44 mg·g-1,显著高于类群Ⅰ、Ⅱ、Ⅲ、Ⅴ;咖啡碱含量较低,平均值为5.40 mg·g-1,显著低于类群Ⅰ、Ⅱ、Ⅲ、Ⅴ;可可碱含量和生物碱总量也较低,平均值分别为0.78 mg·g-1和16.61 mg·g-1。
2.3 嘌呤生物碱含量特异种质的筛选
根据聚类分析结果及各类群生物碱综合表现,在苦茶资源中初步筛选出生物碱含量特异的种质10份(表4)。其中高苦茶碱种质4份(苦茶碱>10 mg·g-1),分别为K11、K13、K14和K29;高苦茶碱和高咖啡碱种质1份(苦茶碱12.36 mg·g-1,咖啡碱34.05 mg·g-1),为K10;高苦茶碱和低咖啡碱种质5份(苦茶碱>10 mg·g-1,咖啡碱<10 mg·g-1),分别为K6、K7、K8、K9和K28。
2.4 不同叶位嘌呤生物碱含量变化分析
为进一步明确嘌呤生物碱在不同叶位间含量变化情况,选取5份资源(K4、K22、K29、K32、K36)的芽头、芽下第一至五叶以及成熟叶进行测定。如图2所示,可可碱、咖啡碱、苦茶碱以及生物碱总量均在芽头中含量最高,并随着叶片成熟度的升高而逐渐降低,成熟叶中含量最低。相比而言,苦茶碱在K29苦茶资源不同叶位降低趋势较为平缓,成熟叶期其含量仍维持在较高水平,表明苦茶碱与其他嘌呤生物碱有着不同的累积特性。
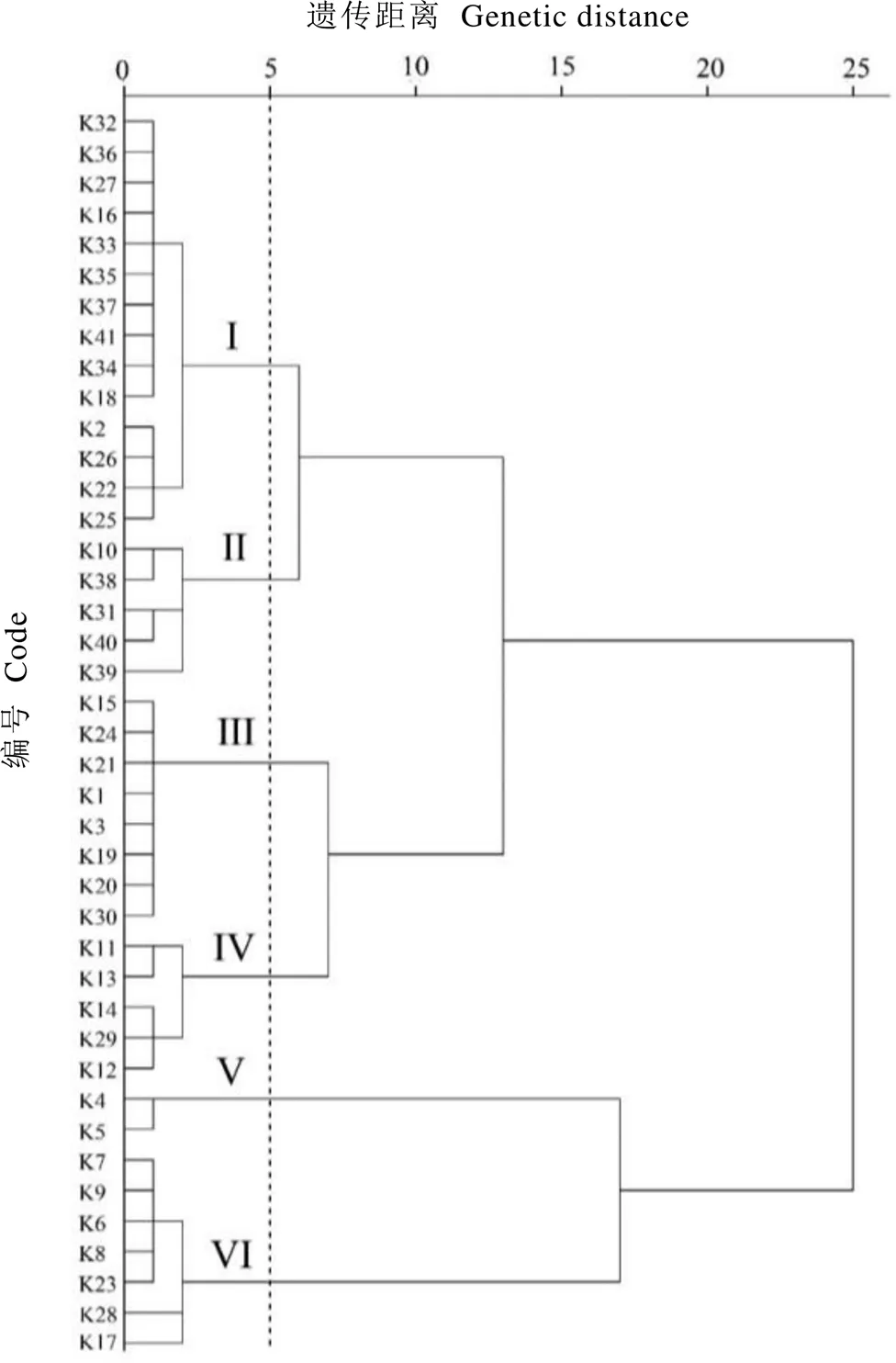
图1 41份茶树种质资源嘌呤生物碱含量聚类图
表3 41份种质资源各类群嘌呤生物碱含量特征表(平均值±标准误)
注:同行数据后不同小写字母表示在0.05水平上差异显著(类群间比较)
Note: Means followed by small letters indicate significance of difference at<0.05 level

表4 嘌呤生物碱含量特异的种质
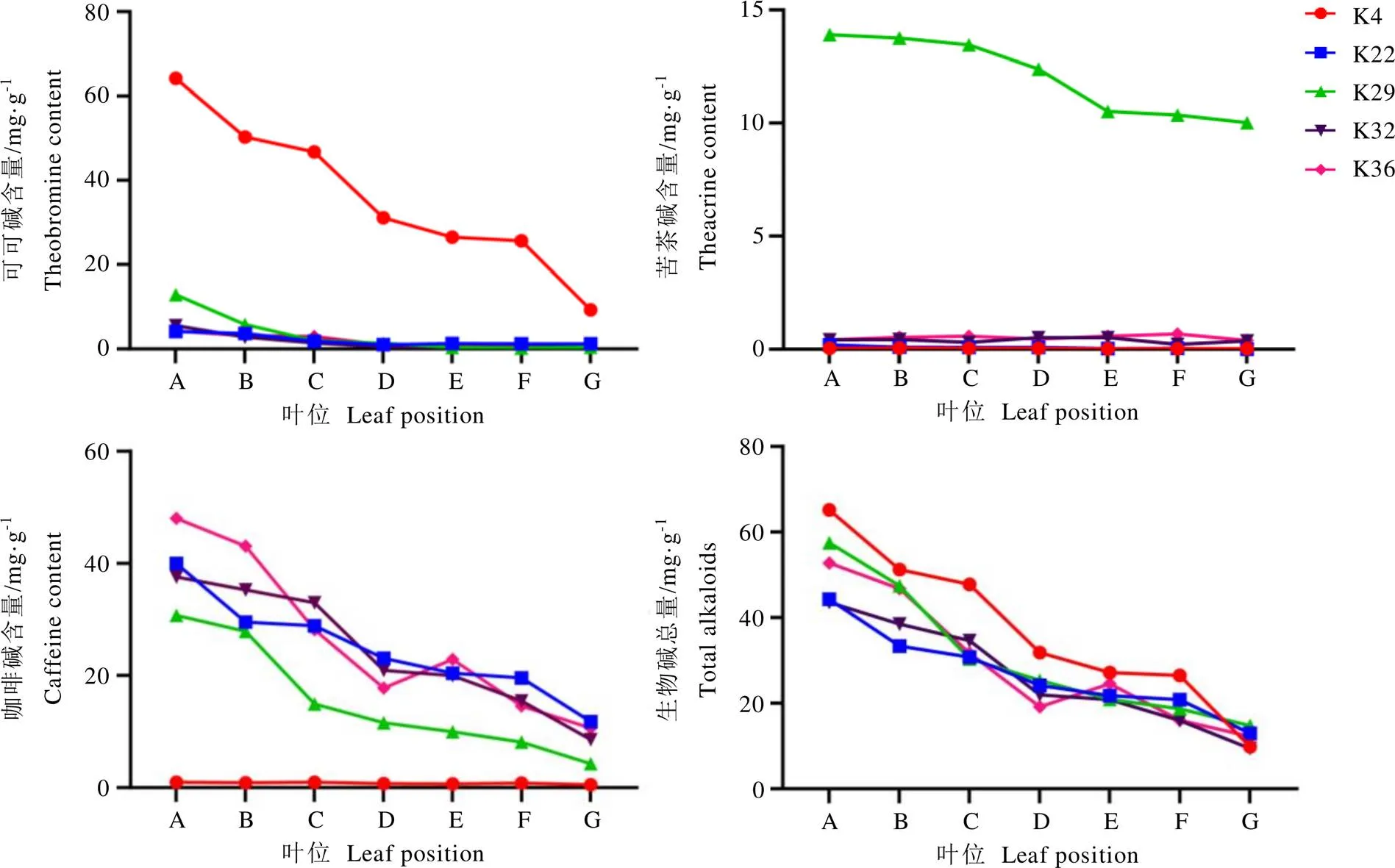
注:A:芽头;B:第一叶;C:第二叶;D:第三叶;E:第四叶;F:第五叶;G:成熟叶
3 讨论
我国是茶树起源地,种质资源十分丰富。苦茶作为我国特有的茶树资源,对其进行适当的评价和筛选将为今后开发利用提供理论参考。本研究对41份茶树资源嘌呤生物碱分析表明,不同种质间各嘌呤生物碱含量差异较大、变异类型丰富,蕴含着广泛的遗传基础,可为茶树育种和嘌呤生物碱的遗传改良提供丰富的基因资源。
基于表型的聚类分析,能够在一定程度上反映各种质的亲缘关系和基因变异[24]。本研究所用常规育成品种(系)全部聚类到第Ⅰ、Ⅱ、Ⅲ类群内,且咖啡碱为其优势嘌呤碱。咖啡碱是决定茶叶风味的物质之一,往往作为茶树品种选育的重要目标。从以上结果推测,以往茶树品种选育过程中把高咖啡碱含量作为一个重要指标,而忽略了低咖啡碱品种的选育。南昆山毛叶茶作为一种天然低咖啡碱种质,单独聚类在类群Ⅴ,可能与此类资源属于比较进化类型有关[25]。类群Ⅳ和类群Ⅵ全部为苦茶资源,且苦茶碱含量显著高于其他类群(表3)。值得注意的是,第Ⅳ类群苦茶中嘌呤生物碱分布模式为咖啡碱>苦茶碱>可可碱,与以往报道的乳源群体[26]、秃房茶[27]等富含苦茶碱种质的嘌呤生物碱分布模式均不相同,可作为一类独特的遗传资源。基于聚类分析结果和嘌呤生物碱含量特征,从苦茶资源中筛选出特异种质10份,其中高苦茶碱种质4份,高苦茶碱、高咖啡碱种质1份,高苦茶碱、低咖啡碱种质5份。
进一步研究5份资源嘌呤生物碱在不同叶位累积情况发现,可可碱、咖啡碱、苦茶碱以及生物碱总量在各种质中均为芽头含量最丰富,并随着叶片成熟度的升高而逐渐降低,在成熟叶中达到最低值(图2)。这与前人报道茶树中生物碱随叶序的增加,其含量递减的变化趋势类似[28],但有别于滕杰等[27]对秃房茶的研究。相比而言,苦茶碱在不同叶位降低趋势较为平缓,在苦茶成熟叶中其含量仍维持在较高水平,这可能与苦茶碱处于嘌呤生物碱代谢途径下游,有着特殊的累积特性有关[29-30]。
综上所述,通过对广东地区苦茶资源嘌呤生物碱分析评价,明确了其遗传特征和分布模式,筛选出了生物碱含量特异的种质资源,拓展了种质基础,对功能性茶树品种的开发利用、优良基因的挖掘具有重要意义。
[1] 王新超, 姚明哲, 马春雷, 等. 我国苦茶资源主要生化成分的鉴定评价[J]. 中国农学通报, 2008, 24(6): 65-69. Wang X C, Yao M Z, Ma C L, et al. Analysis and evaluation of biochemical components in bitter tea plant germplasms [J]. Chinese Agricultural Science Bulletin, 2008, 24(6): 65-69.
[2] 漠丽萍. 勐海县苦茶资源现状及开发利用探析[J]. 现代农业科技, 2017, 699(13): 27-28. Mo L P. Discussion on resource status and development and utilization ofvar.in Menghai County [J]. Modern Agricultural Sciences and Technology, 2017, 699(13): 27-28.
[3] 黎娜, 黄怀生, 钟兴刚, 等. 湖南地方特色茶树资源江华苦茶研究进展[J]. 茶叶通讯, 2018, 45(3): 6-10. Li N, Huang H S, Zhong X G, et al. Research advance on Hunan local tea tree resources[J]. Journal of Tea Communication, 2018, 45(3): 6-10.
[4] 沈程文, 黄意欢, 黄建安, 等. 湖南典型茶树地理种群遗传多样性[J]. 农业生物技术学报, 2007, 15(5): 855-860. Shen C W, Huang Y H, Huang J A, et al. RAPD analysis for genetic diversity of typical tea populations in Hunan Province [J]. Journal of Agricultural Biotechnology, 2007, 15(5): 855-860.
[5] 李丹, 罗军武. 江华苦茶种质资源的亲缘关系及在茶树进化中的地位研究进展[J]. 湖南农业科学, 2011(1): 18-20. Li D, Luo J W. Relationship ofcv. Jianghua germplasm resources and its status in the process of tea evolution [J]. Hunan Agricultural Science, 2011(1): 18-20.
[6] 张宏达. 茶叶植物资源的订正[J]. 中山大学学报(自然科学版), 1984, 23(1): 3-14. Zhang H D. A revision on the tea resource plants [J]. Acta Scientiarum Naturalium Universitatis Sunyatseni, 1984, 23(1): 3-14.
[7] 石祥刚, 郑新强, 宋晓虹. 关于苦茶的新组合[J]. 中山大学学报(自然科学版), 2008, 47(6): 129-130. Shi X G, Zheng X Q, Song X H, et al. A new combination of Kucha [J]. Acta Scientiarum Naturalium Universitatis Sunyatseni, 2008, 47(6): 129-130.
[8] van Dieren S, Uiterwaal C S P M, van der Schouw Y T, et al. Coffee and tea consumption and risk of type 2 diabetes [J]. Diabetologia, 2009, 52(12): 2561-2569.
[9] Heckman M A, Weil J, Mejia E G D. Caffeine (1,3,7-trimethylxanthine) in foods: a comprehensive review on consumption, functionality, safety, and regulatory matters [J]. Journal of Food Science, 2010, 75(3): 77-87.
[10] Li Y F, Chen M, Wang C, et al. Theacrine, a purine alkaloid derived fromvar., ameliorates impairments in learning and memory caused by restraint-induced central fatigue [J]. Journal of Functional Foods, 2015, 16: 472-483.
[11] Qiao H Y, Ye X S,Bai X Y, et al. Theacrine: a purine alkaloid fromvar.with a hypnotic property via the adenosine system [J]. Neuroscience Letters, 2017, 659: 48-53.
[12] 龚频, 石富兄, 陈福欣, 等. 云南苦茶提取物抗氧化应激保护镉致糖尿病肾病小鼠的作用[J]. 现代食品科技, 2019, 35(8): 6-12. Gong P, Shi F X, Chen F X, et al. Anti-oxidative stress of Yunnan bitter tea extract protects mice in high fat-cadmium induced diabetic nephropathy [J]. Modern Food Science and Technology, 2019, 35(8): 6-12.
[13] 宛晓春. 茶叶生物化学[M]. 3版. 北京: 中国农业出版社, 2013. Wan X C. Tea Biochemistry [M]. 3rd ed. Beijing: China Agriculture Press, 2013.
[14] 杨春, 罗军武, 陈涛林, 等. 高咖啡碱含量的特异茶树种质资源的筛选[J]. 湖南农业科学, 2013(17): 9-12. Yang C, Luo J W, Chen T L, et al. Screening specific germplasm resources of tea plant with high caffeine content [J]. Hunan Agricultural Sciences, 2013(17): 9-12.
[15] 唐琴, 孙威江, 陈志丹, 等. 尤溪苦茶资源苦涩味物质测定与分析[J]. 食品科学, 2019, 40(18): 242-247. Tang Q, Sun W J, Chen Z D, et al. Determination and analysis of bitter and astringent substances in Youxi bitter tea resources [J]. Food Science, 2019, 40(18): 242-247.
[16] 叶创兴, 林永成, 苏建业, 等. 苦茶var.Chang et Wang的嘌呤生物碱[J]. 中山大学学报(自然科学版), 1999, 38(5): 82-86. Ye C X, Lin Y C, Su J Y, et al. Purine alkaloids invar.Chang et Wang [J]. Acta Scientiarum Naturalium Universitatis Sunyatseni, 1999, 38(5): 82-86.
[17] 叶创兴, Ashihara Hiroshi, 郑新强, 等. 一种野生茶树的新嘌呤碱模式[J]. 中山大学学报(自然科学版), 2003, 42(1): 62-65. Ye C X, Ashihara H, Zheng X Q, et al. New discovery of pattern of purine alkaloids in wild tea trees [J]. Acta Scientiarum Naturalium Universitatis Sunyatseni, 2003, 42(1): 62-65.
[18] 杨超, 马存强, 周斌星, 等. 苦茶特征性成分研究[J]. 昆明学院报, 2014, 36(3): 75-78. Yang C, Ma C Q, Zhou B X, et al. Characteristic components research in bitter tea [J]. Journal of Kunming University, 2014, 36(3): 75-78.
[19] 黄华林, 李家贤, 何玉娟, 等. 广东茶树种质资源保存与利用现状[J]. 广东茶业, 2006(5): 14-17. Huang H L, Li J X, He Y J, et al. The current status of tea plant germplasm conservation and utilization in Guangdong Province [J]. Guangdong Tea Industry, 2006(5): 14-17.
[20] 吴华玲, 陈栋, 李家贤. 广东特异茶树种质资源选育研究进展[J]. 广东农业科学, 2012(20): 15-17, 24. Wu H L, Chen D, Li J X. The investigation and reflection on “late spring coldness” injury of Guangdong tea area [J]. Guangdong Agricultural Sciences, 2012(20): 15-17, 24.
[21] 陈亮, 杨亚军, 虞富莲, 等. 茶树种质资源描述规范和数据标准[M]. 北京: 中国农业出版社. 2005. Chen L, Yang Y J, Yu F L, et al. Descriptors and data standard for tea () [M]. Beijing: China Agriculture Press, 2005.
[22] 相吉山, 徐峰, 索良喜, 等. 东北地区谷子地方品种和育成品种表型比较分析[J]. 植物遗传资源学报, 2018, 19(4): 642-656. Xiang J S, Xu F, Suo L X, et al. Comparison on the phenotype traits between landrace and caltivars of foxtail millet [(L.)] in northeast China [J]. Journal of Plant Genetic resources, 2018, 19(4): 642-656.
[23] 马小卫, 马永利, 武红霞, 等. 基于因子分析和聚类分析的杧果种质矿质元素含量评价[J]. 园艺学报, 2018, 45(7): 1371-1381.Ma X W, Ma Y L, Wu H X, et al. Assessment of mineral elements contents at the mango germplasm level based on factor analysis and cluster analysis [J]. Acta Horticulturae Sinica, 2018, 45(7): 1371-1381.
[24] 陈常颂, 陈志辉, 钟秋生, 等. 闽育茶树品种生育期与生化成分比较及遗传多样性分析[J]. 茶叶科学,2014, 34(2): 172-179. Chen C S, Chen Z H, Zhong Q S, et al. Comparison of growth period and biochemical composition on Fujian tea cultivars and genetic diversity analysis [J]. Journal of Tea Science, 2014, 34(2): 172-179.
[25] 李斌, 陈国本, 陈娟, 等. 茶树天然无咖啡碱珍稀种质资源的研究[J]. 广东茶业, 2000(4): 7-11. Li B, Chen G B, Chen J, et al. Analysis of a natural caffeine-free tea plant [J]. Guangdong Tea Industry, 2000(4): 7-11.
[26] 金基强, 周晨阳, 马春雷, 等. 我国代表性茶树种质嘌呤生物碱的鉴定[J]. 植物遗传资源学报, 2014, 15(2): 279-285. Jin J Q, Zhou C Y, Ma C L, et al. Identification on purine alkaloids of representative tea germplasm in China [J]. Journal of Plant Genetic Resources, 2014, 15(2): 279-285.
[27] 滕杰, 曾贞, 黄亚辉. 秃房茶嘌呤生物碱组成特点及生化品质成分的研究[J]. 广西植物, 2018, 38(5): 26-34. Teng J, Zeng Z, Huang Y H. Composition characteristics of purine alkaloids and biochemical components of[J]. Guihaia, 2018, 38(5): 26-34.
[28] 王熙. 茶树叶片中主要特征物质的年变化规律研究[D]. 合肥: 安徽农业大学, 2013. Wang X. A study on annual variation characteristics of the major compounds in tea leaves [D]. Hefei: Anhui Agricultural University, 2013.
[29] Zheng X Q, Ye C X, Kato M, et al. Theacrine (1,3,7,9-tetramethyluric acid) synthesis in leaves of a Chinese Tea, kucha (var.) [J]. Phytochemistry, 2002, 60(2): 129-134.
[30] Ashihara H, Sano H, Crozier A. Caffeine and related purine alkaloids: biosynthesis, catabolism, function and genetic engineering [J]. Phytochemistry, 2008, 69(4): 841-856.
Analysis and Evaluation of Purine Alkaloid Contents in Bitter Tea Germplasm Resources from Guangdong
LI Hongjian, QIN Dandan, JIANG Xiaohui, FANG Kaixing, WANG Qiushuang, PAN Chendong, LI Bo, WANG Qing, WU Hualing*
Tea Research institute, Guangdong Academy of Agricultural Sciences/Guangdong Key Laboratory of Tea Plant Resources Innovation and Utilization, Guangzhou 510640, China
The purine alkaloids of 13 bitter tea germplasms from Guangdong Province along with 28 representative cultivars (lines) from different tea producing areas in China were selected for analysis in this paper. The results indicate that genetic diversity index () of purine alkaloids ranged from 0.78 to 1.94, while the coefficient of variation () ranged from 34.89% to 191.19%. The samples could be divided into 6 groups based on cluster analysis: groupⅠ, Ⅱ and Ⅲ were predominantly consisted of conventional cultivars with high caffeine content; groupⅤcontained caffeine-free cultivars from Nankun Mountain, while all samples in groupⅣ and Ⅵ were bitter tea with high theacrine content. A total of 10 purine alkaloid specific bitter tea resources were identified. Moreover, theacrine showed a gradually decrease trend with the increase of leaf positions but maintained a high level in matureleaves of bitter tea. Taken together, our research provided a basisfor genetic improvement of functional tea cultivars with specific purine alkaloids in the future.
bitter tea, purine alkaloids, germplasms, genetic diversity, cluster analysis
S571.1;S324
A
1000-369X(2021)01-071-09
2020-05-27
2020-06-24
广东省基础与应用基础研究基金(2020A1515110397)、国家现代农业产业技术体系建设专项(CARS-19)、广东省农业科学院院长基金(201830)、科技创新战略专项资金(高水平农科院建设)(R2017PY-QY009、R2018QD-99)
李红建,男,助理研究员,主要从事茶树资源与遗传育种研究,lihongjian.hi@163.com。通信作者:wuhualing@163.com
(责任编辑:黄晨)
猜你喜欢
安徽农业大学学报(2022年2期)2022-11-21
当代化工研究(2022年17期)2022-10-12
中国典型病例大全(2022年10期)2022-05-10
绿色科技(2019年14期)2019-11-19
江苏农业科学(2019年5期)2019-09-02
热带农业科学(2018年6期)2018-09-26
江苏农业科学(2017年17期)2017-11-15
家庭医药·快乐养生(2017年6期)2017-06-16
家庭医学·下半月(2016年5期)2016-05-14
家庭科学·新健康(2016年5期)2016-05-12
