
秦岭太白山不同海拔锐齿栎林土壤微生物群落的变化特征*
2021-02-12 06:46冯秀秀赵发珠郭垚鑫任成杰
林业科学 2021年12期
李 益 冯秀秀 赵发珠 郭垚鑫 王 俊 任成杰
(1.陕西省地表系统与环境承载力重点实验室 西安 710127; 2.西北大学城市与环境学院 西安 710127; 3.西北农林科技大学农学院 杨凌 712100; 4.西北大学生命科学学院 西安 710127)
土壤微生物是土壤有机质周转和养分循环的主要驱动力,直接影响着生物地球化学循环等重要生态过程(Kulhnkovetal., 2006; van der Heijdenetal., 2008)。以土壤微生物为主要内容研究陆地生物地球化学循环过程与全球气候变化的关系时,利用海拔梯度模拟气候变化影响情景已成为近年来广泛应用的研究方法(李超男等, 2017)。尤其在森林生态系统中,沿海拔梯度变化的微生物特征及其与生物和非生物因子的响应关系,可为探索环境梯度下的微生物多样性格局提供认识框架,从而为预测气候变化驱动土壤微生物群落变化提供理论依据。
目前,关于土壤微生物多样性随海拔变化的规律并无统一定论(李超男等, 2017; 李桂香等, 2018)。如长白山森林土壤细菌多样性并无明显的海拔分布格局,但苔原带土壤细菌多样性随海拔升高而显著降低(Shenetal., 2013; 2015)。伊朗北部的赫卡尼亚森林中外生菌根真菌的多样性随海拔升高而单调下降(Bahrametal., 2012)。也有研究表明土壤真菌多样性随海拔变化未表现出明显规律性(Shenetal., 2014)或呈现出“空心(hollow)”的分布模式(Wangetal., 2016)。造成以上相异结果的原因可能是海拔梯度变化会影响植被类型,而不同植被可能通过凋落物及根系分泌物对土壤微生物群落产生影响(赵盼盼等, 2019)。王淼等(2014)在罕山的研究表明,不同海拔的植被类型差异显著影响微生物群落的结构组成(Bachetal., 2010)。赵盼盼等(2019)对黄山松(Pinustaiwanensis)林3个海拔梯度的研究表明,同一植被类型不同海拔的土壤微生物碳变化规律与不同植被类型海拔梯度的土壤微生物碳变化规律有所不同。然而,通过控制植被类型,在海拔梯度上对土壤微生物群落变化规律及驱动因子的认识仍然有限,此方面的研究对深入理解和认识森林生态系统的C、N、P等养分循环过程的微生物作用机制具有重要意义。
太白山为秦岭山脉的最高峰,是中国气候的南北分界线(张善红等,2011),也是中国重要的生态屏障,其北坡属于典型的暖温带森林生态系统。秦岭山脉不仅对全球气候变化非常敏感,而且具有典型的植被带垂直分布特征(马新萍等, 2017; 秦进等, 2017),是山地森林生态系统土壤微生物垂直变化的天然研究平台。锐齿栎(Quercusalienavar.acuteserrata)林是秦岭落叶阔叶林区的重要组分和典型植被,在我国北亚热带-暖温带过渡区森林土壤的固碳、水源涵养等方面具有重要作用(牛晓栋等, 2018)。以往研究多集中于大尺度海拔上的植物-土壤-微生物的相互作用,涉及多个植被类型。而本文为控制植被类型变化,仅选择了作为太白山北坡典型植被的锐齿栎林的土壤,利用Illumina测序技术,分析4个海拔梯度下的土壤微生物多样性和群落组成的变化,以期阐明土壤微生物群落特征沿同一植被类型的不同海拔的垂直变化规律及主要驱动因素。
1 材料与方法
1.1 研究区概况
太白山(33°45′—34°10′N,107°19′—107°58′E)研究区内年降水量600~1 000 mm,主要集中在夏季。年均气温为5.9~7.5 ℃,北坡略低于南坡。研究区内海拔1 060~3 771.2 m,沿海拔差异产生了气候、土壤和生物种群的明显的垂直分布带。为控制植被类型变化,选择分布在太白山北坡1 300~1 800 m的锐齿栎林土壤进行微生物研究,该地段属典型的暖温带森林生态系统,主要为棕壤(雷梅等, 2001)。
1.2 样品采集与处理
2016年8月在太白山北坡海拔1 361.6、1 524.2、1 630.5和1 769.3 m处,分别选取地形、地貌、坡度和坡向基本一致的3个重复样地(20 m×20 m),各海拔森林样地的锐齿栎优势度分别为0.75、0.85、0.83和0.85。在每个样地内除去枯枝落叶层后,采用“S”型方法用直径5 cm的土钻收集表层(0~10 cm)土壤样品10个,混匀后作为1个样品,共收集12个混合土样。将每个土样过2 mm筛以去除植物根系和凋落物残体,然后分为4份,其中1份立即测定土壤含水量,1份置于-80 ℃冰箱中用于土壤DNA分析,1份储存在4 ℃冰箱中用于微生物生物量分析,剩余土样于室温下风干后测定土壤的基本理化性质。
1.3 土壤理化指标测定

1.4 土壤DNA提取、PCR扩增和高通量测序
用Omega DNA试剂盒,从每个土样(分成3份,每份0.5 g新鲜土壤,共计1.5 g)中提取土壤微生物DNA,然后用NanoDrop2000分光光度计(Thermo Scientific,Wilmington,DE,USA)评估DNA的浓度和质量。通过使用引物515F(5′-GTGCCAGCMGCCG CGG-3′)和907R(5′-CCGTC AATTCMTTTRAGTTT-3′)进行靶向V4区的细菌16S rRNA的PCR扩增(Ruiz-Gonzalezetal., 2015)。使用引物ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA-3′)和ITS2(5′-GCTGCGTTCTTCATCGATGC-3′)实现真菌ITS1区域的扩增(Waltersetal., 2016)。将每个样品扩增3次,然后将相关的扩增子混合以提供1种最终的PCR产物。同时,将16S rRNA和ITS rRNA的每个混合基因在2%琼脂糖凝胶上电泳以改善PCR产物的质量和浓度。最后,将来自每个样品的等量PCR产物置于单个管中,送至上海生物研究所于Illumina MiSeq平台测序分析。
为使用QIIME对测序结果进行多路分解、质量过滤和处理(Caporasoetal., 2012)。首先,使用USEARCH v5.2.32进行序列分析,通过聚类不相似性<3%的相似序列来过滤和消除来自数据的噪声。其次,使用微生物生态学软件(Microbial Ecology pipeline software)选择16S rRNA和ITS rRNA操作分类单元(OTU),将相似性大于97%的序列归为同一种OTU(Caporasoetal., 2012)。最后,将完整的数据集与国家生物技术信息中心(NCBI)的序列储存(SRA)数据库进行比对。细菌序列号为SRP223561,真菌序列号为SRP223559。
1.5 数据统计分析
利用Excel 2013和SPSS 22进行数据的预处理和统计分析。用Origin 8.0软件绘图。单因素方差分析(one-way ANOVA)用于检验不同海拔的土壤基本性质、微生物生物量和群落组成的差异显著性(P<0.05)。使用Mothur软件(v.1.30.1)通过香农(Shannon)指数估算微生物α多样性;非度量多维尺度(NMDS)分析微生物β多样性;回归分析反映土壤微生物生物量与SOC和TN之间的关系。此外,用R(999个排列)中“vegan package”的相似性(ANOSIM)分析来评价海拔对细菌和真菌β多样性的影响。采用Spearman相关分析确定土壤微生物多样性与土壤性质间的关系。利用CANOCO 5.0软件进行冗余分析(RDA)以确定土壤微生物群落组成与土壤性质之间的相关性。
2 结果与分析
2.1 土壤理化性质的海拔变化


表1 不同海拔梯度的锐齿栎林土壤基本理化性质①Tab.1 General characteristic of soils at different elevational gradients of Quercus aliena var. acuteserrata

图1 微生物生物量(碳和氮)与土壤碳和氮之间的关系Fig. 1 Relationships between the microbial biomass (carbon and nitrogen) and the carbon and nitrogen in soilMBC: 微生物生物量碳Microbial biomass carbon; MBN: 微生物生物量氮Microbial biomass nitrogen. MBC与SOC呈正相关但无显著相关性(P = 0.089 1); MBN与TN呈显著正相关(y = 0.047 0x-4.136 4, R2 = 0.998 3, P = 0.025 9)。MBC was positively but not significantly correlated with SOC(P=0.089 1); MBN was positively and significantly correlated with TN(y = 0.047 0x-4.136 4, R2 = 0.998 3, P = 0.025 9).
2.2 土壤微生物多样性的海拔变化
随海拔升高,细菌和真菌的α多样性(香农指数)均表现出先增后减的趋势(图2),且细菌α多样性变化极显著(P< 0.01),而真菌α多样性在不同海拔间无显著差异(P= 0.818)。土壤细菌群落结构在海拔相近样地可聚集为一类(图3),即归纳为3个类群: 海拔1 361.1 m的低海拔类群、海拔1 524.2 m的中海拔类群及海拔1 630.5和1 769.3 m的较高海拔类群。而土壤真菌β多样性在不同海拔间无明显聚集模式。通过ANOSIM分析发现,海拔对细菌β多样性的影响(ANOSIM,R=0.648,P= 0.001)大于对真菌β多样性的影响(ANOSIM,R=0.142,P=0.128)。

图2 土壤细菌和真菌α多样性(香农指数) 沿海拔的变化Fig. 2 Changes of bacterial and fungal α diversity(Shannon index) along the elevational gradients不同字母表示差异显著(P<0.05)。 下同。Different letters mean significant difference at 0.05 level. The same below.

图3 锐齿栎林土壤细菌(A)和真菌(B)β多样性沿海拔的变化Fig. 3 Changes of soil bacterial(A) and fungal(B) β diversity along the elevational gradients of Q. aliena var. acuteserrata
2.3 土壤微生物群落组成的海拔变化
由图4A可知,细菌群落组成,相对丰度>1%的优势菌门是酸杆菌(Acidobacteria)(41.8%)、变形菌(Proteobacteria)(24.9%)、放线菌(Actinobacteria)(14.1%)、绿弯菌(Chloroflexi)(5.4%)、芽单胞菌(Gemmatimonadetes)(4.3%)、硝化螺旋菌(Nitrospirae)(2.3%)、疣微菌(Verrucomicrobia)(2.0%)和浮霉菌(Planctomycetes)(1.8%)。其中,酸杆菌、放线菌和浮霉菌的丰度随海拔升高而下降,但仅放线菌下降显著(P< 0.05); 变形菌和疣微菌随海拔升高而显著增加(P< 0.05); 绿弯菌、芽单胞菌和硝化螺旋菌随海拔升高先增后减; 其他的细菌门(包括未知的分类群)占总细菌类别的3.4%,且整体随海拔升高无显著差异。此外,在细菌的纲类水平上,变形菌的分支α-变形菌纲(Alphaproteobacteria)占主导地位(14.47%),且随海拔升高而极显著增加(22.67%~28.69%,P<0.01),但另外3个变形菌的分支,即β-变形菌纲(Betaproteobacteria)、γ-变形菌纲(Gammaproteobacteria)和δ-变形菌纲(Deltaproteobacteria)随海拔的升高无明显变化规律(P>0.05)。在细菌的目类水平上,酸杆菌目(Acidobacteriales)、索利氏菌目(Solibacteriales)、红螺菌目(Rhodospirillales)和根瘤菌目(Rhizobiales)最丰富,但随海拔升高变化不显著(P>0.05)。
由图4B可知,真菌群落组成,相对丰度>1%的优势菌门是担子菌(Basidiomycota)(81.5%)和子囊菌(Ascomycota)(17.5%)。随海拔升高,担子菌的丰度总体呈下降趋势,而子囊菌总体呈增加趋势,但二者变化均不显著。分类学分析(表2)表明,伞菌纲(Agaricomycetes),即担子菌的分支,是最丰富的真菌纲,占真菌纲类水平的80%以上,但其随海拔升高变化不显著(P>0.05); 伞菌目(Agaricales)、红菇目(Russulales)和鸡油菌目(Cantharellales)占真菌目类水平的60%以上,且随海拔升高无显著差异(P>0.05)。总体而言,海拔对真菌类群的各分类水平(门、纲、目)的影响小于细菌。
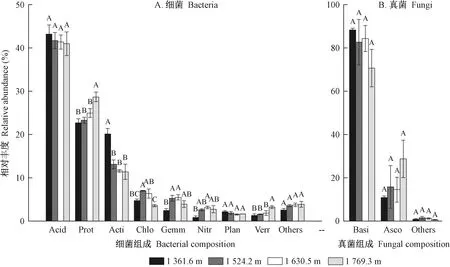
图4 不同海拔梯度锐齿栎林土壤细菌(A)和真菌(B)群落组成在门水平上的相对丰度Fig. 4 Relative abundance of the soil bacterial(A) and fungal(B) communities at the phylum level in different elevational gradients of Q. aliena var. acuteserrataAcid: 酸杆菌 Acidobacteria; Prot: 变形菌 Proteobacteria; Acti: 放线菌 Actinobacteria; Chlo: 绿弯菌 Chloroflexi; Gemm:芽单胞菌 Gemmatimonadetes; Nitr: 硝化螺旋菌 Nitrospirae; Plan: 浮霉菌 Planctomycetes; Verr: 疣微菌 Verrucomicrobia; Basi: 担子菌 Basidiomycota; Asco: 子囊菌 Ascomycota. 下同。 The same below.
2.4 土壤微生物多样性和群落组成与土壤理化性质的关系
Spearman相关性分析可知,细菌α多样性与SOC和C∶P显著正相关,而真菌α多样性仅与SM呈显著负相关; 细菌β多样性(Bray-Curtis Distance)与SOC、TN、TP、C∶P、N∶P和BD显著正相关,而真菌β多样性与所测土壤性质均无显著相关(表2)。
RDA分析表明,细菌门类水平中除酸杆菌、硝化螺旋菌和浮霉菌与土壤性质未呈现显著相关性以外,其余菌门均与土壤性质显著相关(图5A)。其中,BD与变形菌和疣微菌呈显著负相关,而与放线菌呈显著正相关; 芽单胞菌随海拔的变化与SOC和C∶P密切相关。在细菌的纲类和目类水平上,酸杆菌的分支嗜酸杆菌纲(Acidobacteriia)与SOC和C∶P呈显著负相关,酸杆菌目与SOC、TN和C∶P呈显著负相关; 变形菌的分支α-变形菌纲(在变形菌门中占比最大)与BD呈显著负相关,γ-变形菌纲与TN、MBN、N∶P和MBC∶MBN呈显著相关(图5C、E)。
对真菌门类水平,由图5B可知,土壤性质在门类水平上与真菌群落关系不大; 而在真菌的纲类水平中,除伞菌纲(Agaricomycetes)和锤舌菌纲(Leotiomycetes),其余真菌纲均与SOC、TN和N∶P表现出一定的显著相关性(图5D); 在真菌的目类水平中,土壤性质对伞菌目、红菇目(Russulales)、粪壳目(Sordariales)和肉座目(Hypocreales)的影响不显著,仅银合欢目(Agyriales)与SOC、TN和N∶P显著负相关(图5F)。总之,BD、ST、SOC、C∶P和TN极显著地影响了细菌群落组成,而pH、SOC、TN和N∶P对土壤真菌群落组成有较大影响。
3 讨论
3.1 锐齿栎林土壤微生物多样性沿海拔变化的差异性
本研究表明,在一定的海拔范围内,细菌和真菌的α多样性随海拔升高均呈先增后减的变化趋势,这与SOC、TN、TP、C∶P和N∶P沿海拔的变化趋势一致(图2),说明同一植被类型中沿海拔梯度的土壤养分含量对微生物群落起着重要作用,是决定微生物生存、物种组成和代谢的关键因素(赵盼盼等, 2019)。NMDS分析表明,细菌β多样性有明显的海拔分布格局,而真菌β多样性随海拔升高未呈现规律性变化(图3)。这与Meng等(2013)在中国亚热带山地森林中研究土壤微生物海拔变化的结果类似;但与Bahram等(2012)在伊朗北部研究的赫卡尼亚森林的海拔梯度变化结果有差异,这可能是本研究为同一植被类型且海拔梯度带来的温度差异较小,而Bahram等(2012)的研究选择的是不同植被类型且温度差异较大。然而,本研究中真菌β多样性对土壤理化特性的响应与Wang等(2015)对西藏森林的海拔变化研究结果有所不同,其原因可能是本研究为同一植被类型在有限海拔范围(1 361~1 769 m)的变化,而在西藏的研究跨越的海拔范围大(3 351~4 477 m)从而涉及不同植被类型,不同植被类型可通过植物根系的机械作用差异对土壤物理特性产生不同影响,也可借助根系分泌物和凋落物等的不同对土壤微生物活性产生不同的直接或间接影响,进而导致土壤微生物群落的变化(Lucasetal., 1993)。
3.2 锐齿栎林土壤微生物群落组成沿海拔的变化及驱动因子
本研究表明,细菌的优势菌门酸杆菌和浮霉菌在不同海拔间无显著差异。与本研究不同,贺婧等(2019)在秦岭太白山大尺度范围内对7个植被类型(海拔范围800~3 760 m)的研究表明,土壤细菌随海拔升高均有显著变化。这可能是本研究的海拔范围内的土壤水分海拔差异较小,不足以引起二者的丰度显著变化。另外,大多研究表明酸杆菌是寡营养型菌(Kielaketal., 2009; Naetheretal., 2012; Zhangetal., 2014),可能对环境变化的耐受性较强。对真菌群落组成,随海拔升高,担子菌总体呈下降趋势,而子囊菌总体呈增加趋势,但二者变化均不显著(图4)。Ren等(2018)通过研究秦岭太白山大尺度海拔范围内(1 300~2 600 m)的4个植被类型的土壤真菌,与本研究结果不一致,即随海拔增加出现先增后减再增加的变化,其原因可能是不同植被带中植物的凋落物与根系分泌物差异较大,且海拔梯度导致的温度、水分等多种因素之间的紧密联系存在复杂影响机制(Xuetal., 2017),对土壤真菌群落造成不同影响。
本研究发现,随海拔增加变形菌受土壤密度(BD)的显著影响(图5),γ-变形菌纲与全氮(TN)、微生物生物量氮(MBN)、N∶P和微生物生物量碳(MBC)∶MBN呈显著相关,且放线菌与土壤养分[如土壤有机碳(SOC)、TN、全磷(TP)、MBC、MBN、C∶P和N∶P]含量均呈负相关(图5C),说明在一定的海拔范围内,土壤理化性质对土壤微生物群落组成的影响比海拔或气候的影响更显著(Silesetal., 2016; Stroobantsetal., 2014)。此外,本研究发现真菌主要的优势菌门担子菌和子囊菌仅受土壤pH值的显著影响(图5B),这与前人研究结果类似,如Meng等(2013)认为子囊菌与pH值显著相关; Moreira Neto等(2009)认为,pH值是影响担子菌产生的木质素降解酶活性的重要因素。上述结果不仅说明一定海拔范围内的土壤特性(BD、ST、SOC、C∶P、pH和TN)变化是导致土壤细菌和真菌群落组成发生显著变化的主要原因,且进一步说明相比植物养分含量和土壤水热条件,土壤特性对微生物群落组成的影响更强烈。赵盼盼等(2019)在中亚热带戴云山的3个海拔(1 300、1 450、1 600 m) 的黄山松林研究表明,SOC、TN、TP、MBN、MBP和细菌、真菌生物量均随海拔升高而显著下降。因此,在减弱因海拔大范围变化所引起的多个因素变化的影响后,土壤特性对有限海拔范围内的微生物变化起到了主导作用。本研究中MBC与SOC无显著相关性,这与Michelsen等(1999)的研究结果不同,这可能是由于采样过程中及微生物保存过程中的不确定性造成的。
总之,土壤细菌和真菌多样性和群落组成的海拔差异会因海拔变化范围大小不同而存在差异,且具有不同的响应过程。本研究仅针对一定海拔范围内的森林土壤微生物群落的变化特征,还需进一步研究揭示这种差异的关键驱动因素及作用机制。
4 结论
1) 在秦岭太白山的一定海拔范围(1 361.6~1 769.3 m)内,锐齿栎林土壤细菌和真菌的α多样性随海拔升高呈先增后减的单峰变化。细菌α多样性的海拔变化主要与SOC和C∶P的海拔变化相关,而真菌的α多样性海拔变化只与SM的海拔变化显著相关。
2) 土壤细菌的β多样性具有一定的海拔变化规律性,且主要与SOC、TN、TP、C∶P、N∶P和BD的海拔变化有关,但真菌的β多样性沿海拔变化的规律不明显。
3) RDA分析表明,在限定的海拔范围内,土壤BD、ST、SOC、C∶P和TN的变化是影响土壤细菌群落组成显著变化的主要原因,而其对真菌群落的影响较小。
猜你喜欢
中国设备工程(2022年19期)2022-10-12
当代水产(2022年8期)2022-09-20
军事文摘(2022年14期)2022-08-26
九江学院学报(自然科学版)(2022年2期)2022-07-02
昆明医科大学学报(2022年2期)2022-03-29
散文诗(2021年22期)2022-01-12
食品安全导刊(2021年20期)2021-08-30
华东师范大学学报(自然科学版)(2019年3期)2019-06-24
环球时报(2019-05-23)2019-05-23
北京航空航天大学学报(2017年12期)2017-04-23
