
外源茉莉酸诱导对油菜防御酶活性和4种物质含量及植株生长的影响
2021-03-12 03:29冯建雄常静李海平杨凡李妍
植物保护 2021年1期
冯建雄 常静 李海平 杨凡 李妍
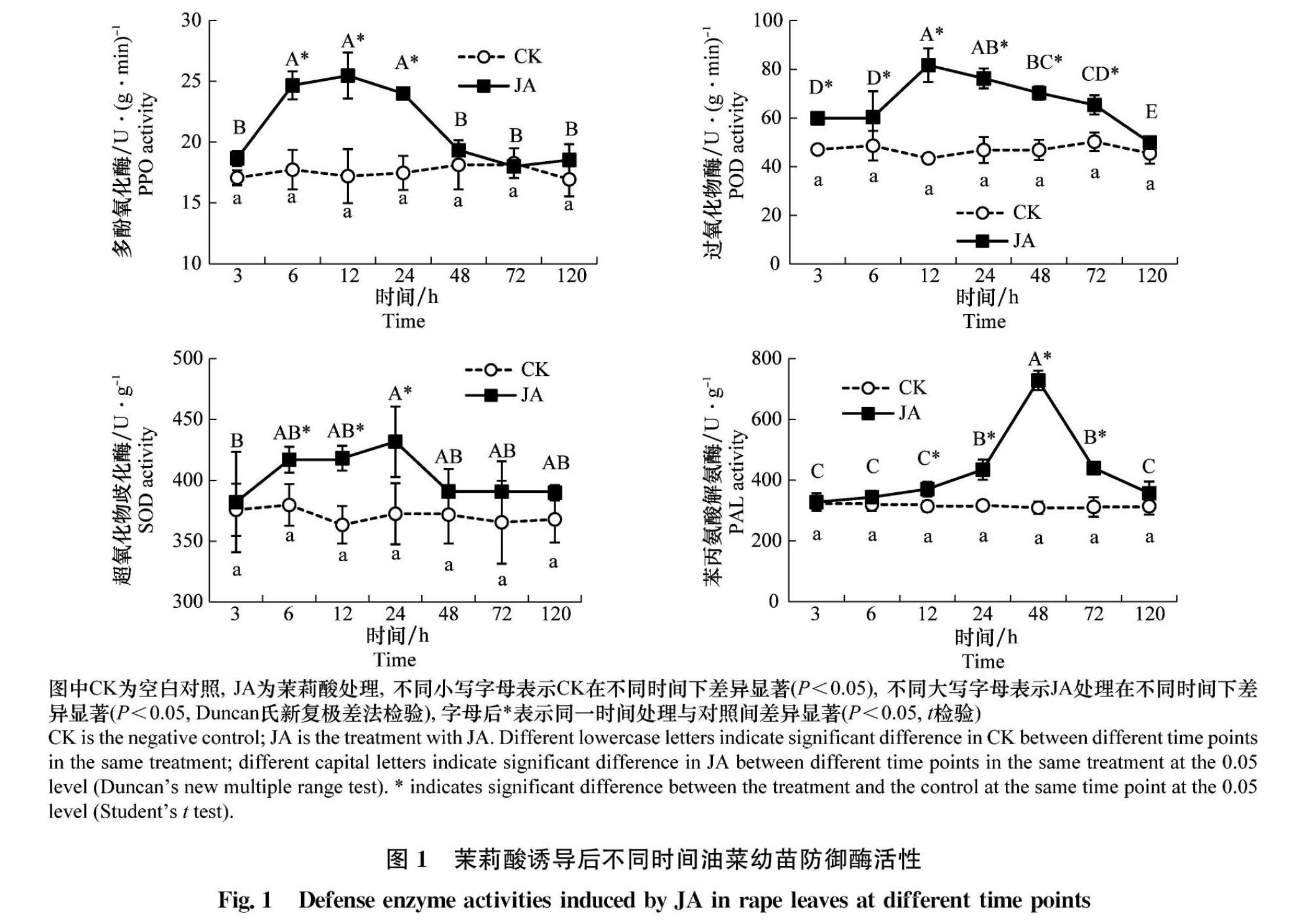
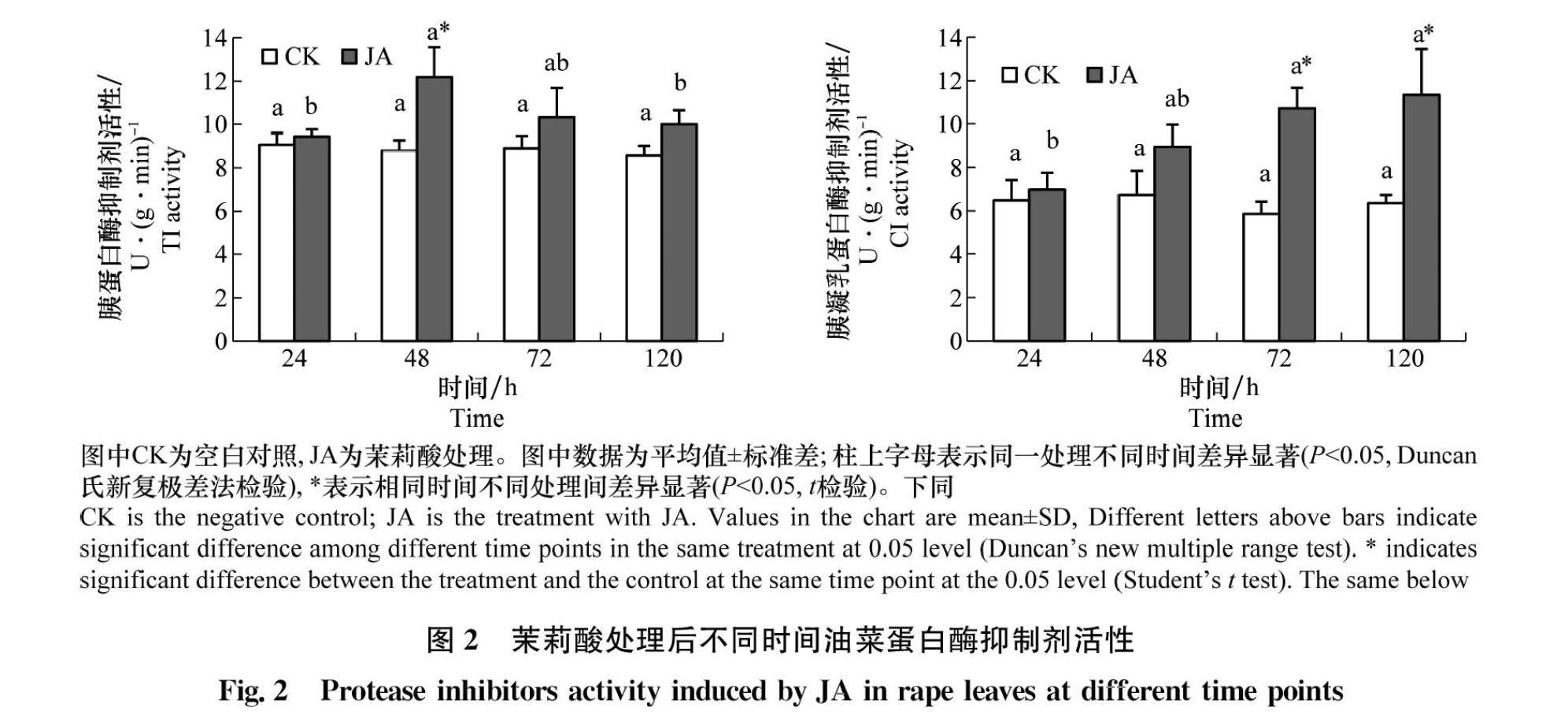

摘要 :油菜是我國重要的油料作物。近10年来油菜虫害呈重发态势,而茉莉酸可以诱导植物产生抗虫性。本研究测定了外源茉莉酸处理对‘大黄油菜幼苗中防御酶活性以及可溶性蛋白、可溶性糖、叶绿素、单宁含量及植株生长状况的影响。结果显示,以100 μmol/L茉莉酸处理后,油菜叶片中过氧化物酶(POD)、多酚氧化酶(PPO)、超氧化物歧化酶(SOD)和苯丙氨酸解氨酶(PAL)活性较对照均有提高。其中对PAL影响最大,48 h时上升为对照的2.36倍;POD和PPO活性均在12 h后达到最高,分别比对照增加了73.46%和49.21%;对SOD活性在处理后24 h比对照增加了14.93%;胰蛋白酶抑制剂(TI)活性在茉莉酸处理后48 h达到最大,比对照提高了38.37%,之后逐渐下降,至120 h时与对照无显著差异;胰凝乳蛋白酶抑制剂(CI)活性在茉莉酸诱导后一直增加,120 h时是对照的1.78倍。可溶性糖和叶绿素含量在处理120 h均降到最低, 分别比对照降低了38.70%和17.83%。可溶性蛋白含量则先降低后升高,在处理48 h后最低,比对照降低47.98%。可溶性单宁在茉莉酸诱导后含量持续升高,在120 h时上升为对照的2.03倍。油菜根长在茉莉酸处理后28 d和35 d显著长于对照;株高和干重均与对照无显著差异。上述结果说明适当剂量的外源茉莉酸诱导可激活油菜的防御体系,增强其抗虫性,且不影响油菜的生长。
关键词 :油菜; 茉莉酸; 防御酶; 营养物质; 可溶性单宁; 生长
中图分类号: S 433.1
文献标识码: A
DOI: 10.16688/j.zwbh.2019536
Effects of exogenous jasmonic acid on the activities of defense enzymes, the contents of four substances and the growth of rape
FENG Jianxiong, CHANG Jing, LI Haiping*, YANG Fan, LI Yan
(College of Horticulture and Plant Protection, Inner Mongolia Agricultural University, Hohhot 010019, China)
Abstract :Rape is one of the most important oil crops in China, and the insect pests of rape have been recurring in recent decades. Jasmonic acid (JA) plays an important role in plant resistance to insects. The changes in the activities of four defense enzymes and the contents of soluble protein, sugar, chlorophyll and tannin in ‘Dahuang rape subjected to exogenous JA treatment were measured in this study. The results showed that the activities of peroxidase (POD), polyphenol oxidase (PPO), superoxide dismutase (SOD) and phenylalanine ammonia lyase (PAL) increased significantly in rape seedling leaves treated with 100 μmol/L of JA compared with those of the control. The effect of JA on PAL activity was the strongest among four defense enzymes, 2.36 times of the control at 48 h. POD and PPO activities reached the peak 12 h after JA treatment, which increased by 73.46% and 49.21% compared with the control, respectively. SOD activity increased by 14.93% compared to the control at 24 h. The activity of trypsin inhibitors (TI) reached its maximum 48 h after JA treatment, which increased by 38.37%, and then decreased gradually. There was no significant difference from the control in 120 h. The activity of chymotrypsin inhibitors (CI) increased after JA treatment, reaching 1.78 times of the control at 120 h. The content of soluble sugar and chlorophyll decreased to the lowest at 120 h, 38.70% and 17.83% lower than those of the controls. The soluble protein content decreased first and then increased, reaching the lowest value 48 h after JA treatment, 47.98% lower than that of the control. The soluble tannin content increased after JA treatment, reaching 2.03 times of the control at 120 h. The root length was significantly different from control after JA treatment for 28 d and 35 d, and there were no significant difference in the stem length and dry weight. The results suggested that appropriate exogenous JA dose could induce defense response and enhance resistance of rape, and there was no significant effect on the growth of rape.
Key words :rape; jasmonic acid; defense enzymes; nutrient contents; soluble tannin; growth
茉莉酸(jasmonic acid,JA)是广泛存在于植物体内的天然化合物[1]。作为植物防御系统的重要信号分子,JA在植物抗逆性(如抵抗昆虫侵扰和防御机械损伤)方面起着重要作用[2]。用JA处理可以激活植物中的相应靶基因,从而增强有关防御酶活性[34]。Luo等利用病毒介导的基因沉默技术沉默了野生烟草2个JAIle羟基化酶基因,发现烟草中重要的抗虫次生代谢物质尼古丁、胰蛋白酶抑制剂以及二萜糖苷类化合物的含量上升,使其对斜纹夜蛾 Spodoptera litura 的抗性大大增强[5]。
油菜是世界上重要的油料作物,近年来随着人们对油菜研究的深入,油菜的用途不仅局限于提供人们的生活食用油,还扩展到饲料、工业原料、医药、绿肥等多个领域。油菜也是我国重要的油料作物之一,2017年全国油菜播种面积为665.30万hm2,产量为1 327.41万t[6]。近10年我国油菜种植面积和单产持续增长,同时油菜病虫害总体也呈重发态势,使油菜产业和油料安全受到重大影响[7]。
油菜上的虫害种类很多,严重为害的包括桃蚜Myzus persicae (Sulzer)、萝卜蚜Lipaphis erysimi (Kaltenbach)、甘蓝蚜Brevicoryne brassicae (Linnaeus)、黄宽条跳甲Phyllotreta humilis Weise、黄曲条跳甲P.striolata (Fabricius)、油菜虱跳甲Psylliodes punctifrons Baly以及小菜蛾Plutella xylostella (Linnaeus)等,其中黄宽条跳甲是油菜苗期的重要害虫,对油菜生产影响很大,化学防治是目前最常用的防治方法。当前,随着国家“两减”政策的实施,以及对绿色优质油菜籽需求的增长,寻找安全有效防治油菜害虫的方法成为当务之急,筛选抗虫品种及提高植物抗虫性可以从根本上解决这一问题。由于茉莉酸在植物抗虫性中的重要作用,本论文研究了外源茉莉酸诱导对油菜防御酶活性和3种营养物质及单宁含量的影响,以及茉莉酸处理后对油菜植株生长状况的影响,为探索使用外源茉莉酸诱导增加油菜抗虫性,降低化学农药的使用量提供理论依据。
1 材料与方法
1.1 供试油菜品种
‘大黄油菜(‘Dahuang rape)属于芥菜型油菜Brassica juncea L.,是呼和浩特市武川县的本地品种。实验室选用蛭石和珍珠岩按1∶1混合装入直径20 cm的花盆中,每盆播10粒种子,待油菜长至4~6葉时选健康且长势一致的幼苗供试。
1.2 试剂
98%茉莉酸(jasmonic acid,JA),氯化硝基四氮唑蓝(NBT),L苯丙氨酸和巯基乙醇(Sigma公司);聚乙烯吡咯烷酮(PVP)和EDTANa2(Amresco公司),其余药品均为国产分析纯。
1.3 油菜幼苗处理
根据前期预试验的结果,选择100 μmol/L茉莉酸进行试验。分为空白对照CK和喷施茉莉酸JA两个处理,各处理30盆4~6叶期的幼苗。丙酮溶解茉莉酸后用蒸馏水将茉莉酸稀释到100 μmol/L,对照选择与处理等量的丙酮、蒸馏水混合溶液,使用小型手持喷雾器将茉莉酸和丙酮蒸馏水混合溶液分别喷施到两个处理上,每盆喷施2 mL。在喷施后的3、6、12、24、48、72、120 h分别取样,去除叶脉称重后置于-80℃冰箱保存备用。
1.4 油菜防御酶活性测定
1.4.1 多酚氧化酶(PPO)活性测定
参考高俊凤的方法[8]。取0.5 g叶片加入液氮,充分研磨后加入5 mL pH 6.5的PBS溶液,在4℃ 5 000 r/min离心15 min,上清液即为PPO粗酶液。吸取0.5 mL粗酶液、1.5 mL PBS、1.5 mL 0.1 mol/L邻苯二酚,对照为煮沸失活的酶液。37℃水浴10 min,立即加入3 mL 20%三氯乙酸终止反应,于525 nm下测定吸光度,以吸光度每分钟变化0.01为一个酶活力单位,单位为U/(g·min)。
1.4.2 过氧化物酶(POD)活性测定
采用愈创木酚法[9]。取0.5 g叶片加入5 mL PBS(pH 7.0,0.1 mol/L),冰浴下研磨匀浆,匀浆液于4℃ 5 000 r/min离心15 min,上清液即为POD粗酶液。吸取0.1 mL酶液加入2.5 mL愈创木粉-H2O2混合溶液,于470 nm下测量吸光度。每隔30 s读一次,连续读3 min,以PBS替代粗酶液为空白对照。以吸光度每分钟变化0.10为一个活力单位,单位为U/(g·min)。
1.4.3 超氧化物歧化酶(SOD)活性测定
采用NBT法[8]。取0.5 g叶片加入5 mL PBS(pH 7.8,0.05 mol/L),冰浴研磨后于4℃ 10 000 r/min离心15 min,上清液即为粗酶液。在试管中加入甲硫氨酸(Met)(终浓度为13 mmol/L)、乙二胺四乙酸二钠(EDTANa2)(终浓度为10 μmol/L)、氯化硝基四氮唑蓝(NBT)(终浓度为75 μmol/L)、核黄素(终浓度为2 μmol/L)和酶液并定容至3 mL,另取两支对照管以缓冲液代替酶液。将上述对照管和一支测定管置于4 000 lx光照下反应20 min。用遮光的对照管为空白对照。以NBT为底物,以抑制NBT光化还原的50%为一个酶活性单位,单位为U/g。
1.4.4 苯丙氨酸解氨酶(PAL)活性测定
采用苯丙氨酸比色法[10]。取0.5 g叶片加入5 mL含5 mmol/L巯基乙醇的PBS(0.1 mol/L, pH 8.8)和少量聚乙烯吡咯烷酮(PVP),在冰浴下研磨成匀浆,于4℃ 5 000 r/min离心15 min,上清液为PAL粗提液。在试管中加入3.8 mL PBS,0.2 mL酶液,1 mL 0.6 mol/L L苯丙氨酸,对照以缓冲液代替L苯丙氨酸。反应体系在30℃水浴保温0.5 h,加入0.2 mL 6 mol/L盐酸终止反应。在290 nm下测定吸光度,以A290值每小时变化0.01为一个活力单位,单位为U/(g·h)。
1.4.5 胰蛋白酶抑制剂活性测定
参考张健等[11]的方法。取0.5 g叶片用TrisHCl缓冲液冰浴匀浆,于4℃ 2 000 r/min离心10 min,上清液即为抑制剂提取液。将0.4 g胰蛋白酶TrisHCl缓冲液溶解定容至50 mL,随配随用。取酶液100 μL和抑制剂提取液20 μL混合,于25℃水浴中反应1 h。取100 μL反应混合液加入2.9 mL含有100 μmol/L底物BAEE的TrisHCl缓冲液,于256 nm下测定吸光度值变化,单位为U/(g·min)。
TI活性=ΔA256/Ew×V×0.01
式中:ΔA256:256 nm处每分钟吸光度变化值;V:反应体系总体积;Ew:所用酶液含量。
1.4.6 胰凝乳蛋白酶抑制剂活性测定
按照张健等[11]的方法稍作修改。取0.5 g油菜叶片加入5 mL含有苯基硫脲(0.256 mg/mL)的TrisHCl缓冲液(pH 8.0),冰浴下研磨匀浆,4℃ 12 000 r/min离心10 min,上清液即为抑制剂提取液。将40%甲醇和TrisHCl缓冲液按照1∶1混合,用混合液溶解底物BTEE,配制成含有0.5 mmol/L BTEE的底物溶液,现配现用。称取0.185 g胰蛋白酶用TrisHCl缓冲液定容至100 mL,使用时稀释10倍。吸取酶液50 μL和提取液80 μL混匀并于25℃恒温水浴中保存20 min。取混合液100 μL加入2.9 mL底物溶液,于253 nm波长下测定吸光度值的变化,单位为U/(g·min)。
CI活性=ΔA253/Ew×V×0.964;
式中:ΔA253:253 nm处每分钟吸光度变化值;0.964:1 mmol/L BTEE的吸光度;V:反应酶液总体积;Ew:所用酶液含量。
1.5 油菜营养物质含量测定
1.5.1 可溶性糖含量测定
采用蒽酮比色法[10]。取0.5 g叶片加入液氮研磨后置于试管中,加5 mL蒸馏水于沸水中提取30 min。待提取液恢复至室温后5 000 r/min离心5 min,将上清液倒入25 mL容量瓶内,重复提取2次后定容。取提取液0.5 mL加入硬质试管中并加入1.5 mL蒸馏水,在冰浴中加入蒽酮乙酸乙酯试剂和5 mL浓硫酸,混匀后迅速于沸水中加热1 min,于630 nm下测定吸光度。用葡萄糖制作标准曲线,计算可溶性糖含量,单位为mg/g。
1.5.2 可溶性蛋白质含量测定
采用考马斯亮蓝G250法[12]。取0.5 g叶片加入液氮研磨后移至预冷离心管,5 000 r/min离心10 min,提取上清液备用。将1.0 mL提取液加入5 mL考马斯亮蓝G250中混匀,室温下放置2 min,于595 nm下测定吸光度。用牛血清蛋白制作标准曲线,计算可溶性蛋白质含量。
1.5.3 叶绿素含量测定
采用丙酮法[10]。取各处理叶片0.5 g放入具塞试管中,并在试管中加入丙酮和乙醇各4.5 mL和1 mL水,提取液总体积为10 mL。将试管放置黑暗条件下提取直到叶片完全变白,然后取上清液在663 nm和645 nm下测定吸光度。
叶绿素总含量=(8.02A663+20.21A645)×V/1 000×FW;
试中A663、A645:663 nm和645 nm的吸光度;V:提取液体积(mL);FW:样品鲜重(g)。
1.6 可溶性单宁含量测定
采用FolinCiocaileu法[13]。取油菜叶片0.5 g,装于25 mL具塞试管中,加水约25 mL,放在60℃左右的保温箱中过夜。第2天,过滤提取液至50 mL容量瓶中;加入5 mL 80℃左右的蒸馏水,置于80℃水浴锅中浸提20 min,反复浸提3~5次并定容至50 mL,即为单宁提取液。吸取1 mL单宁提取液加入7.5 mL蒸馏水,混匀后再加入0.5 mL FolinCiocaileu显色剂,摇匀并放置3 min,加入1 mL饱和碳酸钠。室温下放置1 h后在750 nm下测定吸光度。用单宁酸制作标准曲线,单位为mg/g。
1.7 油菜幼苗生长状况测定
油菜喷施JA溶液后,每隔7 d分别测定油菜的株高、根长及干重,每次测定10株,3次重复,共测定35 d。
1.8 数据处理与分析
采用Microsoft Excel 2010软件对数据进行汇总和处理,采用SPSS 12.0.1統计分析软件对数据进行方差分析,采用Duncan氏法对不同处理之间进行多重比较,处理与对照之间的差异显著性采用t测验;其显著水平设P<0.05。
2 结果与分析
2.1 茉莉酸诱导对油菜幼苗叶片防御酶活性的影响
茉莉酸诱导后油菜叶片中4种防御酶活性的变化见图1。4种防御酶活性基本都呈现先上升后下降的趋势。其中PAL活性变化最大,在48 h达到最大728.00 U/(g·h),显著高于对照(P<0.05),是对照的2.36倍。POD活性12 h达到最高峰81.69 U/(g·min),比对照增加了73.46%。 PPO活性在诱导12 h后达到了最高值27.45 U/(g·min),比对照增加了49.21%。SOD活性变化最小,24 h时比对照增加14.93%。
2.2 茉莉酸诱导对油菜幼苗蛋白酶抑制剂活性的影响
胰蛋白酶抑制剂(TI)的活性仅在处理后48 h显著高于对照(P<0.05),升高了38.37%,其他时间点与对照无显著差异。胰凝乳蛋白酶抑制剂(CI)活性在茉莉酸诱导72 h和120 h时均显著高于对照(P<0.05),分别是对照组的1.83倍和1.78倍(图2)。
2.3 茉莉酸处理对油菜幼苗叶片中营养物质含量的影响
由图3可以看出,外源茉莉酸处理可使油菜叶片可溶性蛋白、可溶性糖和叶绿素含量较对照显著降低(P<0.05)。可溶性糖含量在茉莉酸处理后呈现逐渐下降的趋势,120 h降到最低值3.12 mg/g, 比对照降低了38.70%。可溶性蛋白含量则呈现先降低后升高的趋势,在茉莉酸处理48 h达到最低值6.62 mg/g,比对照降低了47.98%。叶绿素含量呈降低趋势,在120 h达到最低值0.65 mg/g,比对照降低了17.83%。
2.4 茉莉酸处理对油菜幼苗叶片中可溶性单宁含量的影响
茉莉酸处理可使油菜叶片可溶性单宁含量显著增高(P<0.05),在120 h单宁含量达到了1.01 mg/g,是对照的2.03倍(图4)。
2.5 茉莉酸处理对油菜幼苗生长的影响
在茉莉酸处理后28 d和35 d,油菜根长显著高于对照(P<0.05),其他时间均无显著差异。茉莉酸处理对油菜株高和干重均无显著影响(表1)。
3 讨论
茉莉酸在自然界广泛存在,无论是天然还是外源茉莉酸,都对植物有诱导抗性、促进衰老等生理功能[1415]。很多研究已经证实茉莉酸在植物抗虫性中具有重要作用。
PPO能够催化多酚化合物次生代谢,降低植物的营养价值,阻止昆虫取食[1617]。桂连友等研究发现外源JA可以诱导茶树叶片内PPO活性增高,茶尺蠖Ectropis obliqua取食JA处理的叶片后生长受到明显抑制[18]。JA处理可诱导番茄叶片中的PPO活性增加,取食JA处理叶片的烟草天蛾Manduca sexta幼虫生长受到抑制[19]。王杰等的研究也有相似的结论[20]。本研究也发现,在外源茉莉酸诱导后,油菜体内多酚氧化酶(PPO)活性在诱导12 h后增加了49.21%。PPO参与酚类化合物的氧化,这种氧化作用与细胞壁伸展蛋白的偶联有关,因此PPO活性升高可增强油菜细胞壁的强度,从而提高植物的抗虫性。
外源茉莉酸处理油菜幼苗叶片后POD活性显著增加,该结果与王俊斌等在烟草叶片、段文昌等在枸杞叶片、田旭涛等在番茄和菜豆叶片上的结果一致[2123]。POD在植物防御中的作用是多方面的,其产生的醌类和其他氧化物可以与任何的游离氨基酸或蛋白质侧链反应以减少植物对昆虫的吸引。
PAL是连接初级代谢和苯丙烷类代谢并促进苯丙烷类代谢第一步反应的酶,是苯丙烷类代谢的关键酶和限速酶[24]。苯丙烷类代谢的中间产物经过一系列反应可转化为木质素、黄酮、生物碱等次生代谢物,一般这些次生代谢物被认为在物理和化学方面对植物生长发育和抗虫抗病性起着重要作用。SOD在植物清除活性氧方面起着重要作用,植物的抗逆性与植物体内能否维持较高SOD活性有关。段文昌等发现,外源茉莉酸诱导可显著提高枸杞叶片中PAL和SOD的活性[22]。杨世勇等用茉莉酸诱导棉花发现PAL活性增强[25]。同样用茉莉酸甲酯诱导巨桉幼苗也使巨桉叶中4种防御酶活性增强[26],与本研究的结果一致。
蛋白酶抑制剂在植物抗虫性中是重要的化学防御物质之一。很多研究发现,茉莉酸在不同植物中均可以诱导植物蛋白酶抑制剂活性上升。胡留成等研究发现,外用茉莉酸甲酯处理能系统性增强油菜胰蛋白酶抑制剂活性[27]。越慧芳等的研究也表明,不同浓度的外源茉莉酸处理青杨叶片后,胰凝乳蛋白酶抑制剂和胰蛋白酶抑制剂活性较对照均显著增加[28],本论文也有相似结论。蛋白酶抑制剂是植食性昆虫获取蛋白质难以克服的障碍,因此被作为茉莉酸处理后产生诱导抗性的标志[29]。
植物体内的营养物质是维持植物体正常运转必需的物质,也是植食性昆虫获取营养的主要来源。茉莉酸诱导会降低植物中主要营养物质的含量,使昆虫不能获得足够的营养,从而达到抗虫的目的。王予彤等在青杨上,李顺欣等在菜豆叶片上,宫玉艳等在枸杞叶片上以及孙兴华等在黄瓜叶片上的研究已经证实[3033]。植物防御是一个消耗能量的过程,植物在需要时才加以防御,是一种经济上可行的策略。本研究表明外源茉莉酸诱导会降低油菜体内可溶性蛋白和可溶性糖的含量,是该策略在实际操作中的具体体现形式[29]。
单宁是植物中常见的次生代谢物,能与昆虫唾液蛋白和消化酶结合,降低植物适口性,阻碍昆虫对寄主植物营养物质的消化和吸收[34]。外源茉莉酸可以使油菜叶片中可溶性单宁的含量明显上升,与严俊鑫等[35]在重瓣玫瑰上,宫玉艳等[32]在枸杞叶片上得到的结果一致。
除抗逆作用外,茉莉酸在植物生长上也有作用,如诱导植物开花结果,或是促进株高、根系的生长。植物体内的JAs可以增高POD的活性从而促进侧生根的生长[36]。本研究结果表明,茉莉酸处理28 d和35 d 后油菜根长显著高于对照,对株高、干重没有明显的促进作用。这可能是由于茉莉酸处理降低了油菜体内一些营养物质的含量,从而导致其对植物生长没有明显的促进作用。
目前的研究表明,外源茉莉酸處理不同植物后会增加植物体内抗虫性物质含量,提高植物的抗虫性。田旭涛等研究发现, 0.1、1 mmol/L 茉莉酸处理菜豆,显著降低了美洲斑潜蝇成虫对菜豆的取食和产卵选择性[37],茉莉酸诱导在葡萄抗根瘤蚜[38]、菜豆抗西花蓟马[31]、茶树抗茶尺蠖[18]等研究中也有相似的结果。但目前这些结果都是实验室内的研究结果,和茉莉酸的浓度有密切的关系,在大田当中使用是否有相同的结果,成本是否合适等,没有明确的结论。外源茉莉酸诱导后油菜体内的防御酶活性和一些物质含量发生了明显的变化,但油菜对黄宽条跳甲的抗虫性是否增加,在大田中是否有应用的可能性,还有待进一步更深入的研究。
油菜是我国重要的油料作物之一,我国油菜种植主要分为春油菜和冬油菜,其中春油菜主要分布在我国西北部地区,如内蒙古、青海、甘肃等地。内蒙古作为春油菜的主产地之一,冬季气候干燥、夏季冷凉湿润、日照强、昼夜温差大,适宜于春油菜的种植。且这样的气候条件也造就了春油菜良好的品质优势:含油量高,对人体有益的亚油酸含量高,因此油菜产业在内蒙古地区占有非常重要的地位。另外油菜在内蒙古地区还有观光、绿肥等特殊用途。因此降低油菜病虫害的发生,减少化学农药的使用,寻找安全有效的虫害防治方法,对满足绿色油菜籽的生产需求和促进油菜产业的发展都有重要的意义。
参考文献
[1] SEMBDNER G, PARTHIER B. The biochemistry and the physiological and molecular action of jasmonates [J]. Annual Review of Plant Physiology Plant Molecular Biology, 1993, 44: 569589.
[2] WASTERNACK C, HAUSE B. Jasmonates and octadecanoids: signals in plant stress responses and development [J]. Progress in Nucleic Acid Research and Molecular Biology, 2002, 72: 165221.
[3] KESSLER A, BALDWIN I T. Plant responses to insect herbivory: the emerging molecular analysis [J]. Annual Review of Plant Biology, 2002, 53: 299328.
[4] WEBER H. Fatty acidderived signals in plants [J]. Trends in Plant Science, 2002, 7(5): 217224.
[5] LUO Ji, WEI Kun, WANG Shuanghua, et al. COI1regulated hydroxylation of jasmonoyllisoleucine impairs nicotiana attenuatas resistance to the generalist herbivore Spodoptera litura [J]. Journal of Agricultural and Food Chemistry, 2016, 64(14): 28222831.
[6] 刘成,冯中朝,肖唐华,等. 我国油菜产业发展现状、潜力及对策[J]. 中国油料作物学报, 2019,41(4): 485489.
[7] 杨清坡, 刘万才, 黄冲. 近10年油菜主要病虫害发生危害情况的统计和分析[J]. 植物保护, 2018, 44(3): 2430.
[8] 高俊凤. 植物生理学实验指导[M]. 北京: 高等教育出版社, 2006.
[9] 孙群, 胡景江. 植物生理学研究技术[M]. 咸阳: 西北农林科技大学出版社, 2006.
[10]李合生. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2000.
[11]张健, 严善春, 王琪. 落叶松结实量对其防御酶和蛋白酶抑制剂活力的影响[J]. 林业科学, 2009, 45(8): 96100.
[12]BRADFORD M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of proteindye binding [J]. Analytical Biochemistry, 1976, 72: 248254.
[13]陈佩, 李平, 郝艳宾, 等. 柿果成熟过程中可溶性果胶和单宁含量的变化[J]. 中国农业大学学报, 2012, 17(1): 8892.
[14]CREELMAN R A. Jasmonic acid distribution and action in plants: Regulation during development and response to biotic and abiotic stress [J]. Proceeding of the National Academy of Science of the United States of America, 1995,92: 41144119.
[15]CREELMAN R A, MULLET J E. Biosynthesis and action of jasmonates in plants [J]. Annual Review of Plant Physiology and Plant Molecular Biology, 1997, 48: 355381.
[16]桂連友, 刘树生, 陈宗懋. 外源茉莉酸和茉莉酸甲酯诱导植物抗虫作用及其机理[J]. 昆虫学报, 2004, 47(4): 507514.
[17]THALER J S. Induced resistance in agricultural crops: effects of jasmonic acid on herbivory and yield in tomato plants [J]. Environmental Entomology, 1999, 28(1): 3037.
[18]桂连友, 陈宗懋, 刘树生. 外源茉莉酸甲酯处理茶树对茶尺蠖幼虫生长的影响[J]. 中国农业科学, 2005, 38(2): 302307.
[19]CIPOLLINI D F J R, REDMAN A M. Agedependent effects of jasmonic acid treatment and wind exposure on foliar oxidase activity and insect resistance in tomato [J]. Journal of Chemical Ecology, 1999, 25(2): 271281.
[20]王杰, 孟昭军, 王琪, 等. 外源茉莉酸诱导方式对长白落叶松针叶内防御蛋白活力的影响[J]. 东北林业大学学报, 2015, 43(5): 8489.
[21]王俊斌, 王海凤, 崔新仪, 等.茉莉酸甲酯对烟草抵御棉铃虫的诱导作用[J]. 中国农学通报, 2010, 26(17): 277280.
[22]段文昌, 段立清, 李海平, 等. 外源茉莉酸及枸杞瘿螨危害诱导的枸杞防御反应[J]. 昆虫学报, 2012, 55(7): 804809.
[23]田旭涛, 李丹, 张箭, 等. 外源茉莉酸对菜豆和番茄幼苗3种防御酶的诱导效应[J]. 西北农业学报, 2013, 22(9): 136140.
[24]AMRITA K, BRIAN E E. The phenylalanine ammonialyase gene family in raspberry, structure, expression, and evolution [J]. Plant Physiology, 2001, 127(1):230239.
[25]杨世勇, 王蒙蒙, 谢建春.茉莉酸对棉花单宁含量和抗虫相关酶活性的诱导效应[J]. 生态学报, 2013, 33(5): 16151625.
[26]张华峰.茉莉酸甲酯诱导巨桉对桉树枝瘿姬小蜂的抗性[J]. 森林与环境学报, 2016, 36(2): 216220.
[27]胡留成, 崔巍, 汪霞, 等. 斜纹夜蛾幼虫诱导的油菜抗虫性及其与茉莉酸信号途径的关系[J]. 昆虫学报, 2010, 53(9): 10011008.
[28]越慧芳, 段立清, 李海平,等. 外源茉莉酸诱导的青杨叶片保护酶活性变化及其对舞毒蛾幼虫生长发育的影响[J]. 昆虫学报, 2013, 56(3): 270275.
[29]KARBAN R, BALDWIN I T. Induced response to herbivory [M]. London: University of Chicago Press Ltd., 1997: 187191.
[30]王予彤, 越慧芳, 王晓丽, 等. 外源茉莉酸诱导的青杨生化抗性及其对舞毒蛾幼虫食物利用的影响[J]. 昆虫学报, 2015, 58(6): 673679.
[31]李顺欣, 郅军锐, 杨广明, 等. 外源茉莉酸诱导的菜豆叶片生化抗性及其对西花蓟马体内保护酶和解毒酶活性的影响[J]. 应用生态学报, 2017, 28(9): 29752983.
[32]宫玉艳, 段立清, 王爱清. 茉莉酸诱导对枸杞叶生化物质及酶活性的影响[J]. 植物保护, 2010, 36(2): 6165.
[33]孙兴华, 周晓榕, 庞保平, 等. 南美斑潛蝇为害对黄瓜体内主要营养物质、次生代谢物质及叶绿素含量的影响[J]. 昆虫学报, 2012, 55(10): 11781184.
[34]BARBEHENN R V, JAROS A, LEE G, et al. Hydrolyzable tannin as “quantitative defense”: limited impact against Lymantria dispar caterpillars on hybrid poplar [J]. Journal of Insect Physiology, 2009, 55(4): 297304.
[35]严俊鑫, 许凌欣, 宇佳, 等. 茉莉酸甲酯对重瓣玫瑰抗虫生理指标和双斑萤叶甲取食的影响[J]. 东北林业大学学报, 2017, 45(1): 7781.
[36]XUE Rengao, ZHANG Biao. Increased endogenous methyl jasmonate altered leaf and root development in transgenic soybean plants [J]. Journal of Genetics and Genomics, 2007, 34(4): 339346.
[37]田旭涛, 张箭, 李丹, 等. 茉莉酸处理菜豆对美洲斑潜蝇抗性的影响[J]. 植物保护学报, 2013, 40(8): 345349.
[38]杜远鹏, 季兴龙, 蒋恩顺, 等. 外源水杨酸和茉莉酸诱导巨峰葡萄抗根瘤蚜[J]. 昆虫学报, 2014, 57(4): 443448.
(责任编辑:田 喆)
猜你喜欢
茶道(2022年3期)2022-04-27
国外畜牧学·猪与禽(2022年1期)2022-04-23
中学生天地(A版)(2020年3期)2020-04-10
故事作文·高年级(2020年3期)2020-03-17
中学生理科应试(2017年7期)2017-08-09
红领巾·萌芽(2016年1期)2016-09-10
小小说月刊·下半月(2015年11期)2015-05-14
英语学习(2015年12期)2015-02-01
时代风采(2009年14期)2009-03-23
