
烟酰胺与丁酸钠对高密度笼养肉鸡肝脏和肠道氧化应激与炎症反应的影响
2021-03-15 05:40张雨晴武玉钦袁建敏
中国畜牧杂志 2021年3期
张雨晴,武玉钦,袁建敏
(中国农业大学动物科学技术学院,动物营养学国家重点实验室,北京 100193)
在现代畜禽生产中,养殖场常通过提高饲养密度来降低成本,导致家禽发生高密度氧化应激[1]。高密度氧化应激会导致家禽血清中皮质酮水平升高[2],生产性能、胴体品质下降[3],产生炎性介质致使细胞死亡[4]。核因子E2 相关因子2(Nrf2)信号通路是细胞氧化应激反应中的关键通路,可转录大量炎性因子[5]。研究发现,烟酰胺(NAM)可通过磷酸戊糖途径改善细胞氧化还原平衡[6],提高免疫细胞的增殖潜能[7]。丁酸钠可通过改善线粒体功能,提高机体抗氧化能力,维持小鼠肠道屏障完整性[8]。
目前,饲粮中添加烟酰胺和丁酸钠能否缓解高密度应激导致的肉鸡炎症、改善畜禽健康,还未见报道。本试验旨在研究烟酰胺和丁酸钠对高密度笼养肉鸡生产性能、血清及肝脏中抗氧化指标、盲肠扁桃体炎症因子的影响,为改善高密度饲养肉鸡的营养调控技术提供科学依据。
1 材料与方法
1.1 试验材料及动物 试验动物为AA+雄性肉鸡,1 日龄雏鸡购于北京某公司;烟酰胺(99%)购于江西兄弟医药有限公司;包被丁酸钠(有效成分30%)购于杭州康德权饲料有限公司。
1.2 试验设计与饲粮 1 日龄肉鸡在相同的环境和饲养管理饲养至26 日龄后,选择342 只体重相近约为1.2 kg 的肉鸡随机分为5 组,分别为正常密度组(12.9 只/m2)、高密度组(17.1 只/m2)、高密度+烟酰胺组(50 mg/kg)、高密度+丁酸钠组(500 mg/kg)和高密度+复合添加剂组(50 mg/kg 烟酰胺+500 mg/kg 丁酸钠)。每组6个重复,正常密度组每个重复9 只,高密度组每个重复12 只,饲养至46 日龄。基础饲粮参考NRC(1994)肉鸡营养需要配制玉米-豆粕型颗粒饲料,饲粮组成及营养成分见表1。
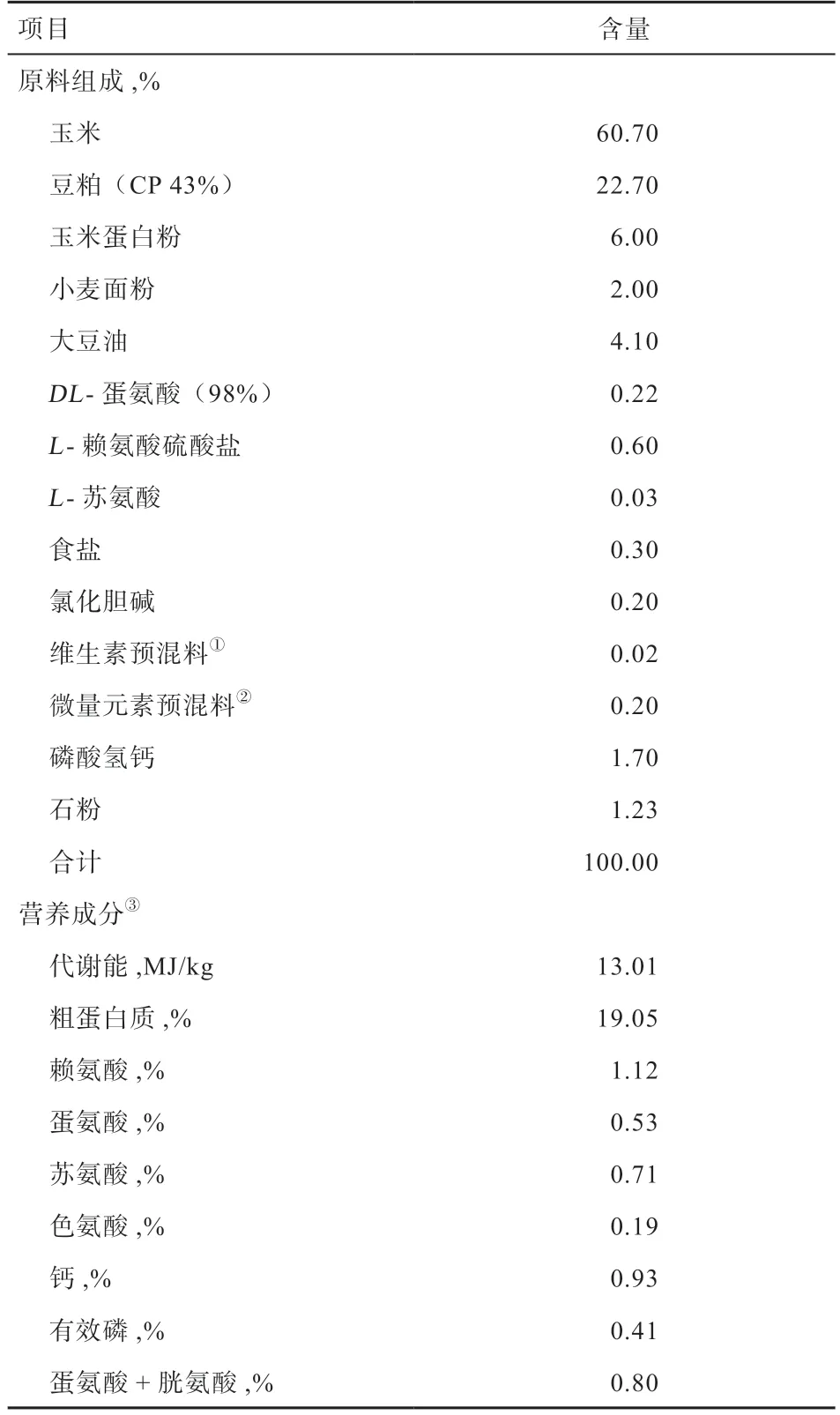
表1 基础饲粮组成及营养成分
1.3 饲养管理 试验在中国农业大学涿州养殖基地进行,试验期20 d。采用笼养(笼底面积0.7 m2),低密度9 只/笼(12.9 只/m2),高密度12 只/笼(17.1 只/m2),饲养管理参照AA+肉鸡饲养管理手册执行,自动化控制鸡舍温度、湿度和通风,并进行消毒、免疫。
1.4 样品采集和测定指标 46 日龄时,鸡只空腹12 h 后,以重复为单位进行称重,记录采食量,计算26~46 日龄体增重、采食量、耗料增重比。从每个重复选择1 只接近平均体重的鸡,翅静脉采血,2 500 r/min 离心10 min得到血清,置于-20℃储存待测。之后静脉注射戊巴比妥钠麻醉剂将肉鸡处死,迅速打开腹腔,采集肝脏、盲肠扁桃体分子样品,迅速放液氮中,而后置-80℃冰箱待测。
1.5 测定指标及方法
1.5.1 血清氧化还原酶活性 取待测血清,使用南京建成生物工程研究所试剂盒按照说明书测定谷胱甘肽过氧化物酶(GSH-Px,cat#A005)、超氧化物歧化酶(SOD,cat#A001-3)活性。
1.5.2 肝脏、盲肠扁桃体 mRNA 相对表达量 肝脏、盲肠扁桃体RNA 使用Trizol 试剂(TaKaRa,日本)进行提取,用核酸测定仪(Nano-drop2000)测定RNA的纯度和浓度。当OD260nm/OD280nm在1.8~2.0 说明RNA 质量较好,用于下一步试验。参照TaKaRa Prime ScriptTMRT reagent Kit with gDNA Eraser(Perfect Real Time)(TaKaRa,日本)反转录试剂盒说明将RNA反转录为cDNA。按照TB Green™ Premix Ex TaqTM(Tli RNaseH Plus)(RR420A,TaKaRa,日本)说明书进行荧光定量PCR,反应仪器为7500 荧光检测系统(Applied Biosystems),PCR 反应条件:95℃预变性30 s 后,进行95℃变性5 s,60℃复性34 s,40 个循环反应。引物DNA 由上海生工生物工程公司合成。PCR扩增完成后观察熔解曲线,并进行琼脂糖凝胶电泳鉴定扩增基因片段是否符合设计长度,验证扩增产物的特异性。肝脏抗氧化基因所用引物序列见表2,盲肠扁桃体免疫相关基因所用引物序列见表3,基因表达的结果采用2-ΔΔCT进行分析比较,以β-肌动蛋白(β-actin)作为参照基因校准。
1.6 统计分析 数据采用SPSS(v.20.0,SPSS Institute,Chicago,IL)的ANOVA 进行组间单因素方差分析,并进行Duncan's 多重比较,P<0.05 时表示差异显著。
2 结果
2.1 饲粮添加烟酰胺和丁酸钠对肉鸡生产性能的影响如表4 所示,饲养密度对肉鸡采食量、末重和体增重均有显著影响,高密度组采食量、末重和体增重均低于低密度对照组(P<0.05)。高密度单独添加烟酰胺或丁酸钠对采食量、体增重和耗料增重比均没有缓解效果,但高密度添加烟酰胺有提高末重的效果,高密度联合添加烟酰胺和丁酸钠可缓解高密度造成的末重和体增重下降,达到正常密度效果。
2.2 饲粮添加烟酰胺和丁酸钠对肉鸡血清抗氧化酶活的影响 由表5 可知,高密度组添加丁酸钠和烟酰胺对肉鸡血清中GSH-Px、SOD 活性没有显著性影响。

表2 肝脏抗氧化相关基因 Real-time PCR 引物序列

表3 盲肠扁桃体免疫相关基因 Real-time PCR 引物序列
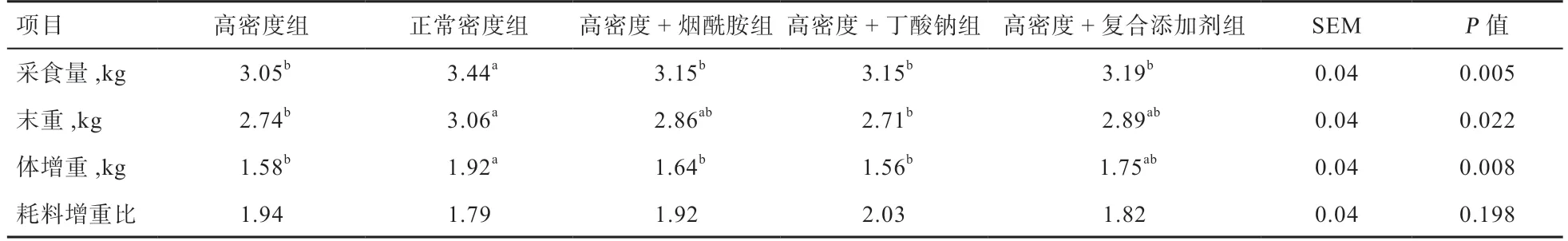
表4 饲粮添加烟酰胺和丁酸钠对肉鸡生产性能的影响

表5 饲粮添加烟酰胺和丁酸钠对肉鸡血清抗氧化酶活的影响
2.3 饲粮添加烟酰胺和丁酸钠对高密度饲养肉鸡肝脏抗氧化相关基因表达量的影响 如表6 所示,与正常密度组相比,高密度组的肝脏HO-1mRNA 表达量上调(P<0.05);高密度组单独添加丁酸钠、复合添加烟酰胺和丁酸钠后进一步上调了肝脏HO-1mRNA 表达(P<0.05);高密度降低了肝脏GSH-PxmRNA 的表达(P<0.05),高密度组添加烟酰胺和丁酸钠后可以提高GSH-PxmRNA 表达(P<0.05),达到正常密度的效果。
2.4 饲粮添加烟酰胺和丁酸钠对高密度饲养肉鸡盲肠扁桃体炎症相关基因表达量的影响 如表7 所示,高密度组的盲肠扁桃体促炎因子TNF-αmRNA 低于正常密度组(P<0.05),促炎因子IL6、抑炎因子IL10mRNA高于正常密度组(P<0.05);高密度组单独添加烟酰胺或丁酸钠均可提高TNF-αmRNA 表达(P<0.05),但高密度添加丁酸钠导致TNF-αmRNA 表达高于正常密度组,高密度添加烟酰胺+丁酸钠可以提高TNF-αmRNA 表达(P<0.05),达到正常密度的效果。
高密度组单独添加烟酰胺或丁酸钠以及复合添加组均可以降低IL6mRNA 表达(P<0.05),达到正常密度效果。高密度复合添加烟酰胺和丁酸钠组可降低盲肠IL10mRNA 表达(P<0.05),达到正常密度组一致的效果。
2.5 烟酰胺和丁酸钠对高密度饲养肉鸡 Keap1-Nrf2-ARE 抗氧化应激信号通路相关基因表达量的影响 由表8 可知,在Keap1-Nrf2-ARE 通路中,与正常密度组相比,高密度组盲肠扁桃体Nrf2mRNA 上调(P<0.05),高密度组单独添加丁酸钠或复合添加烟酰胺+丁酸钠进一步上调了Nrf2mRNA 表达(P<0.05)。
3 讨 论
3.1 烟酰胺和丁酸钠对高密度饲养肉鸡生产性能的影响 高饲养密度可降低肉鸡采食量、体增重和出栏重。Piva 等[9]研究发现,在高饲养密度下,高剂量烟酰胺(150 mg/ kg)可使24 日龄和40 日龄肉鸡体重分别提高6.8%和2.4%;Song 等[10]研究表明,丁酸是肠细胞首选的能量来源,日粮补充丁酸钠会降低肉鸡的平均日采食量、耗料增重比和死亡率。本试验中,高密度组的采食量、末重和体增重均显著低于低密度组,生产性能下降;单独添加烟酰胺和复合添加组均可改善高密度造成的肉鸡末重下降,复合添加剂组还可缓解高饲养密度引起体增重的降低,说明丁酸钠和烟酰胺具有协同效应,并且可以达到正常密度的效果。
3.2 烟酰胺和丁酸钠对肉鸡抗氧化功能的影响 肉鸡产业中,高饲养密度会对动物生产健康造成许多负面影响[11]。越来越多的证据表明,家禽生产中的大部分问题都与氧化应激有关[12],导致机体代谢紊乱,产生大量活性氧[13]。氧化应激还会产生大量应激蛋白(如CRP、热休克蛋白)并激活机体氧诱导的限速酶(如HO-1)加速组织氧化损伤[14]。CAT、SOD 和 GSH-Px 是机体最主要的酶促抗氧化系统,其主要作用是清除自由基和活性氧、防止过氧化物产生。Wu 等[15]试验表明,日粮添加丁酸钠可提高血清SOD 和CAT 活性,降低血清MDA 水平,还可提高肉鸡空肠黏膜的总抗氧化能力,降低空肠和回肠黏膜的MDA 含量。Choi 等[16]研究发现,添加烟酰胺可抑制ROS 生成,降低脂质过氧化和蛋白质氧化水平,提高线粒体还原水平。尽管本研究发现高密度添加烟酰胺+丁酸钠并没有改善血清中GSH-Px、SOD 含量,但肝脏的抗氧化基因GSH-PxmRNA 表达发生显著性差异,说明高密度添加烟酰胺和丁酸钠对肝脏的抗氧化能力产生了调节效应。本试验中添加丁酸钠和烟酰胺对高密度组肝脏中血红素加氧酶具有显著性调控作用,但对应激蛋白没有显著影响,有待于进一步研究。
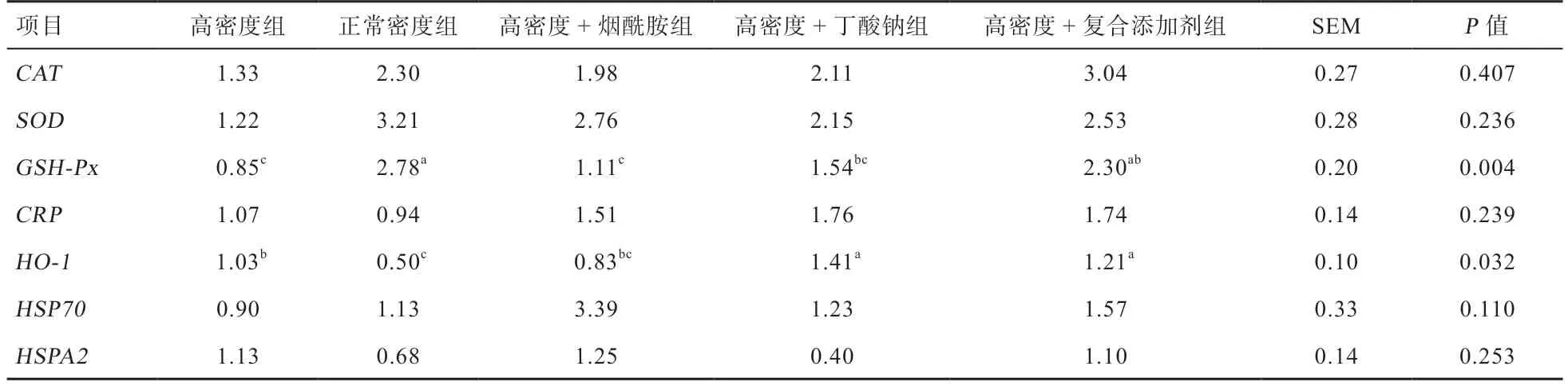
表6 饲粮添加烟酰胺和丁酸钠对高密度饲养肉鸡肝脏抗氧化相关基因表达量的影响

表7 饲粮添加烟酰胺和丁酸钠对高密度饲养肉鸡盲肠扁桃体炎症相关基因表达量的影响
3.3 烟酰胺和丁酸钠对机体炎症反应的影响 细胞因子虽在机体内含量较低,但作用高效,在机体免疫防御过程中发挥重要作用。Yarahmadi 等[17]研究表明,密度应激能够提高促炎细胞因子IL-1β和TNF-α的释放,促炎因子可以介导炎症发生,但是有研究发现适量的TNF-α增加能够增强机体的抗感染能力[18]。这解释了本试验中高密度组TNF-αmRNA 表达低于正常密度组,说明密度应激后抗感染能力下降,复合添加丁酸钠和烟酰胺可以提高TNF-αmRNA 表达,细胞因子的增加激活了机体第二道免疫防线。Lappas 等[19]研究表明,烟酰胺(NAM)可降低LPS 刺激的胎盘中促炎细胞因子TNF-α、IL-6以及炎性介质前列腺素PGE2等基因的表达。Yanez 等[20]研究其抗炎机制发现NAM 可通过抑制核因子κB,激活二磷酸腺苷核糖聚合酶(PARP1)生成,减少促炎因子IL8 的产生。丁酸钠不但可调节肉鸡盲肠扁桃体中免疫相关基因的表达,降低十二指肠黏膜中促炎因子水平以及NF-κB 的活化水平[21],还可降低鸡血清中抑炎因子IL10 的水平[22]。本试验中高密度单独添加烟酰胺或丁酸钠以及复合添加剂组均降低了促炎因子IL6mRNA 表达,只有复合添加剂组IL10mRNA 表达显著降低。因为促炎因子和抑炎因子具有协同作用,炎症早期,机体向促炎性免疫一端倾斜,加速免疫细胞进行病原体清除,说明营养调控对早期的炎症起了作用。IL10 是潜在抗炎因子,在炎症后期分泌,可以诱导T细胞免疫耐受,此结果说明烟酰胺和丁酸钠有助于恢复后期过度的炎症反应。
3.4 烟酰胺和丁酸钠对Keap1-Nrf2-ARE 通路的调控Kelch 样环氧氯丙烷相关蛋白-1(Keap1)-核因子E2相关因子2(Nrf2)-抗氧化反应元件(ARE)信号通路是细胞氧化应激反应中的关键通路,其调控下游相关代谢酶和抗氧化蛋白酶的合成[23]。抗氧化通路会影响炎症反应通路,活性氧在NF-κB 炎症信号通路中可以激活巨噬细胞表面上的Toll 样受体(TLR4),增加炎性因子IL6、IL8、TNF-α的基因表达,同时iNOS 会被激活[24]。另一方面,当 NF-κB 分子在信号通路被激活时,会引起一系列的级联反应和泛素化修饰,启动促炎因子的转录,进而导致炎症反应[25]。iNOS 是体内生成NO 的主要催化酶之一,在正常情况下并不表达,但受促炎因子和LPS 等刺激后会大量产生[26]。本试验发现,添加烟酰胺和丁酸钠后,Keap1-Nrf2-ARE 通路中Nrf2基因表达具有显著性变化。Nrf2 的化学组分是碱性亮氨酸,可以加速清除体内多余氧自由基。本试验中高密度组Nrf2基因表达明显升高,说明Nrf2 是细胞氧化应激反应中的关键因子,机体出现了应激反应。营养调控后进一步升高了Nrf2基因表达,说明活化的Nrf2 从Keap1 解离后进入细胞核,与Maf 蛋白结合为异二聚体后再同ARE 序列结合,启动下游抗氧化蛋白基因转录[27]。抗氧化通路中Keap-1 没有显著性变化,可能是Nrf2 的氨基酸有6 个保守的功能区,有的区域有丝氨酸残基,对Nrf2 的调节不依赖于Keap-1。另外,Nrf2本身含有氧化应激感受器,在氧化还原不平衡状态下较为敏感。
4 结 论
本试验结果显示,在高密度笼养条件下,家禽饲粮中添加烟酰胺和丁酸钠可以提高肉鸡的生产性能,缓解高密度氧化应激导致的肉鸡炎症基因表达。Keap1-Nrf2-ARE 信号通路可能参与了烟酰胺和丁酸钠对肉鸡抗氧化能力的调控。
猜你喜欢
动物营养学报(2022年9期)2022-10-12
动物营养学报(2022年8期)2022-09-01
动物营养学报(2022年4期)2022-05-12
当代水产(2022年3期)2022-04-26
现代农村科技(2022年1期)2022-01-21
资源信息与工程(2021年5期)2022-01-15
家庭医药(2021年13期)2021-12-03
安徽化工(2021年5期)2021-09-07
科学家(2021年24期)2021-04-25
科学家(2021年24期)2021-04-25
