
分子标记辅助选择改良黄华占和五山丝苗的褐飞虱抗性
2021-03-23 08:37毛方明王红波牟同敏
江西农业学报 2021年3期
毛方明,王红波,牟同敏
(1.华中农业大学 作物遗传改良国家重点实验室,湖北 武汉 430070;2.江西省农业科学院/江西省超级水稻研究发展中心,江西 南昌 330200;3.湖北省农业科学院 粮食作物研究所,湖北 武汉 430064)
褐飞虱是水稻上危害性最强的单食性害虫,具有极强的繁殖能力,主要危害亚洲栽培稻和野生稻。在2006~2015年的10年中,我国每年因稻飞虱造成的水稻产量损失可达到11.9亿kg,占水稻五大病虫害损失总量的29.51%[1]。褐飞虱的频繁暴发不仅对水稻的产量和品质造成了严重的影响,还因为杀虫剂的过量使用增加了水稻生产成本,破坏了自然生态系统平衡。多年实践证明,利用褐飞虱抗性基因培育抗褐飞虱水稻新品种,是最经济、环保且符合可持续发展战略的褐飞虱防治手段。
到目前为止已至少鉴定到34个主效的褐飞虱抗性基因位点[2-3]。除此之外,还有约100个褐飞虱抗性相关QTLs被报道。Bph3、Bph6、Bph9、Bph14、BPH18、Bph26、bph29和Bph32等8个抗性基因已经被克隆[3-6]。位于第3号染色体长臂端的Bph14基因是首个通过图位克隆技术获得的褐飞虱抗性主效基因[4]。Bph14基因可以使水稻叶鞘中产生大量胼胝质,保持筛管堵塞,同时还能促进胰蛋白酶抑制剂合成来达到抑制褐飞虱取食的目的[7]。Bph15位于第4号染色体着丝粒临近区域的重组冷点区约580 kb区间内,虽然该基因还未被克隆,但实践显示该基因具有较好的抗褐飞虱效果,已被广泛应用于水稻褐飞虱抗性育种中[8]。
利用MAS技术可快速进行优良基因渗入及多基因聚合。实践证明,利用MAS聚合褐飞虱抗性基因可以有效提高水稻的褐飞虱抗性。Wang等利用MAS将Bph6和Bph9基因聚合于93-11背景中,成功选育出农艺性状和稻米品质与93-11相似的双基因聚合系珞扬69,其褐飞虱抗性显著高于Bph6和Bph9的单基因导入系,并且由珞扬69组配的携带Bph6和Bph9杂合基因的杂交组合也表现出较高的褐飞虱抗性[9]。李进波等通过9311和1826水稻品种创建了Bph14和Bph15基因的单基因和双基因导入系,褐飞虱抗性鉴定显示其双基因导入系均达到高抗水平,Bph14、Bph15单基因导入系分别表现出中抗以上和抗或高抗水平,说明双基因的效果强于单基因[10]。
本研究分别以常规籼稻‘黄华占’和‘五山丝苗’为母本,与携带褐飞虱抗性基因Bph14和Bph15的供体材料‘HB13001-14-5’(简称HB1-14-5)进行杂交和连续回交,结合分子标记检测技术和田间表型选择,最终选育出2个褐飞虱抗性明显提高、其他性状综合表现与轮回亲本‘黄华占’相似的新株系‘HB6-8’和‘HB7-40’,以及2个褐飞虱抗性明显提高、其他性状综合表现与‘五山丝苗’相似的新株系‘HB1-44’和‘HB7-50’。
1 材料与方法
1.1 供试材料和分子标记
受体亲本‘黄华占’和‘五山丝苗’是广东省农业科学院水稻研究所选育的优质高产常规水稻新品种。本研究的‘黄华占’和‘五山丝苗’种子分别由湖北省种子集团有限公司和安徽荃银种业有限公司提供。供体亲本‘HB1-14-5’是本实验室从‘新6134/B5’后代选育的携带Bph14和Bph15基因的中间材料,抗褐飞虱。
本研究用于检测Bph14基因的紧密连锁的Indel标记为76-2,正反序列分别为5’-CTGCTGCTGCTCTCGTATTG-3’和5’-CAGGGAAGCTCCAAGAACAG-3’[4]。用于检测Bph15基因的Indel标记为InD4,正反序列分别为5’-AGAATGCTAAAGATGACTGAA-3’和5’-AACGGTATTGTTCTTGTCTAA-3’[8]。DNA的提取参照CTAB微量法;PCR扩增程序参考Du等的分子检测实验方法[4]。
1.2 苗期褐飞虱抗性的鉴定方法
本试验在华中农业大学褐飞虱鉴定网室进行。每个待鉴定的材料各3次重复,B5和TN1分别作为抗虫对照和感虫对照。具体操作方法和评价标准参考国际标准苗期集团筛选法[11]。
1.3 产量、主要性状和稻米品质的测定方法
于2018年夏季,在华中农业大学鄂州水稻育种基地进行产量比较试验,每个材料每个重复种植200株,行距是20 cm,株距是16.7 cm;每个材料3次重复,随机区组设计,四周种植保护行。在移栽后统一进行正常的田间管理。在生育期间记载生育期。在成熟时,每小区取样5株,进行株高、单株有效穗数、穗长、每穗总粒数、结实率和千粒重等主要性状的考查。分小区收获,在扬净、晒干(含水量13%)后,测定产量。稻米品质分析参照徐鹏等的方法[12]。
2 结果与分析
2.1 改良株系的创建过程
于2014年夏季,在武汉种植受体亲本‘黄华占’、‘五山丝苗’和供体亲本‘HB1-14-5’,并分别进行杂交。在同年冬季,在海南种植2个杂交组合的F1各20株;在苗期,利用与Bph14、Bph15基因紧密连锁的分子标记对全部单株进行分子检测;在抽穗期,选择两个目标基因均杂合的单株(即真杂种),分别与对应的轮回亲本回交,分别收获BC1F1种子;从BC1F1至BC3F1,每个世代都进行分子标记检测,筛选同时携带两个抗性基因的单株,并选择表型与轮回亲本最相似的单株与对应的轮回亲本进行回交。于2016年5月,在武汉分别种植具有‘黄华占’和‘五山丝苗’遗传背景的BC3F1世代,选择其中同时携带杂合Bph14、Bph15基因,表型与对应轮回亲本最相似的单株,收获BC3F2种子;在BC3F2世代,从880株的‘黄华占’/‘HB1-14-5’群体中筛选获得携带纯合Bph14和Bph15基因的单株39个,从720株‘五山丝苗’/‘HB1-14-5’群体中筛选获得携带纯合Bph14和Bph15基因的单株28个,分别收获自交种子;从BC3F3到BC3F5世代,每个世代继续利用分子标记对目标基因进行检测,并跟踪选择,同时对中选株系的苗期褐飞虱抗性进行鉴定;对BC3F5株系进行3次重复的产量比较试验,分析主要性状及稻米品质。在2018年秋季,决选出2个携带纯合Bph14和Bph15基因,主要性状及稻米品质与‘黄华占’相似的新株系,分别命名为HB6-8和HB7-40;同时决选出2个携带纯合Bph14和Bph15基因,主要性状及稻米品质与‘五山丝苗’相似的新株系,分别命名为HB1-44和HB7-50。改良株系的创建过程以及新株系抗性基因的分子检测结果分别见图1和图2。
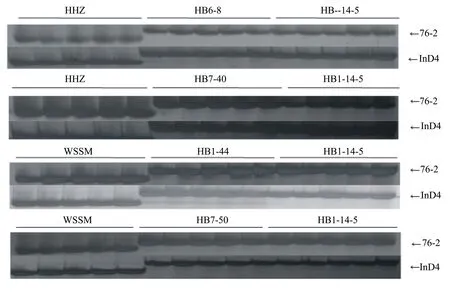
阴性对照为‘黄华占’和‘五山丝苗’;阳性对照为HB1-14-5;每个株系含5个泳道,76-2和InD4分别检测Bph14和Bph15基因。
2.2 改良株系的遗传背景回复率分析
将4个决选株系的种子进行催芽,从中随机选取30粒种子的幼芽进行混合取样。样品送至中国种子集团公司生命科学中心,使用水稻育种芯片RICE6K进行遗传背景分析。RICE6K芯片共包含5102个均匀覆盖水稻12条染色体的SNP和InDel标记,其中SNP的分布密度为每1 Mb 12个SNPs[13]。亲本间差异分析结果表明:在‘黄华占’与‘HB1-14-5’之间有678个SNP标记表现出多态性;而在‘五山丝苗’与‘HB1-14-5’之间有689个SNP标记表现出多态性。在改良株系中,来源于供体‘HB1-14-5’的染色体片段主要集中在抗性基因Bph14和Bph15的上下游区段(图3);除此之外,在其余染色体上也有少量供体来源的片段分布;株系HB6-8、HB7-40、HB1-44和HB7-50的遗传背景回复率分别为94.52%、90.91%、94.57%和92.69%。
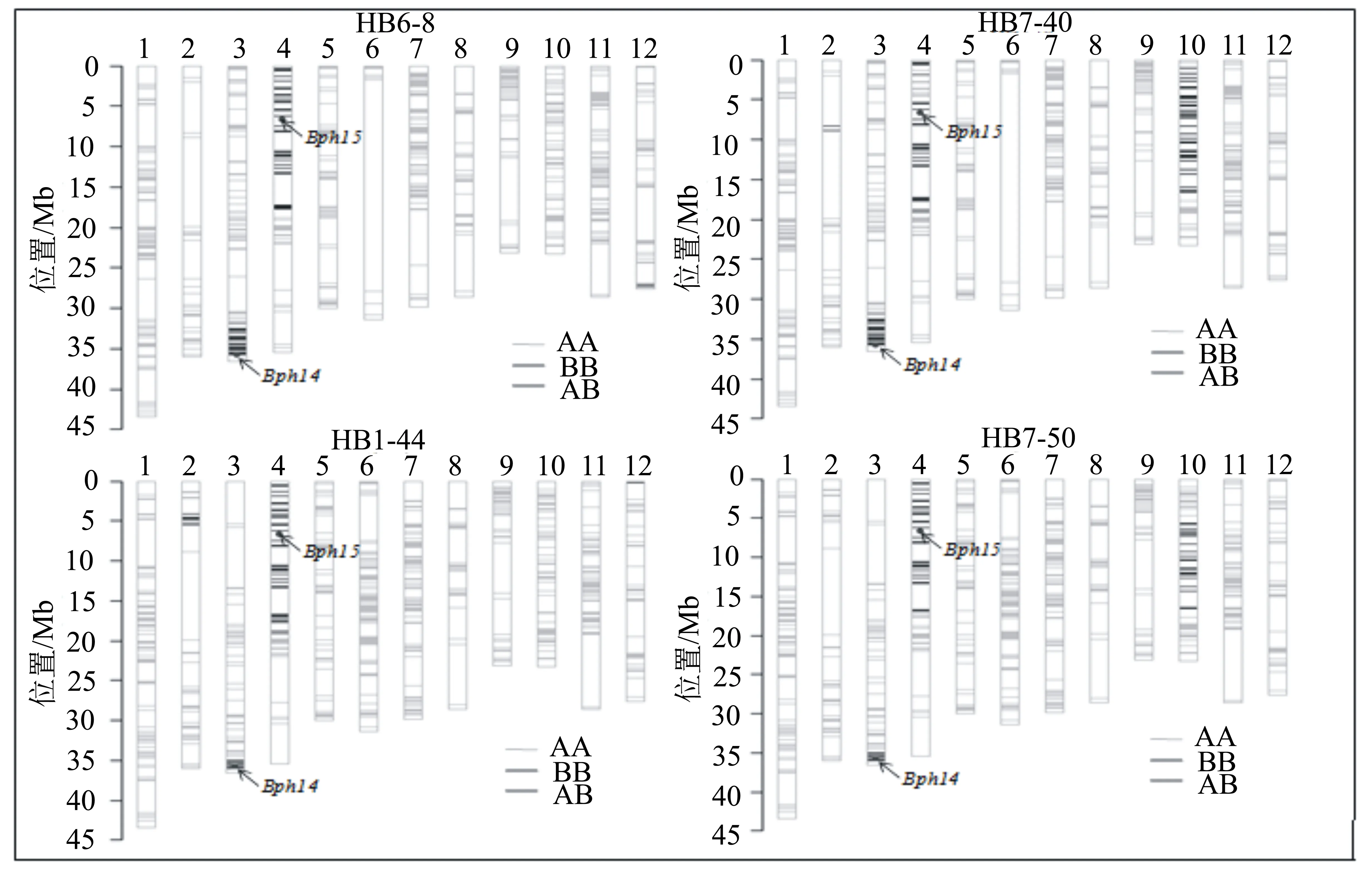
BB表示来源于供体亲本的染色体片段;AB表示杂合的染色体片段;AA表示来源于受体亲本的染色体片段。
2.3 改良株系在苗期的褐飞虱抗性表现
于2018年7月,在武汉对改良株系的苗期褐飞虱抗性进行了鉴定。以TN1作为感虫对照,以携带Bph14和Bph15基因的B5作为抗虫对照,每份材料3次重复。抗性鉴定结果表明:受体亲本‘黄华占’和‘五山丝苗’以及感虫对照TN1的平均受害等级分别为7.0、8.3和8.3级,表现为感或高感褐飞虱;供体亲本‘HB1-14-5’和抗虫对照B5的平均受害等级分别为3.0和1.7级,分别表现为抗和高抗褐飞虱;4个改良株系的平均受害等级为1.0~2.3级,表现为抗或高抗褐飞虱,其中具有‘黄华占’背景的两个改良株系HB6-8和HB7-40的平均受害等级均为2.3级,表现为抗褐飞虱;具有‘五山丝苗’背景的两个改良株系HB1-44和HB7-50的平均受害等级分别为1.7和1.0级,均表现为高抗褐飞虱(图4a和图4b)。
2.4 改良株系的产量、主要性状及稻米品质表现
改良株系的产量比较试验和主要性状考察结果(表1)表明:株系HB6-8和HB7-40的株高、单株有效穗数、穗长、每穗总粒数、产量与受体亲本‘黄华占’没有显著差异,但是播始历期比‘黄华占’缩短了2~3 d,在高温条件下结实率和千粒重比黄华占提高;HB1-44的播始历期、单株有效穗数、穗长、结实率和产量都与受体亲本‘五山丝苗’没有显著差异,但是株高和千粒重显著提高,每穗总粒数极显著降低;株系HB7-50除播始历期和每穗总粒数极显著低于‘五山丝苗’外,产量、其他性状与五山丝苗无显著性差异。
稻米品质分析结果(表2)表明,在分析的10项理化指标中,改良株系HB6-8和HB7-40与‘黄华占’基本相似,而HB1-44和HB7-50与‘五山丝苗’基本相似。
3 讨论
3.1 分子标记辅助选择技术有助于快速改良水稻的抗性
分子标记辅助选择(MAS)技术结合常规育种是目前进行作物育种改良的有效途径,其准确率和选择效率高,育种周期短。本研究采用该方法将Bph14 和Bph15基因渗入优良常规稻‘黄华占’和‘五山丝苗’背景中,分别获得了2个褐飞虱抗性显著提高,且产量、主要性状和稻米品质与受体亲本相似的新株系,可以作为黄华占和五山丝苗的抗褐飞虱替代品系进一步进行多点试验和区域试验。
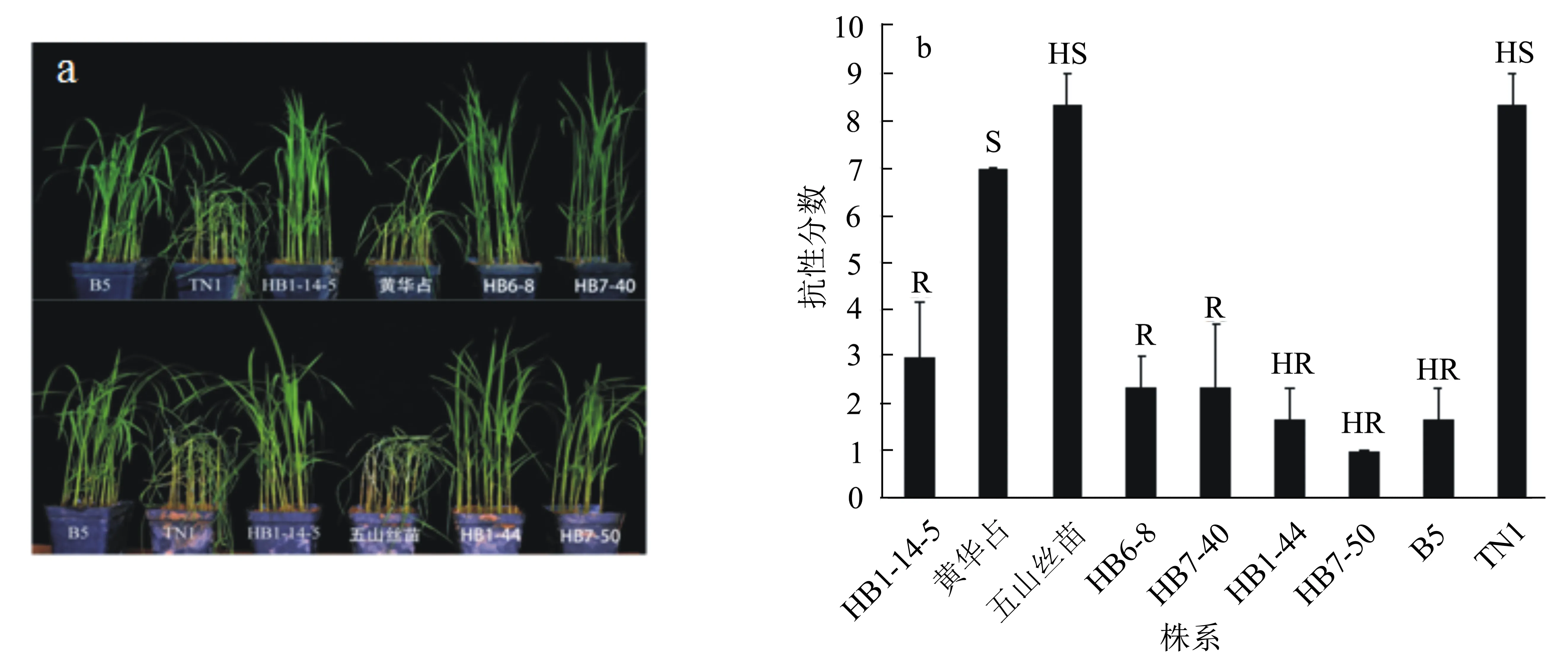
a:褐飞虱取食后幼苗的表现。b:幼苗的抗性分数及抗性水平。B5:抗虫对照。TN1:感虫对照。S、HS、R和HR分别表示感、高感、抗和高抗褐飞虱。
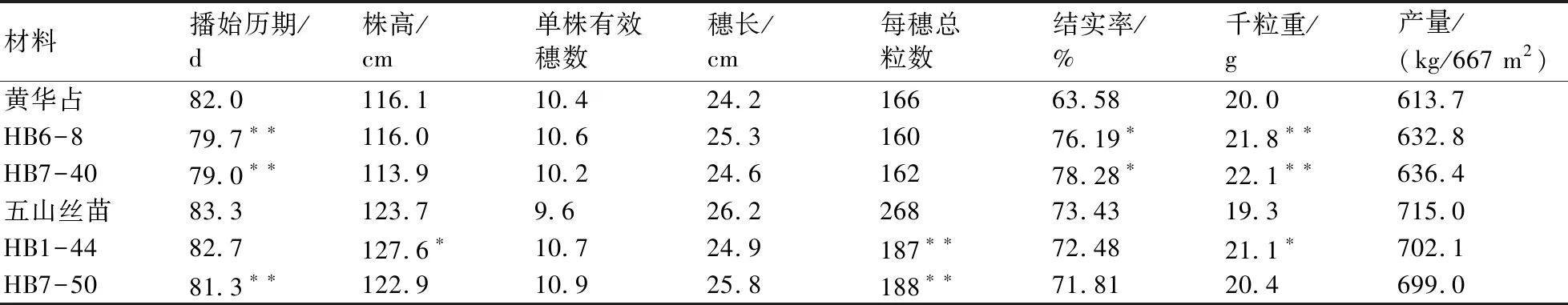
表1 改良株系2018年在鄂州的产量和主要性状表现
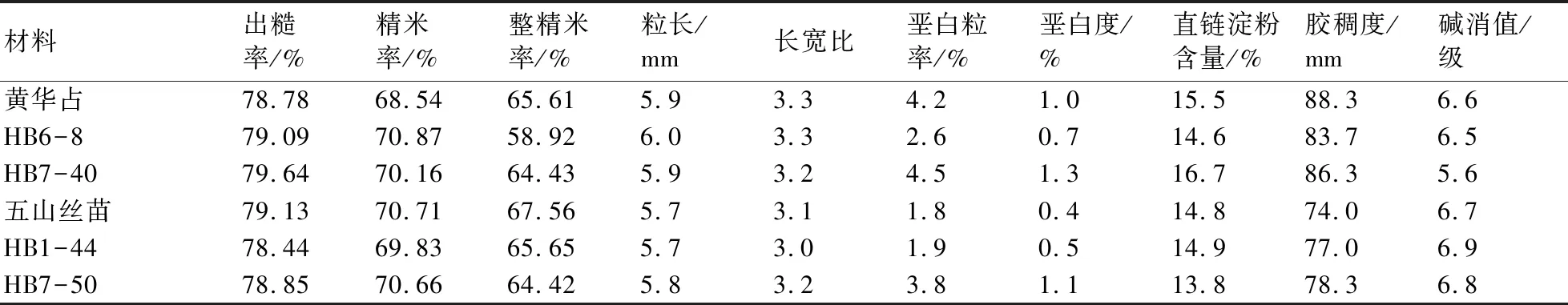
表2 改良株系2018年在鄂州的稻米品质分析结果
作物的抗虫、抗病等抗性基因主要受主效基因的控制,受外界环境因素的影响较小。通过分子标记辅助选择技术将这些抗性基因导入目标品种,以改良其抗性是一种十分有效的途径。例如,任西明等利用Bph14和Bph15基因,通过MAS成功改良了两系不育系C815S的褐飞虱抗性[14];徐鹏等通过MAS将Pi9、Xa23、Bph14 和Bph15基因聚合于水稻恢复系R1813中,获得了稻瘟病、白叶枯病和褐飞虱抗性均明显提高的新株系[12]。
利用MAS技术除了可以进行前景(目标基因)选择外,还可以对遗传背景进行选择。随着分子标记的普及和不断开发,SSR、RFLP、AFLP、SNP等分子标记技术被广泛应用于生物学科研的各个领域。前期利用分子标记进行遗传背景选择时,可以快速淘汰一些不符合育种要求的单株;对于背景回复率高的单株,则需要继续在未恢复为轮回亲本的位点附近继续增加标记进行回交选择,以此缩小与轮回亲本间的差异。MAS虽然能够给作物育种和改良提供便利,但在应用过程中也暴露出一些缺点,比如MAS只能从构成表型的所有变异中捕获主效基因带来的变异,而非主效基因累加的变异被忽视。为了达到更加精确的育种目标,可以利用全基因组选择方法,在基因组水平上对与目标性状相关的所有QTL进行检测,从而捕获主效和非主效基因带来的变异。
3.2 褐飞虱抗性基因Bph14和Bph15的利用价值及策略
Bph14和Bph15基因是目前褐飞虱抗性基因研究中使用较为广泛的两个主效基因,前人研究均表明其具有良好的褐飞虱抗性。本研究利用MAS将Bph14和Bph15基因分别聚合在常规稻‘黄华占’及‘五山丝苗’中,获得的4个新株系的褐飞虱抗性均达抗级水平以上,表明纯合Bph14和Bph15基因聚合能够达到抗褐飞虱的效果。Hu等的研究表明,Bph14和Bph15在‘明恢63’背景中表现出较高的褐飞虱抗性,在其所配的杂交组合中,Bph14和Bph15在单基因杂合、单基因纯合、双基因杂合、双基因纯合状态下均表现出一定的褐飞虱抗性,且抗性逐渐增强[16]。Wang等的研究结果表明,携带纯合Bph14和Bph15的亲本或杂交组合在苗期均具有优良的褐飞虱抗性,而携带杂合Bph14和Bph15基因的杂交组合在苗期均表现为感褐飞虱[17]。上述研究结果表明纯合Bph14和Bph15基因在不同遗传背景中均能表现出优良的抗性,而杂合Bph14和Bph15基因在不同遗传背景下其抗性可能有差异,表明Bph14和Bph15基因具有剂量效应,是不完全显性表达的。因此,若利用Bph14和Bph15基因进行抗虫杂交稻选育,最好选择携带相同褐飞虱抗性基因的不育系和恢复系进行配组,使杂交稻携带纯合的褐飞虱抗性基因,从而达到最佳的抗褐飞虱效果。
猜你喜欢
上海农业科技(2022年4期)2022-08-15
种子(2022年6期)2022-08-03
热带作物学报(2022年5期)2022-06-06
热带农业科学(2022年5期)2022-06-02
小学阅读指南·教研版(2022年3期)2022-04-28
包装工程(2022年3期)2022-02-22
上海师范大学学报·自然科学版(2020年5期)2020-12-18
江苏农业科学(2020年14期)2020-08-28
农业与技术(2019年6期)2019-05-29
南方农业学报(2018年9期)2018-09-10
