
不同微生境中水光温变化对毛尖紫萼藓叶绿素荧光特性的影响
2021-04-08 02:05王志侠陈慧妹殷美丽左世晨
生态学报 2021年4期
钱 申,王志侠,陈慧妹,殷美丽,左世晨,陶 冶,2,邱 东,2,*
1 安庆师范大学,生命科学学院/皖西南生物多样性研究与利用安徽省重点实验室, 安庆 246133 2 中国科学院新疆生态与地理研究所,荒漠与绿洲生态国家重点实验室, 乌鲁木齐 830011
作为生态系统中的先锋者和拓荒者,藓类植物在森林、高山、苔原及荒漠等生态系统中发挥重要生态功能,一些生长于极端环境中藓类植物经过长期进化适应,在形态、生理、遗传调控等方面形成了一定的避免不利条件伤害的策略[1]。如藓类植物呈垫状丛生以提高毛细管系统的持水力,叶细胞表面的密疣可以反射太阳辐射以减小蒸发,并有利于水分传导[2];叶先端的白色毛尖不仅可以增大生物结皮粗糙度、反射强光以减少植物体水分蒸发[3],同时还是一个高效的多尺度水分吸收与运输系统,极大提高了耐旱藓类的水分收集和固持能力[4- 5]。此外,在面对干燥失水情况时藓类叶片通常会皱缩,以减少水分散失及避免强光损伤[6]。尽管如此,藓类植物生长过程中也经常面临不确定的干旱、高温、缺水胁迫以及多种复合胁迫。藓类植物如何在形态、生理等多种功能性状间进行有效权衡,是其成功应对这些胁迫的关键。相比于微结构特征,藓类植物的生理特征往往具有更强的可塑性,在面临胁迫环境时可及时得到调整,以维持正常生命活动。
光合生理是植物最重要的生理过程,可以反映植物的生长状态,而叶绿素荧光特性能从内在揭示植物光合生理的表现[7- 8]。其中,叶绿素荧光参数最大光化学效率Fv/Fm反映的是PSII原初光能转化效率及PSII潜在活性,是反映光抑制程度的可靠指标;实际光化学效率Yield (ΦPSII)为PSII的实际光能转化效率,与PSII的活性呈正相关,其变化可以直接反映植物受逆境胁迫的状况[7]。研究表明,温度、光照、水分变化对藓类植物生理功能具有显著影响。脱水胁迫使藓类植物的渗透调节物质及抗氧化酶活性得以提高[9],可在一定时间内维持光合活性,而一旦植株含水量小于一定的阈值(如毛尖紫萼藓Grimmiapilifera含水量阈值为35%),其光合活性便迅速下降[10]。温度变化能引起藓类暗呼吸强烈变化,使CO2交换速率变化明显,并呈现复杂的协变关系[11]。高温胁迫下藓类植物叶绿素含量逐渐降低,光合系统遭到一定的破坏,活性降低[12]。低温胁迫下藓类植物PS II电子传递活性减弱,叶片暗反应受阻,光化学效率降低以及非光化学耗散增加[13]。光照是植物进行光合作用的前提,在一定的光强范围内随着光强的增大耐旱藓类的净光合速率、蒸腾速率和水分利用效率逐渐增加[14],其个体生物量及相对生长速率也显著增大[1];相比之下,弱光则使藓类的株高和叶绿素总含量增加,以利于捕获光能[15]。此外,随着植物遭受干旱、高低温、强辐射等逆境胁迫时间的延长,植物体Fv/Fm、Yield及电子传递速率(ETR)等均会呈现出一定的下降趋势[16]。尽管如此,水分、光照和温度3个因素组成的复合因子对藓类植物尤其是耐旱藓类光合生理的影响尚不明晰。
藓类植物的微生境复杂多样,其微环境的变化更多地表现在水分、温度和光照的变化上。遮荫和向阳是2种最常见的微生境类型,其中维管植物的遮挡是遮荫环境的重要形成方式[17]。灌丛显著影响维管植物生长发育,以灌丛中心向外延伸,土壤种子库密度、草本植物密度、盖度、生物量均呈现明显的递减趋势[18]。对藓类植物而言,灌丛遮荫的影响也十分明显。在干旱和半干旱地区,在沙丘背阴处藓类植物通常生长繁茂,且在地表的裂缝或降水过后的小冲沟中,也常常聚集着大量藓类[2]。在高温干旱期裸露地的齿肋赤藓(Syntrichiacaninervis)可溶性糖含量显著高于灌丛生境,体现出较高的抗旱性;而灌丛遮蔽环境下的齿肋赤藓植物盖度和生长状况要好于裸露地的藓类,但其抗性较弱[19]。因此,微生境显著影响了具有土壤基质的耐旱藓类生理特征[19]、个体生长发育和种群繁衍[20]。然而,一些石生的耐旱藓类生存环境更为残酷,其着生的岩石是绝对干燥的基质,岩石表面温度日变化幅度大且无法有效持水[21]。那么,这种石生的耐旱藓类在面对变化微环境时,具有何种生理适应特征?
毛尖紫萼藓广泛分布于我国南北各省,生长在裸露的岩石表面,是典型的耐旱藓类,已成为研究藓类植物耐旱机理的模式种之一[10]。毛尖紫萼藓微生境类型多样,因此其生长过程必然受到水光温变化的强烈影响[10]。但目前在不同微生境条件下毛尖紫萼藓的光合生理活性有何响应尚不清楚。为此,本研究以安徽省安庆市大龙山国家森林公园分布的毛尖紫萼藓为研究对像,在野外原位测定的基础上,以水分变化为主线,设置不同的降水量、降水频次(反映脱复水频次)以及光照和温度的组合,模拟荫蔽和向阳2种微生境,通过测定叶绿素荧光参数,分析毛尖紫萼藓对水光温条件复合变化的光合生理响应,并揭示各因素的影响作用大小。研究结果将为进一步探究石生耐旱藓类对不同微生境的适应策略提供一定的科学依据。
1 材料与方法
1.1 研究区概况
研究地点位于安徽省安庆市大龙山国家森林公园(116.994°—117.087° E,30.624°—30.629° N),属北亚热带湿润季风气候区,面积40.18 km2,最高海拔693.4 m,年平均气温14.5—16.6℃,降水量1300—1500 mm,年日照时数2012 h,四季分明,雨量充沛,光照充足,年无霜期247 d[22]。大龙山地处沿江湖泊丘陵区,水资源充足,植物资源丰富,有800余种维管植物;土壤以红壤、黄壤、黄棕壤类型为主[23]。该地区花岗石垂直节理发育和球状风化,具有典型的球状花岗岩地貌兼湖泊景观特色;毛尖紫萼藓便生长于表面凸凹不平的花岗石上。
1.2 野外原位叶绿素荧光参数测定
在多次踏查基础上,选择向阳和竹林遮蔽2种裸岩生境,选择长势相同、密度均匀的藓丛,放置直径4 cm不锈钢环标定待测藓类斑块,并以中心点作为待测位点。在连续3个晴天后,在30 min内用喷壶对整个藓丛(含待测斑块)进行多次喷水,保证藓类植物充分吸水。用遮荫布遮盖藓类斑块进行30 min暗适应[10],随即测定藓类植物的叶绿素荧光参数,间隔为每1 h一次,共5次;24 h后再测定一次干燥状态下的叶绿素荧光参数。每个生境5个重复斑块,每个斑块重复测定,且所有时间点测定的位置不变。
1.3 室内水光温控制实验
毛尖紫萼藓采自向阳和竹林遮蔽环境下的裸岩表面,每个生境5个重复斑块,取样时尽可能保证相同生境中采集的藓类斑块及藓类个体大小一致。在实验室内将2种生境藓类植物样品复水1 h,随后选择大小相近的20株藓类植物均匀插入不锈钢纱网中,并将其放在底部有滤纸、四周开孔的瓶盖(直径30 mm,高13 mm)中,制作人工斑块(各微生境所需具体数量见下文),以模拟其野外丛生状态;然后将斑块放入相对湿度80%、温度25℃、光照时间9 h的培养箱内培养3 d,使其恢复生长[24]。培养过程中,每天加水1次,每次2 mm,使用滴管施加在苔藓丛内。
1.3.1脱水过程中叶绿素荧光参数测定
用已培养3 d的向阳和荫蔽生境毛尖紫萼藓人工斑块进行脱水处理。首先将人工斑块瓶盖底部滤纸抽去,调整培养箱相对湿度为60%、温度为25℃,在培养箱内进行脱水[10]。每隔1.5 h对样品重复测定一次叶绿素荧光参数,共6次(7.5 h); 2组处理各5个重复,共10个人工斑块,各样品重复测定。
1.3.2光温复合处理下的叶绿素荧光参数测定
本研究依据毛尖紫萼藓生长旺盛期(3—4月)裸岩表面温度昼夜变化特点及向阳和荫蔽微生境光照和温度的实际差异,设置2个温度梯度及2个光照强度梯度(本研究所指温度和光照的高低均为2种生境下的相对值,用以体现2种生境间的微环境差异)。光照培养箱的光照强度设为50 μmol m-2s-1和250 μmol m-2s-1(光照9 h),白天温度设为20℃和30℃,夜晚温度设为15℃。本研究采用相对的低温-弱光组合及相对的高温-高光组合来模拟荫蔽生境和向阳裸岩生境的光温状况。培养时水分保持充足,实验连续开展12 d。每2 d重复测定一次叶绿素荧光参数,测定时间点统一为加水1 h后(当日18:00)。该实验要求样品来源一致,因此统一选择向阳裸岩生境藓类。2组处理各5个重复,共10个人工斑块,各样品重复测定。
1.3.3水光温交互处理下的叶绿素荧光参数测定
以温度和光照的低低组合和高高组合来模拟遮蔽和向阳2种微生境下温度和光照(9 h)的协同变化(即一个组合的单因素),与水分频次和水量梯度相结合开展3因子实验。模拟将水量梯度为2 mm/次(7 mL)和4 mm/次(14 mL)共2个处理,模拟降水频次梯度为1次/(1 d)、1次/(2 d)和1次/(3 d),模拟降水量分3次施加完毕,以避免单次施加造成流失。样品均为向阳裸岩生境藓类,连续开展12 d;分别在第6天(频次的最小公倍数)和第12天当日最后一次喷水1 h后开始测定(当日18:00)。每处理5个重复,共60个人工斑块,各样品重复测定。
1.4 叶绿素荧光的测定方法及相关参数计算
利用便携式调制叶绿素荧光仪PAM- 2100 (Walz, 德国)测定毛尖紫萼藓的叶绿素荧光参数,得出最大光化学效率Fv/Fm和实际光化学效率Yield (ΦPSII)。室内测定时关闭培养箱光源,野外原位测定时用遮荫布遮住藓类斑块,均进行30 min暗适应,随后在波峰为650 mm 强度0.1 μmol m-2s-1的红光下测得初始荧光(F0);在波峰为650 mm强度10000 μmol m-2s-1的饱和脉冲处理0.8 s后测得最大荧光(Fm);在光照强度为250 μmol m-2s-1的光化光下测定稳态荧光(Ft);待Ft稳定后打开一次饱和脉冲光测得光适应后的最大荧光(F′m)[10,17]。
根据以上参数计算PSII的最大光化学效率:
Fv/Fm=(Fm-F0)/Fm
(1)
光适应下PSII的实际光化学效率:
ΦPSII=(F′m-Ft)/F′m
(2)
式中,Fv为暗适应下叶片给光的最大可变荧光值,ΦPSII即Yield。
1.5 统计分析
对不同处理(光温、处理时间等)间的叶绿素荧光参数(Fv/Fm和Yield)进行单因素方差分析(One-way ANOVA),采用LSD法进行多重比较(α=0.05);采用t-检验分析2个微生境间各指标在不同处理间的差异性。用一般线性模型(GLM)中的3因素方差分析解析不同光-温、降水量、降水频次及其交互作用对毛尖紫萼藓叶绿素荧光参数的影响,用单因素方差分析进行处理间的多重比较(α=0.05)。使用SPSS 19.0软件完成所有统计分析,并用Excel 2016对数据做图。图中数据均以平均值±SE表示。
2 结果与分析
2.1 野外原位脱水过程中毛尖紫萼藓Fv/Fm和Yield的变化
2种生境毛尖紫萼藓在野外原位饱和吸水后的脱水过程中(图1),除荫蔽生境脱水4 h与5 h间有显著差异外(P<0.05),2生境Fv/Fm和Yield (ΦPS II)在脱水1—5 h内均无显著差异,且在脱水24 h后接近0。荫蔽生境下毛尖紫萼藓的光合活性均高于向阳生境,且在前4 h呈显著差异(P<0.05)。可见,在环境条件相对较好的荫蔽生境中,毛尖紫萼藓具有更高的光合活性,而向阳生境毛尖紫萼藓光合活性始终较低。

图1 2种微生境毛尖紫萼藓原位脱水过程中光化学效率的差异性Fig.1 Differences in photochemical efficiency of Grimmia pilifera in two habitats during in situ dehydration不同大写字母表示向阳生境毛尖紫萼藓脱水过程中光化学效率差异显著(P < 0.05); 不同小写字母表示荫蔽生境毛尖紫萼藓脱水过程中光化学效率差异显著(P < 0.05); *和ns表示各时间点各指标在2生境间差异显著(P < 0.05)和不显著(P > 0.05)
2.2 室内相同环境下毛尖紫萼藓脱水过程中Fv/Fm和Yield的变化
在室内脱水过程中(图2),荫蔽生境毛尖紫萼藓Fv/Fm在前3 h保持较高值,但第4.5小时急剧下降(P<0.05);向阳生境在前4.5 h均保持较高且稳定的Fv/Fm,至第6小时才显著下降(P<0.05)。在1.5—6小时之间,两生境间均有显著差异,其中在第1.5和第3小时为荫蔽生境显著(P<0.05)高于向阳生境,而在第4.5和第6小时为向阳生境显著(P<0.05)高于荫蔽生境。对于Yield,在前3 h间除向阳生境在第1.5小时有显著降低外(P<0.05),2生境均保持较高值;在第4.5 h,2生境藓类Yield均急剧下降(P<0.05),但此时和随后的第6小时,向阳生境的Yield显著高于荫蔽生境(P<0.05)。可见,尽管脱水前期荫蔽生境毛尖紫萼藓Fv/Fm及Yield有时高于向阳生境,但荫蔽生境藓类植物有效光合活性的维持时间却显著短于向阳生境。
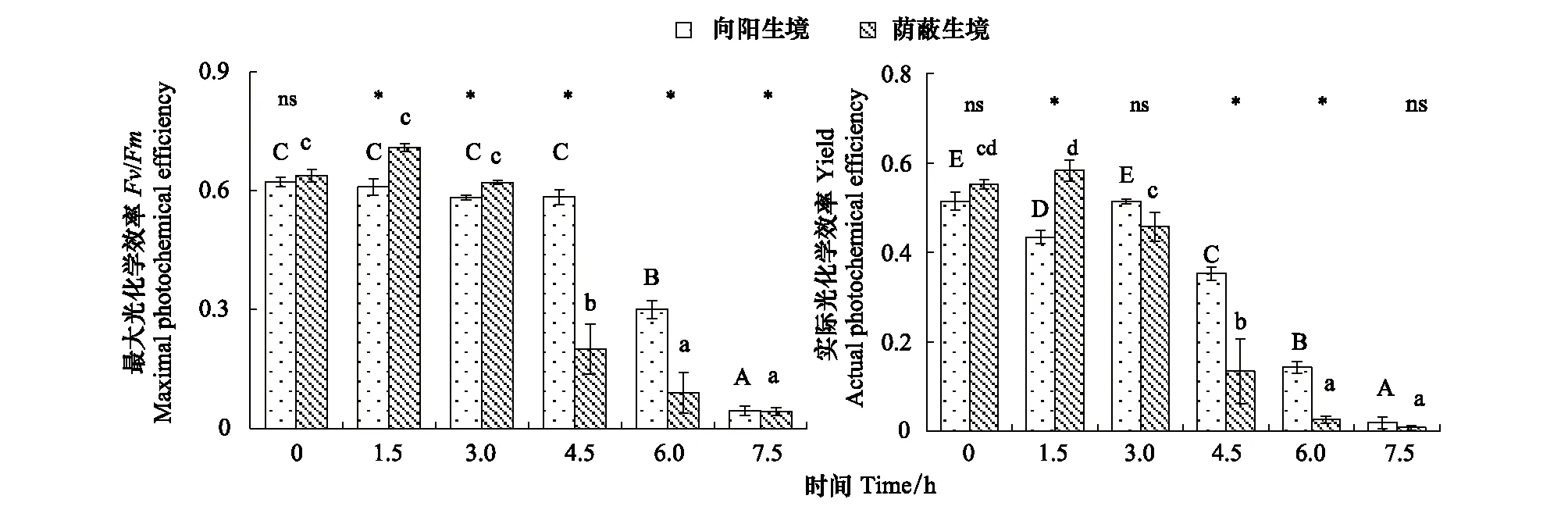
图2 2种微生境毛尖紫萼藓在室内相同环境脱水过程中光化学效率的差异性Fig.2 Differences in photochemical efficiency of Grimmia pilifera in two habitats during in door dehydration under the same environmental condition
2.3 模拟微生境中光照和温度变化对毛尖紫萼藓Fv/Fm和Yield的影响
由图3可知,弱光低温处理下毛尖紫萼藓Fv/Fm和Yield高于高光高温处理,且随着处理时间的延长,这种差异更加显著。处理时间对高光高温条件下毛尖紫萼藓Fv/Fm和Yield有显著影响(P<0.05),表现为处理时间越长光合活性越低:Fv/Fm从第4天开始下降,第6—12天均显著低于初始值,且两生境有显著差异(P<0.05)。Yield的变化趋势与Fv/Fm类似,只是在第10天时又稍增高;尽管如此,2生境Yield在第4—12天均有显著差异(P<0.05)。处理时间对弱光低温组的光化学效率无显著影响。由此可见,高光高温会限制毛尖紫萼藓的光合生理活性,而弱光低温则相对有利。

图3 不同处理时间弱光低温和高光高温条件下毛尖紫萼藓光化学效率的差异性Fig.3 Differences in photochemical efficiency of Grimmia pilifera under low-light and low-temperature, and high-light and high-temperature in different treatment times
2.4 模拟微生境中水光温复合变化对毛尖紫萼藓Fv/Fm和Yield的影响
3因素方差分析表明,光温和降水频次对毛尖紫萼藓光化学效率均有显著和极显著影响(表1);且随着时间的延长,二者对Yield的交互作用也呈显著趋势。模拟降水量及其与降水频次交互作用在前期对毛尖紫萼藓Fv/Fm有显著影响,但后期无显著影响;光温和降水量在第12天时对Yield呈现出显著的交互作用。此外,3个因素对2个测定时间毛尖紫萼藓的Yield均存在极显著的交互作用。
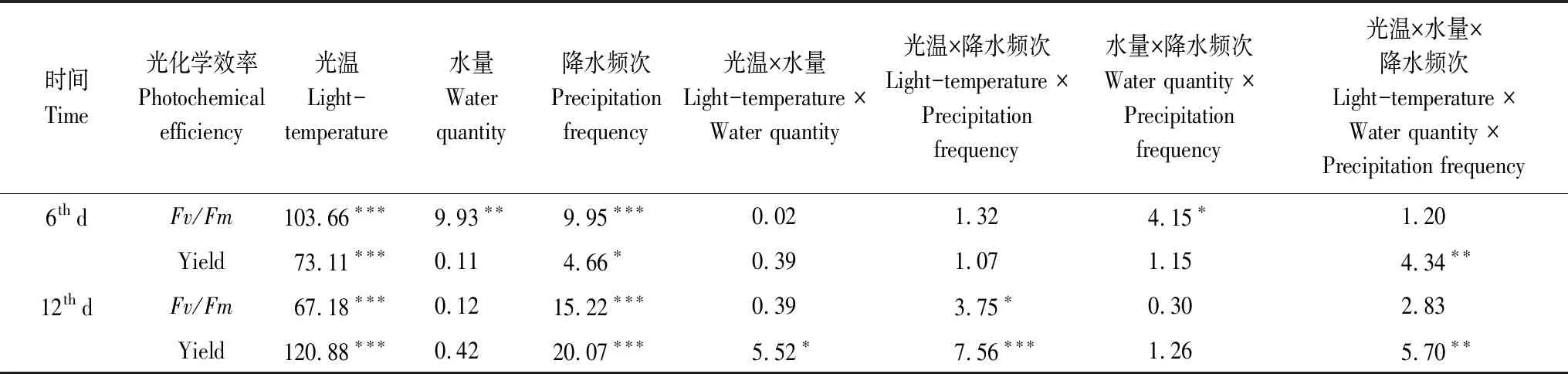
表1 光温、降水量及降水频次对毛尖紫萼藓光化学效率影响的3因素方差分析(F值)
由图4可知,对于不同模拟降水频次,两个测定时间点弱光低温组Fv/Fm在不同降水量条件下呈现不同的变化趋势,其中低水量时1次/(2 d)的Fv/Fm显著高于1次/(3 d),而高水量组Fv/Fm则为呈现1次/(1 d)>1次/(2 d)>1次/(3 d)的趋势;在高光高温条件下,低水量时1次/(2 d)的Fv/Fm高于1次/(1 d)和1次/(3 d),而高水量组Fv/Fm在第6天时呈现1次/(1 d)>1次/(2 d)>1次/(3 d)的趋势,但在第12天则呈1次/(2 d)显著高于1次/(1 d)和1次/(3 d)的趋势。毛尖紫萼藓Yield的变化趋势与Fv/Fm不完全相同。在弱光低温组,Yield在不同降水量条件下也呈现不同的变化趋势;其中在第6天时,低水量、1次/(3 d)的Yield显著低于1次/(1 d)和1次/(2 d),而高水量组Yield在3个降水频次间无显著差异;在第12天时,低水量、1次/(2 d)的Yield明显高于其他两个频次,而高水量Yield则随降水频次减小而显著降低;在高光高温条件下,第6天时低水量Yield在3个降水频次间无显著差异,而高水量条件下则以1次/(2 d)最高;而在,第6天时,无论水量多少,均以1次/(2 d)的Yield最高,以1次/(3 d)最低。
此外,无论哪种降水频次,高光高温条件下毛尖紫萼藓光合活性总体上低于弱光低温条件。可见,水分、温度和光照的复合处理对毛尖紫萼藓叶绿素荧光特性有显著影响,不同因素的影响大小有一定差异性,毛尖紫萼藓在弱光低温条件始终比高光高温条件活性更高,且中等频次模拟降水对促进毛尖紫萼藓生长、生理及室内培养更具优势。

图4 不同处理时间不同水光温处理对毛尖紫萼藓的光合效率的影响Fig.4 Effects of different water-light-temperature treatments on photosynthesis efficiency of Grimmia pilifera in different times不同小写字母表示毛尖紫萼藓光化学效率在不同处理间差异显著(P < 0.05)
3 讨论
藓类植物微生境的差异主要体现在土壤含水量、土壤养分(石生藓等无土种类除外)、空气湿度、温度、光照强度和UVB辐射强度等方面,因此微生境是多种微环境因子的综合反映[17]。不同微生境对藓类植物生长、生理及繁殖有明显影响。Herrnstadt等对3种不同微生境下旱生真藓属植物Bryumdunense繁殖策略的研究表明,微生境差异决定了繁殖器官的类型和数量,且荫蔽生境下藓类有性繁殖现象更加明显[25]。对荒漠齿肋赤藓的研究表明,灌丛下藓类植物的盖度和生长状态要好于裸露地的藓类[26];在生理活性方面,裸露地高辐射、高温、高蒸发条件导致齿肋赤藓脯氨酸、可溶性糖等渗透调节物质累积和SOD、CAT及POD等抗氧化酶活性提高,以增强抗性;而灌丛的“湿岛效应”虽然减弱了干旱对齿肋赤藓造成的伤害,但却导致灌丛下藓类植物缺乏抗性锻炼而呈现较低的耐受性[17]。正如本研究发现,野外原生生境下荫蔽生境毛尖紫萼藓的光合活性显著高于向阳生境,但在相同环境下其光合活性对脱水的持续时间却低于向阳生境,体现了较弱的耐脱水性。该结论与不同微生境下齿肋赤藓生理活性差异的结论相吻合[17,19]。这表明,无论沙生的齿肋赤藓还是石生的毛尖紫萼藓,向阳/暴露生境植物体对干旱高温(水分亏缺)的适应性较强,而荫蔽生境植物体的适应性较弱[27-28]。尽管我们对耐旱藓类生理和光合活性对不同微生境的响应有了一定认识[26- 28],但还缺乏从具体的微环境因子角度解析这种差异的原因。
为探讨微生境中水光温各因子及其交互作用对毛尖紫萼藓光合活性的影响,在室内实验中设置不同水-光-温及其不同组合。结果显示,光-温高低及施水频次对毛尖紫萼藓叶绿素荧光特性的影响最大,而现有2个降水量梯度(2 mmm和4 mm)的影响较小。大量研究表明,降水能够增加藓类植株含水量,激活光合生理活性,但降水量过大又将加剧膜脂过氧化程度,影响叶片气孔导度及CO2扩散速率,从而降低光化学活性[29],不利于藓类植物生长[30]。张静等指出,齿肋赤藓在最适水分条件下光化学效率最高,而随着含水量逐渐增加,其光化学效率逐渐降低[31]。相反,低水分条件也无法满足藓类正常生长需求,会使细胞膜处于渗透胁迫状态,引发细胞质膜过氧化反应,威胁细胞膜正常代谢活动[29]。本研究发现,不同梯度的水分处理对毛尖紫萼藓光合活性有一定影响,但影响程度与模拟降水频次有关,随着处理时间的延长,1次/(2 d)的中等降水频次逐渐显现优势。在研究影响毛乌素沙地藓结皮野外定植发育的因素时发现,每隔2 d加水处理的藓结皮叶绿素a含量高于每1天加水和每6天加水处理[32]。许红梅等研究发现,随降水频次增加,齿肋赤藓叶绿素含量及叶绿素a/b呈现先升后降的趋势,而在干旱和完全湿润处理条件下则表现出黄化现象[33]。本研究结果也同样说明,施水量及降水频次过少或过多都不利于耐旱藓类生长,尤其是在高光高温环境下;小降水量使毛尖紫萼藓无法获得充足的水分,而频繁降水又使毛尖紫萼藓处于频繁脱复水状态,均会显著降低其再水化后的生理恢复能力[6]。
对于光照强度,尽管一些研究发现随着光强增加藓类植物净光合速率逐渐增加并趋于稳定[34],但光强增加也确实会增加藓类植物的氧损伤,且干旱敏感型藓类植物受影响程度更大。如对干旱敏感的小曲尾藓(Dicranellapalustris)在高光下导致氧损伤加剧,色素含量减少,而耐旱的拟山墙藓(Tortularuraliformis)光合色素含量和恢复能力均未受明显影响[35]。温度变化能强烈影响藓类植物的光合生理[11,13,36],但在藓类植物实际微生境中,白天的温度和光照通常是协同变化的,即向阳生境高光高温而荫蔽生境弱光低温。研究表明,高光高温条件同时存在,会抑制植物D1蛋白的修复,致使PSII反应中心失活[37];相比之下,弱光低温条件下植物则有较高的色素含量,有利于复水时光合活性快速恢复[38]。本研究中,相对于自然光和岩石表面真实温度,尽管2个光照-温度不属于胁迫范畴,但仍然显著影响了毛尖紫萼藓的光合活性;且随处理时间延长,毛尖紫萼藓的Fv/Fm和Yield均表现出高光高温条件低于弱光低温条件。该结论不一定反映了相对的“高光高温”破坏了毛尖紫萼藓细胞膜结构的完整性进而降低其光合效率,但却反向证实相对的“弱光低温”条件有利于毛尖紫萼藓生长和维持较高的光合活性。
环境因子间通常是相互影响的,并给植物生长和生理带来不同的效应。在温度和水分交互影响下,藓类植物往往通过失复水和渗透调节物质来维持细胞渗透压平衡[39]。当环境湿度较高时,藓类植物体内含水量和代谢水平升高,此时温度和光照对其影响较大[40];但当外界湿度过低时,水分供给满足不了藓类植物生长的需求而呈干燥休眠状态,此时温度和光照对藓类的影响会相对减弱[6]。在本研究中,“光温”组合及降水频次对毛尖紫萼藓光合活性影响最大,且在水分相对充足的情况下,光照和温度的影响作用更大,体现了多个环境因子之间的交互作用。藓类脱水过程实质是在光、温等因子影响下的水分胁迫过程,高温、强光会加速脱水进程。正常脱水过程中,藓类植物会逐渐降低生长代谢活动[2],并通过抗氧化酶来清除体内多余的活性氧和氧自由基[9,31],提高渗透调节物质以减少对细胞及其内部结构的损伤[41-43]。而一旦脱水进程在高温、高光或干燥条件下被加快,藓类植物体内蛋白质含量会迅速减少,细胞膜完整性被破坏,进而降低了PSII反应中心活性,严重降低复水后的光合恢复能力[1,6,44]。可见,水光温等因子的交互作用对藓类植物生理活性的影响是显著且十分复杂的。
本研究以荫蔽和向阳2种微生境为例,选择特定时间(3—4月)对应的水光温条件,探讨了毛尖紫萼藓叶绿素荧光特性的差异;因此,本研究不属于极端环境条件下(如极端低温、高温,强光,极端干旱等)的对比研究。尽管如此,本研究结果对于理解耐旱藓类脱水耐受机制仍然具有一定的科学意义。本研究发现,野外荫蔽生境毛尖紫萼藓脱水过程中的光合活性始终高于向阳生境,这印证了向阳生境强光和高温导致的频繁快速脱水显著抑制了植株光合活性。而在室内相同环境脱水过程中,荫蔽生境毛尖紫萼藓有效光合作用时间明显低于向阳生境。可见,长期隐蔽环境尽管给原生境毛尖紫萼藓带来更高的光合活性,但由于其没有经历长期快速脱水和干旱胁迫,其控制体内水分散失的能力较弱,体现出较弱的抗旱性。总之,本研究证实毛尖紫萼藓叶绿素荧光特性在不同微生境和水光温处理间有显著差异,但其在抗氧化酶系统、基因表达、光合系统恢复与细胞损伤等方面有何差异化响应尚待进一步探究,以充分揭示这一石生耐旱藓类模式物种对不同微生境的响应和适应机制。
4 结论
微生境差异引起的水光温变化对毛尖紫萼藓的光合活性有显著影响。荫蔽生境为毛尖紫萼藓提供了相对湿润、弱光和低温的环境,而向阳裸岩生境则具有干燥、高光和高温的特征。在野外原位条件下,荫蔽生境毛尖紫萼藓比向阳生境具有更高的光合活性,但在相同环境条件下前者的有效光合作用时间明显降低。向阳裸岩生境毛尖紫萼藓由于长期适应极端环境,具有更高的抗逆性。这表明,一旦荫蔽环境遭到破坏(如维管植物死亡、移除等等),该生境中的藓类植物将面临更严峻的生存考验。此外,弱光低温条件和中等模拟降水频次有利于毛尖紫萼藓的个体生长,对于其室内培养研究具有一定指示意义。
猜你喜欢
现代盐化工(2021年3期)2021-08-27
故事会(蓝版)(2020年7期)2020-07-21
食品与生物技术学报(2020年4期)2020-01-06
大众科学(2018年10期)2018-12-11
贵茶(2018年6期)2018-05-30
农产品市场周刊(2017年18期)2017-06-07
新东方英语·中学版(2017年1期)2017-02-25
小天使·一年级语数英综合(2016年11期)2016-11-28
新课程·下旬(2015年10期)2015-10-21
