
若尔盖高原湿地3种生境地下芽库特征及其差异
2021-04-16 07:33刘振亚刘耘硕王好才肖德荣
水土保持通报 2021年1期
王 娜, 夏 敏, 王 行, 刘振亚, 刘耘硕, 王好才, 肖德荣, 吕 梅
(1.西南林业大学 国家高原湿地研究中心/湿地学院, 云南 昆明 650224;2.温州大学 生命与环境科学学院, 浙江 温州 325035; 3.贵州省威宁县林业局, 贵州 威宁 553100)
地下芽库在维持植物群落的结构和功能,以及在受损植被的恢复中发挥着基础性作用[1-4]。研究发现,在未受破坏的高草草原中有超过99%地上植株来源于地下芽库[5],在受破坏的高草草原中也有80%~85%的地上植株来自地下芽库[6]。环境因子影响甚至决定着植物地下芽的形成和发展[7]。掌握地下芽库多样性特征及环境作用机制,对于保护和恢复植物多样性具有重要的理论价值。
当前,环境因子对植物地下芽库的影响正受到越来越多的关注[8]。水分是影响植物地下芽库类型和密度的关键因子[9]。植物芽库的数量随着降水量或土壤水分含量的增加而显著增加[10],土壤水分的亏缺可导致地下芽库密度的下降[11]或无明显影响[2];另外,土壤水分也对植物芽库的类型产生影响[12]。土壤养分是影响植物地下芽库建成和动态的另一重要因素[3]。适度的土壤养分,有利于植物芽库的形成和萌发,其中,增加土壤氮素可有效提高植物地下芽库的密度[13]。外界干扰对植物地下芽库也会产生影响[14]。放牧可显著增加植物地下芽库的总密度[15],或导致禾草类[16]、非禾本草本[17]植物芽库中芽的数量减少;随放牧强度的增加,根茎芽和鳞茎芽的密度显著降低,但芽库总密度保持不变[1]。通常情况下,植物地下芽库的密度随着海拔的增加而呈现增大的趋势[15]。可见,植物地下芽库的多样性与环境因子间有着极为复杂的关系。
在高寒地区,地下芽库对植物种群及多样性的维持发挥着重要作用[18]。在北美高寒草原,99%的地上植物茎来自地下芽库[10];在阿尔卑斯山,84%的高山草原以克隆植物为主导[19]。若尔盖高原湿地是中国“两屏三带”生态安全格局中“川滇生态屏障”的重要组成,具有重要的生物多样性与特有性保护价值,但其生态系统极为脆弱[20]。研究表明,中国高寒地区分布的大部分植物都具有较强的无性繁殖能力[21],其植物通过强大的克隆繁殖,在区域水土保持、生物多样性保护中发挥着不可替代的作用。若尔盖高原湿地生境的空间异质性高,生境间环境因子存在较大差异[22]。若尔盖高原湿地不同生境间的植物地下芽库多样性是否存在差异?影响这种差异的环境因子是什么?其影响途径如何?目前尚缺乏研究。
本研究选择若尔盖高原湿地的沼泽、沼泽化草甸和草甸3种生境(见图1),重点研究各生境下芽库的组成和密度,芽形态及生物量,芽生态化学计量特征,环境因子对植物芽密度、形态、生物量及生态化学计量的影响规律。通过研究,拟为若尔盖高原湿地植物多样性的保护和恢复提供一定的理论依据。

图1 若尔盖高原湿地典型生境类型
1 研究区概况
研究区地处青藏高原东北麓,横断山脉北段,四川省阿坝藏族羌族自治州北部,若尔盖县、红原县、玛曲县等境内,地理位置为北纬32°25′—34°6′,东经102°4′—103°8′,平均海拔3 430 m,属高原寒温带湿润气候,年均温0.7 ℃。年降水量660~750 mm,86%降雨量集中于4—10月,相对湿度64%~73%,年均蒸发量1 232 mm,日照2 389 h。
随地形及水文条件的差异,若尔盖高原湿地在空间形成了沼泽、沼泽化草甸和草甸等主体生境类型[23]。若尔盖高原湿地植物物种分布较为丰富,主要有木里苔草(Carexmuliensis)、藏嵩草(Kobresiatibetica)、华扁穗草(Blysmussinocompressus),以及毛果苔草(Carexlasiocarpa)、乌拉苔草(Carexmeyeriana)、四川嵩草(Kobresiasetchwanensis)、线叶嵩草(Kobresiacapillifolia)、无脉苔草(Carexennrvis)、垂穗披碱草(Elymusnutans)等。
2 材料与方法
2.1 样品采集
2018年5—6月和2019年5—6月,在植物的生长初期,分别在以木里苔草(Carexmuliensis)、藏嵩草(Kobresiatibetica)、华扁穗草(Blysmussinocompressus)为优势物种的沼泽(MA)、沼泽化草甸(MM)和草甸(ME)3种生境设置1 m×1 m样方共计51个,其中,沼泽样方11个,沼泽化草甸样方17个,草甸样方23个。分别调查样方内3种优势物种植物高度、盖度、生物量等(表1)。在每个样方,用剪刀齐地刈割25 cm×25 cm[24]面积内的地上植物并装袋,以测定植物地上生物量。高原湿地植物根系集中分布于土壤表层[25],因此,用小刀和铁铲将刈割地上植物后的20 cm的地下土块(地下植物及土壤)取出,装袋带回实验室以测定植物芽库和地下生物量。用定深泥炭钻(型号:荷兰Eijkelkamp 0 423SA)采集0—20 cm混合土壤样品,用环刀(D:37 mm,H:50 mm)对泥炭钻中的土壤进行取样,剩余混合土壤装入自封袋带回实验室,以测定土壤理化指标。
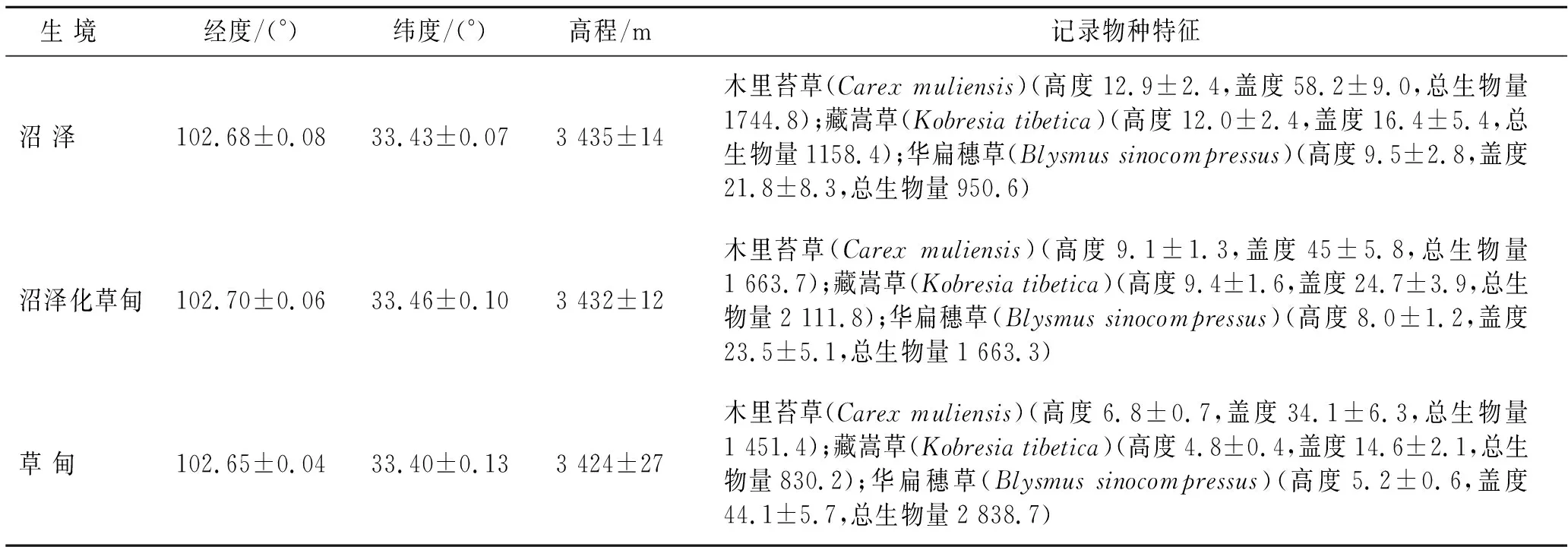
表1 3种生境中优势植物物种的高度、盖度及生物量特征
2.2 室内分析
2.2.1 芽指标的测定 依据芽在母株上的位置[26],本研究将地下芽的类型分为水平生长根茎顶芽、向上生长根茎顶芽、根茎节芽和分蘖节芽4种类型。
在室内,通过手工初步去除用于芽库的研究样品的泥土,然后放置清水池中洗净以取出植物样品(尽量避免伤害根茎和芽之间的连接)。将取出的植物样品放置实验室晾干,采用放大镜(放大10倍)对不同生境的植物芽和芽痕进行观察,并分别记录芽的类型及数量。在此基础上,计算植物芽库的总密度(芽/m2),以及4种类型芽的分密度(芽/m2)[26]。用游标卡尺测定地下芽的长度(mm)和直径(mm),并计算芽长/直径比等形态指标。用小刀将植物芽从根茎上取下,采用烘干称重法(65 ℃,48 h),测定地下芽的生物量[26]。采用H2SO4-H2O2消煮法,测定地下芽的全氮、全磷等化学计量指标。
2.2.2 土壤理化指标的测定 土壤含水率、pH值参照《土壤农化分析》进行测定。采用H2SO4-H2O2消煮法,测定土壤全氮、全磷。采用1 mol/L氯化钾溶液浸提法,测定土壤速效氮。采用M3浸提法,测定土壤的速效磷。采用酸洗法,测定土壤有机碳含量。通过公式(1)计算土壤容重(SBD)[27]:
SBD=0.857 2e-0.033 1OC+0.744 6e-0.002 7OC
(1)
式中:SBD为土壤容重(g/cm3);OC为土壤有机碳含量(g/kg)。
2.3 数据统计与分析
采用SPSS 25.0的单因素方差分析(one-way ANOVA)法,对3种生境植物地下芽库的数量组成、形态及生物量,以及芽化学计量指标等进行差异性检验。对不符合单因素方差齐性检验的数据进行lg(x+1)的处理,显著性水平设定为α=0.05。通过Pearson相关性分析法,研究地下芽库的密度、形态及生物量,以及化学计量学指标与土壤的理化指标,采样的地理位置(经纬度、高程)、大气温度、降水,以及植物盖度、生物量的相关性。
在上述基础,进一步选取与芽库密度、形态、生物量及芽化学计量存在着显著相关性的指标,通过Stepwise回归分析方法再进一步筛选影响芽库特征的关键因子。使用R语言3.6.3版本中的Agricolae软件包,对影响植物芽库特征的关键因子进行路径分析,最终研究直接影响因子对植物芽库作用的路径和贡献率。
3 结果与分析
3.1 不同生境的土壤理化特征
若尔盖高原湿地3种生境的土壤理化性质存在差异(表2)。沼泽的土壤水分、全氮、全磷、速效氮、速效磷含量分别显著高于沼泽化草甸和草甸的对应指标,沼泽和沼泽化草甸的土壤有机质含量显著高于草甸的土壤有机质含量,而沼泽和沼泽化草甸的土壤有机质含量无显著差异,沼泽化草甸的土壤水分、全氮、速效氮、有机质含量分别显著高于草甸的对应指标,沼泽化草甸和草甸的土壤全磷、速效磷含量均无显著差异,沼泽和沼泽化草甸的土壤容重分别显著低于草甸的土壤容重,而沼泽和沼泽化草甸的土壤容重无显著差异。3种生境土壤的pH值无显著差异。
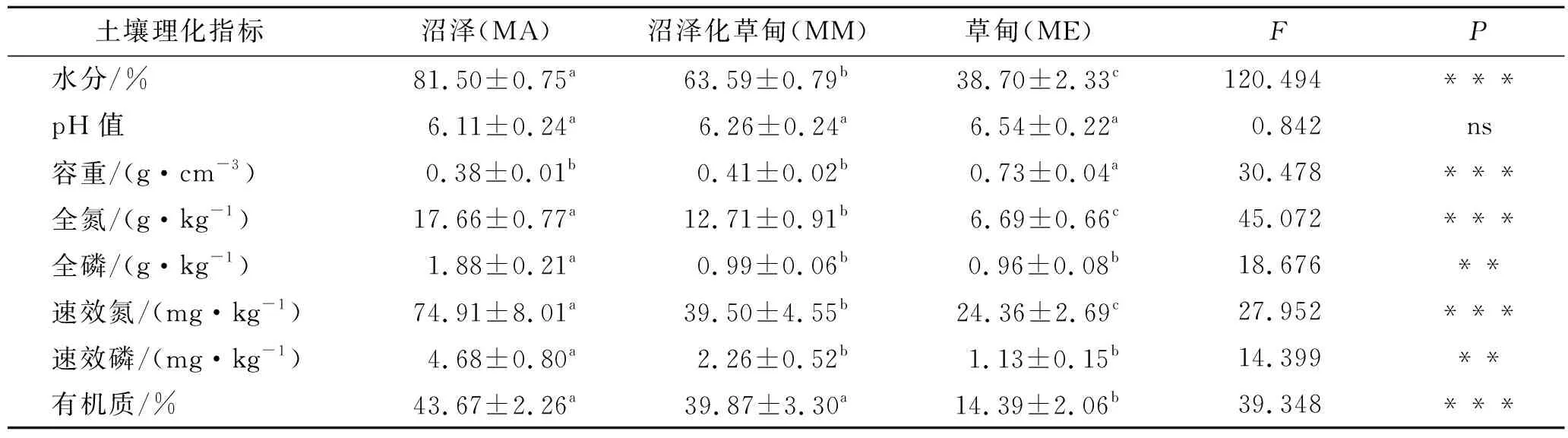
表2 若尔盖高原湿地3种生境的土壤理化特征
3.2 芽库组成及密度
3种生境的芽库均由水平生长根茎顶芽、向上生长根茎顶芽、根茎节芽、分蘖节芽等4种类型组成,但组成比例不同(图2a)。沼泽的芽库以水平生长根茎顶芽为主(34.01%),沼泽化草甸和草甸的芽库则分别以根茎节芽、分蘖节芽为主(48.50%,60.43%)。沼泽的芽库总密度(1 497±77 个/m2)显著高于沼泽化草甸和草甸的芽库总密度(691±27 个/m2,294±32 个/m2),后两者间同样存在显著差异。
沼泽的芽库水平生长根茎顶芽密度为(509±100 个/m2),显著高于沼泽化草甸和草甸的芽库水平生长根茎顶芽密度(126±31 个/m2,40±12 个/m2),后两者间无显著差异。沼泽的芽库向上生长根茎顶芽密度(304±50 个/m2)显著高于沼泽化草甸和草甸的芽库向上生长根茎顶芽芽密度(93±21 个/m2,35±11 个/m2),后两间无显著差异。沼泽和沼泽化草甸的芽库根茎节芽密度(441±70 个/m2,335±45 个/m2)间无显著差异,两者均显著高于草甸的芽库根茎节芽密度(40±12 个/m2)。3种生境的芽库分蘖节芽密度均无显著差异。
3.3 芽的形态及生物量
沼泽的芽库的芽长度(24.13±2.52 mm)显著大于沼泽化草甸和草甸的芽库的芽长度(13.50±1.84 mm,8.97±0.64 mm),后两者间同样存在显著差异。沼泽和沼泽化草甸芽库的芽直径(1.50±0.16 mm,1.26±0.09 mm)显著大于草甸的芽库的芽直径(0.86±0.06 mm),沼泽和沼泽化草甸芽库的芽直径间无显著差异(详见图2b)。沼泽芽库的芽长/直径比(18.12±2.67)显著高于沼泽化草甸和草甸芽库的芽长/直径比(11.83±1.89,11.93±1.57),后两者间无显著差异(图2b)。沼泽芽库的芽生物量(9.55±1.14 g/m2)显著高于沼泽化草甸和草甸芽库的芽生物量(5.81±0.53 g/m2,4.48±0.86 g/m2),后两者间无显著差异(图2c)。
3.4 芽的生态化学计量
沼泽芽库的全氮含量(36.63±2.23 g/kg)显著高于沼泽化草甸和草甸芽库的全氮含量(23.16±1.44 g/kg,16.18±1.27 g/kg),后两者同样存在显著性差异。沼泽,沼泽化草甸和草甸芽库的全磷含量分别为(3.77±0.32 g/kg,3.11±0.55 g/kg,2.48±0.32 g/kg),三者间无显著差异。沼泽,沼泽化草甸和草甸芽库的氮/磷比分别为(10.31±0.9,16.55±4.9,10.35±1.8),三者间无显著差异(图2d)。
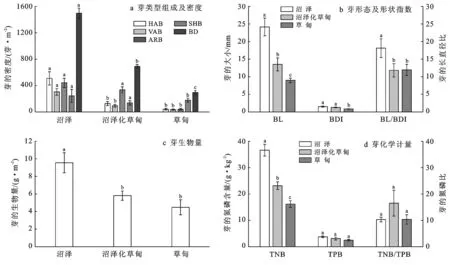
注:①HAB为水平生长根茎顶芽; VAB为向上生长根茎顶芽; ARB为根茎节芽; SHB为分蘖节芽; BD为芽密度; BL为芽长度; BDI为芽直径; BL/BDI为芽长/芽直径; TNB为芽全氮; TPB为芽全磷; TNB/TPB为芽氮磷比; ②不同小写字母代表地下芽库各指标在不同生境类型下p<0.05水平差异显著。
3.5 芽库的影响因素
芽库的各项指标与地理位置(经纬度、高程)、大气温度、降水无显著相关关系。芽库的氮/磷比与木里苔草和华扁穗草的地上生物量有正相关性,芽长度与木里苔草的盖度有正相关关系,但并不显著,其他芽库的指标与3种优势植物的盖度、生物量均无显著相关性。芽库的总密度、长度、直径、全氮含量与土壤水分、土壤全氮、土壤速效氮等理化指标均呈显著正相关关系,与土壤容重均呈负相关关系,与土壤的pH值无显著相关关系。芽库的生物量与土壤水分、全氮、全磷等理化指标呈正相关关系,与土壤容重、有机质、pH值无显著相关关系。芽库长/直径比与土壤全磷呈正相关关系,与其他土壤理化指标无显著相关关系。芽的全磷含量与土壤速效氮呈正相关关系,与其他土壤理化指标无显著相关关系。芽的氮/磷比与土壤理化指标无显著相关关系(表3)。
土壤水分、速效氮和全磷是影响植物芽库密度、生物量、形态和生态化学计量的直接因子(图3)。土壤水分直接影响芽的密度、直径、生物量和总氮含量,其中对芽库总密度的影响最大(其通径系数为0.916)。土壤速效氮、全磷是直接影响芽库的密度、芽形态(芽长、芽直径)以及芽总氮含量的直接因子,其中,土壤速效氮、土壤全磷对芽的长度的影响最大(其通径系数分别为0.447,0.340)。
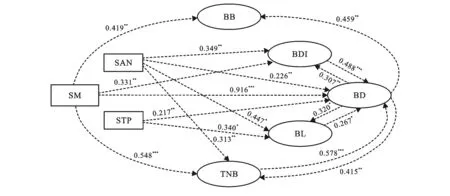
注:①SM为土壤水分;STP为土壤全磷;SAN为土壤速效氮;BD为芽密度;BL为芽长度;BDI为芽直径;BB为芽生物量;TNB为芽全氮。②箭头旁边的数字是标准化的路径系数,表示关系的强度; ③*表示p<0.05;**表示p<0.01;***表示p<0.001。
4 讨论与结论
本研究表明,若尔盖高原湿地3种生境下植物芽库的组成、形态及生态化学计量等存在差异,这与其所处环境中土壤的水分、养分(全氮、全磷等)等环境因子密切相关(表3)。通常,植物地下芽的产生、生长与存活受土壤水分的制约[28],充足和稳定的水分供给能促进植物地下芽的形成和发育[29],并显著提高植物芽在旱季的存活率[30]。本研究中的沼泽常年淹水,其土壤能为植物芽的生长与发育提供较为稳定的水资源,促进了沼泽植物地下芽的形成、生长和发育,并有利于地下芽在若尔盖高原地区旱季的存活。因此,沼泽生境植物芽库的存量相对较大、密度最高。沼泽化草甸和草甸生境的土壤水分供给条件较沼泽次之,这对植物地下芽库的形成、生长和发育较为不利,进而导致植物芽库的存量小、密度低。这与Ding等[31]研究结果相似。另外,较高的土壤水分能促进根茎芽的发育[31],而土壤水分的亏缺对分蘖节芽的形成有利[26]。因此,土壤水分的差异也是导致若尔盖沼泽和沼泽化草甸以根茎芽为主、而草甸以分蘖节芽为主的重要因子。这与张继涛等[26]对羊草地下芽库组成与环境因子关系的研究结果相似。
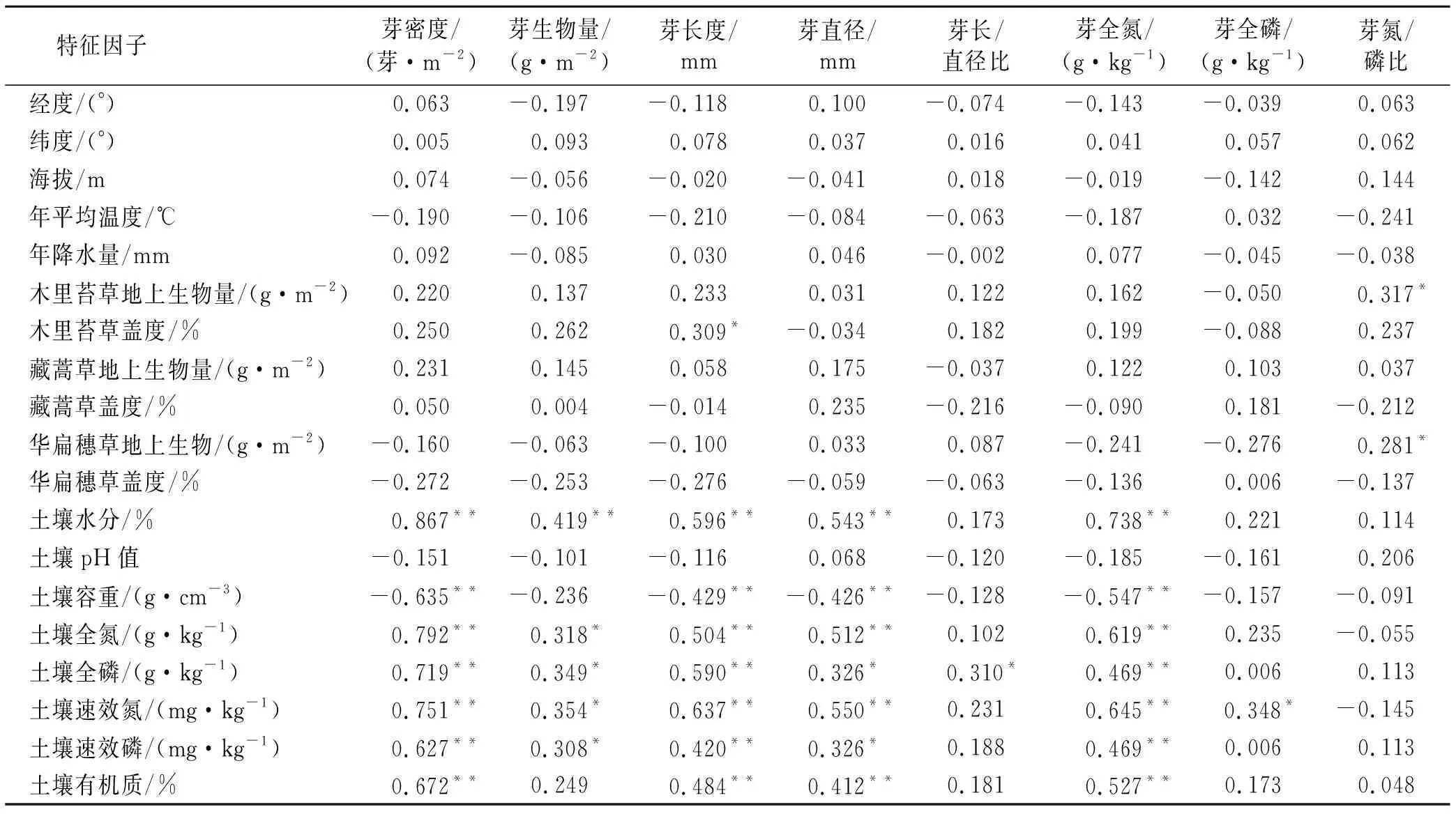
表3 芽库相关指标与地理位置、植物和土壤等环境因子的相关性分析
芽库存量与土壤养分含量呈显著正相关[4]。适宜的土壤养分(碳水化合物、全氮、全磷等营养元素)有利于植物地下芽的生命活动[4,8],进而促进植物芽形成和萌发[7]。本研究中,沼泽生境的土壤养分含量(全氮、全磷、速效氮等)最高,可有效地促进植物芽的产生和生长[16],这也是导致其芽密度最高的重要环境因子。而草甸生境的土壤养分含量最低,这可能对植物芽的产生与发育产生一定的抑制,从而使得该生境的芽密度最低。沼泽化草甸土壤养分条件介于沼泽与草甸之间,故其芽密度低于沼泽生境而高于草甸生境。适宜的土壤水分和养分有利于植物芽的生长[32],从而对植物地下芽的形态特征和生物量产生影响。本研究中,沼泽植物地下芽的直径、芽长等指标最高,其次为沼泽化草甸,最小的为草甸。这同样与土壤水分和养分含量密切相关(表3)。沼泽的土壤水分和养分含量最高,有利于植物地下芽的生长;草甸土壤水分和养分含量最低,不利于地下芽的生长;而沼泽化草甸土壤水分和养分介于两者间,从而导致其芽直径和长度低于沼泽但高于草甸生境。但是,3种生境芽的形状指数(长/直径比)与环境因子无显著相关关系,这可能是植物对差异环境的适应有关[33],相关调控机制有待进一步研究。
植物体内的氮、磷含量不仅与其长期进化过程中的适应机制有关,还与植物生长速率和养分利用效率密切相关[34]。环境因子通过影响植物的生长和养分的利用效率,进而对植物的化学计量指标产生影响[35]。不同水分和养分条件下,植物对于氮、磷的利用效率存在差异,影响着植物体的养分积累[36-37]。本研究中,3种生境植物地下芽的全氮含量存在显著差异(图2d),并与土壤水分和养分等相关指标存在显著相关关系(表3),这可能与沼泽适宜的土壤水分和养分条件促进植物体对氮的利用以及在芽部的积累密切相关。但是,3种生境下的地下芽的氮/磷比等生态化学计量指标间无显著差异(图2d),并与土壤环境因子无显著相关关系(表3),其对不同生境适应的调控机制有待深入研究。
地下芽的形成与发育主要通过与母株的生理整合来获得营养[38],芽库的建成同样依赖于母株自身的生理特点[39]。在湿地生态系统中,地下芽库与地上植物的相似性可达20%~67%[40]。本研究中优势植物主要为木里苔草、藏蒿草、华扁穗草,三者的优势度随着生境不同而存在差异。优势植物在稳定芽库数量方面发挥着基础性作用,甚至不受物种组成的影响[38]。因此,本研究中3种优势植物的芽库随着生境的不同而存在差异,这体现了该区域湿地植物的克隆繁殖策略与生长环境相关。本研究中的植物芽库密度、生物量及形态指标与3种优势植物的盖度、生物量等指标基本无显著相关关系,这进一步表明优势植物对芽库数量的稳定具有决定性作用,而与物种组成关系不显著。但是,由于本研究对地下芽的来源植物未准确判定,这为深入探讨地下芽库特征与地上植物多样性组成关系带来不确定性。高原湿地植物多样性丰富,生境的空间异质性极高,地上植物多样性、外界环境及其两者对地下芽库的协同作用机制有待于深入研究。
通过本研究,土壤水分、速效氮和全磷是影响植物地下芽密度、形态、生物量和生态化学计量指标的直接因子(图3)。土壤水分对芽库总密度,芽的生物量、直径和全氮均有极高的解释度,其中,对芽库总密度的解释度超过90%。研究表明土壤水分是影响若尔盖高原湿地植物地下芽库多样性的关键因子,维护和恢复湿地水环境是保护和恢复该区域湿地植物多样性的基础。
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
农业工程学报(2022年5期)2022-06-22
水土保持学报(2022年3期)2022-05-26
现代农村科技(2021年11期)2021-11-05
海外文摘·文学版(2021年3期)2021-05-19
活力(2019年21期)2019-04-01
中国水土保持科学(2017年1期)2017-04-18
剑南文学(2016年11期)2016-08-22
时代英语·高三(2014年5期)2014-08-26
