
不同类型杉木人工林林下草本植物多样性特征
2021-04-25 08:24张涵丹康希睿邵文豪张建锋刘学全陈光才
生态学报 2021年6期
张涵丹,康希睿,邵文豪,杨 旭,张建锋,刘学全,陈光才,*
1 中国林业科学研究院亚热带林业研究所,杭州 311400 2 湖北省林业科学研究院湖北大巴山森林生态系统国家定位观测研究站,武汉 430075
林下草本植物是森林生态系统的重要组成部分,其通过一系列的生理生化活动,不断改变林下微环境,对提升土壤肥力[1]、保持水土[2]、增加森林生态系统多样性[3]、维持生态系统稳定[4]、促进生态系统的演替和发展[5- 6]起着重要作用,其生物多样性的变化亦是衡量森林群落结构和功能的重要指标[7- 9]。然而,草本植物的多样性受多种因素影响[10- 16]。如,赵建生等[15]指出,华北落叶松林下草本植物多样性随林分密度的增大而减小;朱媛君等[11]在对杨树林的研究中得到相似结果,并指出,林分死亡率、平均枝下高、平均胸径和林分密度主要影响草本层的物种丰富度和Shannon-Wiener指数,而对于Simpson指数与均匀度指数则无明显影响;高艳鹏等[10]对晋西黄土高原典型林地的研究又进一步指出,林下草本多样性具有林地差异性,且混交林样地较纯林样地有更高的草本丰富度[14]。海拔高度[12]、坡度[17]、坡向[13]、光照[18]、人为活动等[16]亦是影响植物多样性的重要因素。此外,亦有研究表明林下草本植被的多样性变化与所处林地的演替周期有关[19]。丰富的生物多样性不仅能维持生态系统的稳定,更会促进生态系统功能的优化。研究表明,在干旱地区,植物群落物种丰富度的增加能显著提高生态系统的抗旱能力[7]。Huang等[8]通过对亚热带森林群落连续5年的观测发现,生物多样性的增加不仅能增加地上生物量,还能缓解气候变化。因此,研究林下草本多样性及其物种组成对于了解森林生态系统的稳定性及功能多样性至关重要,是林地合理经营和可持续发展的重要依据。
杉木是我国亚热带地区的特有树种,亦是重要的用材林树种,据统计,其人工林面积占亚热带人工林总面积的60—80%,在我国森林资源和亚热带森林生态系统中占有重要地位[20]。但因人工林树种单一,种植密度过大,常造成林地退化、生产力下降、病虫害频发等现象[21- 23]。林下草本植被作为人工林生态系统的重要组成部分也会因林地地力退化而受到影响,进而直接威胁人工林的可持续发展。因而,对于人工林生态系统中林下植被多样性变化的研究至关重要。此外,由于人工纯林出现的各种弊端,林地的阔叶化改造逐渐受到重视[23-24],而混交树种的选择、混交林树种配置以及混交程度的确定等是实现森林群落可持续经营需要解决的关键问题。研究以湖北省西北部九华山林场内不同树种配置的杉木人工林为研究对象,通过分析林下草本植物物种组成及其多样性之间的差异,探讨林地树种配置对草本植物多样性的影响,以期为杉木人工林的合理经营提供理论依据。
1 研究区概况
研究区位于湖北省十堰市竹山县九华山林场(100°08′—110°12′E,32°01′—32°06′N),海拔600—1311 m,年平均降水量1064—1375 mm,年均气温12.9℃,雨热同期,四季分明,属于北亚热带湿润气候区。该地区土壤以山地黄棕壤、黄棕壤、黄棕壤性土和山地黄棕壤性土为主,其中,黄棕壤和黄棕壤性土主要分布在海拔800 m以下的低山;山地黄棕壤和山地黄棕壤性土主要分布在海拔1200 m的高山。试验林地主要分布在海拔900—1200 m的山坡上,属于中山地貌,地形复杂多样,坡度介于14°—33°之间,土层较薄。九华山林场于1989年人工种植了杉木(Cunninghamialanceolata)、檫木(Sassafrastzumu)、亮叶桦(Betulaluminifera)、枫香(Liquidambarformosana)、马尾松(Pinusmassoniana)、栎属(Quercus)等常见树种,生长至今,形成了郁闭度高达0.78—0.9的人工林。其余乔木树种有华山松(Pinusarmandii)、微毛樱桃(Cerasusclarofolia)、山杨(Populusdavidiana)等,主要草本植物有高粱泡(Rubuslambertianus)、金星蕨(Parathelypterisglanduligera)、鱼腥草(Houttuyniacordata)、荩草(Arthraxonhispidus)、山马兰(Kalimerislautureana)等。
2 研究方法
2.1 样地设置与群落调查
2019 年6月,在全面踏查的基础上,选择相同林龄,不同树种配置的杉木(Cunninghamialanceolata)人工林作为研究对象,包括杉木纯林(I)、杉木-檫木(Sassafrastzumu)混交林(II)和杉木-檫木-亮叶桦(Betulaluminifera)混交林(III)3种林地类型。每个类型各设置3个面积为20 m×20 m的标准样地,所有样地相互间隔超过30 m。每个样地内按对角线法设置5个1 m×1 m的草本样方,共计设置9个乔木样地,45个草本样方。记录每个标准样地的海拔、坡度、坡向等地形参数(表1),对标准样地内的林木进行每木检尺,记录树高、胸径、冠幅等(表2)。记录草本样方内每个物种的高度、盖度、多度等,并采用收获法剪取样方内所有物种的地上部分,带回实验室烘干测算生物量。30年间,因管理需求,九华山林场对各林地进行了不同程度的人为疏伐,如表1所示,为调整林分密度,加速林木生长,杉木纯林样地在2015年进行了一次生长伐;在2017年又对所有样地进行了一次生态疏伐。
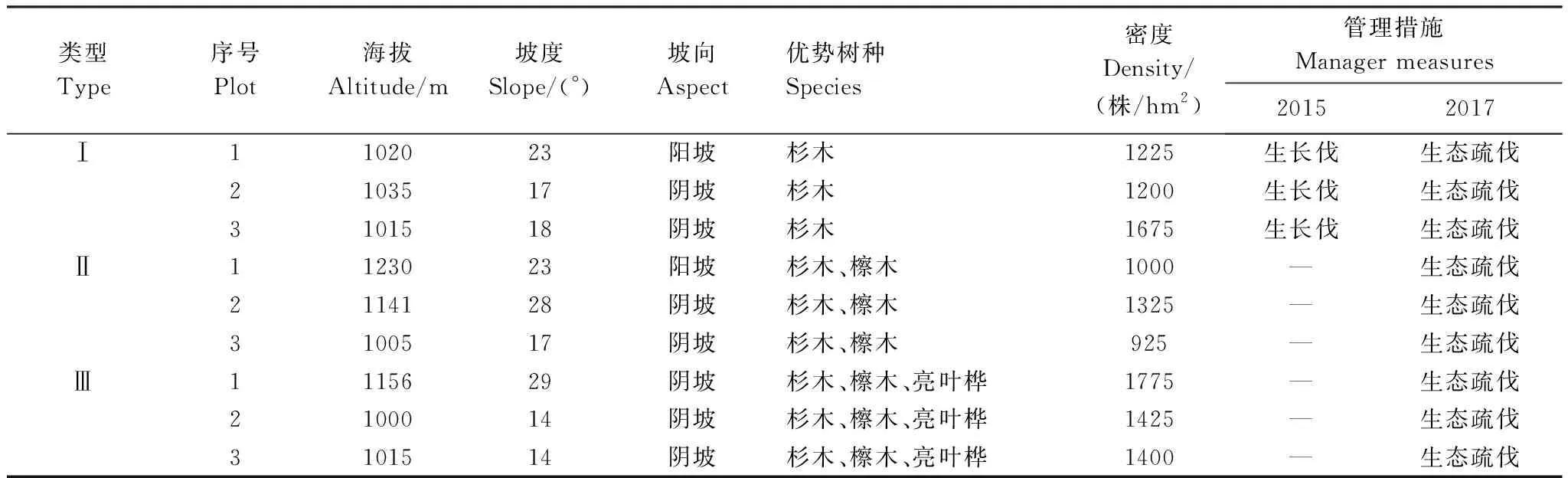
表1 样地基本信息
2.2 草本层群落特征及多样性指数
草本植物的物种拉丁学名以《Flora of China》为准,根据林下草本群落调查结果,以物种相对多度(RA)、相对高度(RH)、相对盖度(RC)计算物种的重要值pi,进而计算物种多样性指数,具体计算公式如下[25]:
pi=(RA+RH+RC)/3
J=H/lnS
式中,pi为样方内第i种物种的重要性占全部物种重要值的比值,H为Shannon-Wiener指数,D为Simpson指数(优势度指数),J为Pielou指数(均匀度指数),S为物种数量。
2.3 林地指标测量
2.3.1林分郁闭度
林分郁闭度主要采用LAI- 2200C冠层分析仪测量指标中的无截取散射(DIFN)进行换算。在各林地内均匀选取8—10个测量点,测量林地DIFN,换算得到林地郁闭度。
2.3.2混交度
混交度主要由实地测量所得。以样地内某一树种i为参照树,计算与其相邻的n株相邻木中,与参照树树种不同的树种所占的比例,主要用于描述树种的空间隔离程度。具体计算公式为:
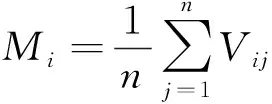
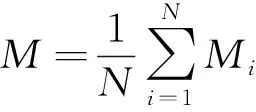

2.3.3更新潜力
通过实地踏查,统计样地内优势乔木树种(杉木、檫木、亮叶桦)的幼苗幼树数量,分析其占样地乔木数量的比例,即为林地更新潜力。
2.3.4土壤理化性质
在各样地上、中、下坡位分别选取3个样点,去除表层植被后,以环刀选取原状土壤,测量土壤容重和土壤含水量;此外,在各点位采集0—20 cm的土壤样品,混匀,测定土壤pH,土壤粒径组成、土壤有机质、全磷、全氮等理化指标。
2.4 统计分析
研究采用Excel 2019和SPSS 20.0对实验数据进行整理和分析,并在Origin 2018中完成作图。其中,对不同类型林地间林木组成、混交度、生物量、林下草本多样性等的差异性分析采用单因素方差分析(one-way ANOVA);林木组成与林下草本组成及多样性的相互关系、土壤理化性质与林下草本组成及多样性的相互关系、林下草本物种间的相互关系等则采用Person相关分析,显著性水平设定为P<0.05。
3 结果与分析
3.1 林地特征
通过对研究区杉木人工林的调查发现,区域内主要的树种配置类型有杉木纯林(I)、杉木-檫木混交林(II)以及杉木-檫木-亮叶桦混交林(III)(表2)。杉木纯林样地(I)以杉木为主,其比例达90.51%,因管理需要及人为影响,林内有少量的檫木、亮叶桦、栎属和微毛樱桃等树种,比例分别为1.37%,2.05%,3.54%和2.53%;杉木-檫木混交林样地(II),以杉木(38.33%)和檫木(34.08%)为主;杉木-檫木-亮叶桦混交林样地(III)中杉木、檫木、亮叶桦的比例分别为37.78%,17.46%和36.72%,三类样地之间优势树种具有显著差异(P<0.05)。据计算,III型样地的混交度最高(0.60),且显著高于I型(0.06)和II型(0.30)样地(P<0.05)。样地间林下幼苗的更新潜力亦表现为混交林样地(II和III)高于纯林样地,但差异不显著。此外,根据生物量模型[26]计算乔木地上生物量可知,I型样地的生物量分别比II型和III型样地高71.25%和 50.6%,而草本生物量则不同,I型、II型、III型的草本生物量分别为1.53 t/hm2、1.73 t/hm2和1.66 t/hm2。
3.2 林下草本层物种组成
研究区林下草本植物较为丰富,共有植物72种,分属于37科65属。其中,纯林样地(I)共有27科38属38种,杉木-檫木混交林(II)共有22科36属40种,杉木-檫木-亮叶桦混交林(III)25科38属40种。纯林样地的植物科、属略高于混交林样地,而物种数量则是混交林样地较高。图1是以9个标准样地林下草本层的主要物种及其重要值大小所作的热度图,由图1可知,标准样地I草本层以高粱泡、金星蕨和鱼腥草为优势种,其重要值均值分别为13.65%、8.79%和5.27%;标准样地II和III均以金星蕨、荩草和山马兰为优势种,重要值分别为10.75%、8.37%、7.47%和11.84%、6.16%、5.82%。冰草、日本蹄盖蕨Athyriumniponicum、牛筋草Eleusineindica(6.06%)、常春藤Hederanepalensis等草本植物在各样地的分布各有差异。研究发现,在本研究所涉及的72种草本植物中,有67种为多年生草本,占全部草本植物的93.06%,一年生或两年生草本仅占6.94%,其中I型样地多年生草本占89.47%,II型和III型混交林多年生草本分别为95%和90%(表3)。
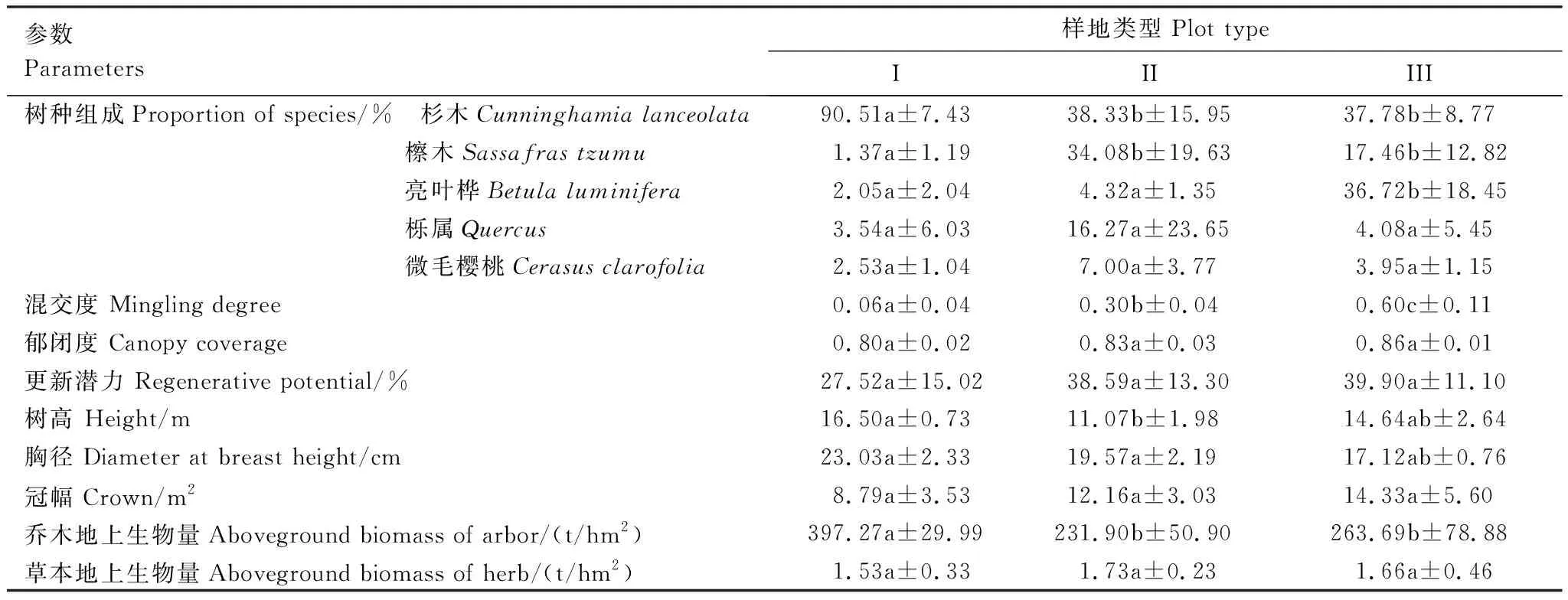
表2 林地特征

表3 杉木人工林林下草本植物名录
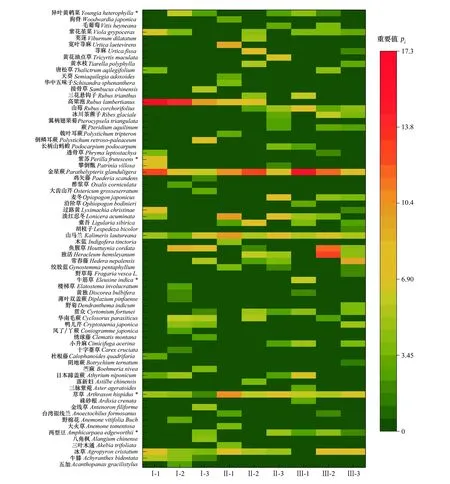
图1 样地林下草本群落重要值热度图Fig.1 Heat map of important values of understory herbaceous communities *表示一年生或二年生草本,其余为多年生草本
3.3 不同林地群落多样性分析
物种丰富度指数是样地内物种的数量,是群落物种多样性丰富程度的反应,物种数越多,物种丰富度越大。杉木纯林样地(I)的物种数均值为18.33,杉木-檫木混交林样地(II)物种丰富度均值为19,杉木-檫木-亮叶桦混交林(III)物种丰富度均值21。III型样地的物种丰富度略高于I型和II型样地,但没有显著性差异(图2)。物种Shannon-Wiener指数H、物种Pielou均匀度指数J以及物种Simpson优势度指数D均表现出相似的变化规律,林地间差异性不显著(图2)。
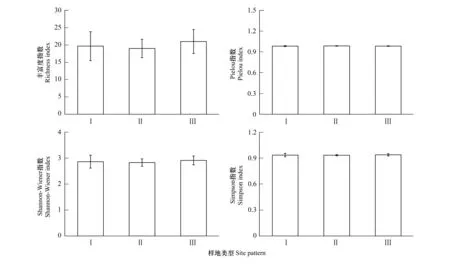
图2 各类样地林下草本层生物多样性指数Fig.2 Biodiversity index of understory herb layer in different plotsI:杉木纯林 pure forest with Cunninghamia lanceolata;II:杉木-檫木混交林 mixed forest with C. lanceolate and Sassafras tzumu plantations;III:杉木-檫木-亮叶桦混交林 mixed forest with C. lanceolate, S. tzumu and Betula luminifera plantations
3.4 林地特征与物种多样性指数及优势物种之间的关系
由以上结果可知,样地间林下草本层群落多样性差异不显著,但物种组成各有差异(图1,图2)。基于此,对样地土壤理化性质、林地树种组成与林下草本多样性、优势物种/常见种进行相关分析(表4),可知,土壤容重与草本多样性指数之间呈正相关关系,尤其与Simpson指数D之间,呈显著相关(P<0.05),而土壤含水量、土壤有机质、全氮、全磷与草本多样性指数之间呈负相关,但该相关性并不明显(P>0.05)(表4)。此外,草本层的优势物种显著影响土壤理化性质:如,金星蕨与土壤含水量之间呈显著正相关(P<0.05),高粱泡有助于增加土壤磷含量(P<0.05),而麦冬则对磷的需求较高(P<0.05)。
林地树种组成对草本多样性及优势物种亦有重要影响(表4)。对多样性指数而言,栎属树种比例的增加在一定程度上增加了草本多样性(P<0.05);林地郁闭度与林下草本物种多样性指数呈负相关。对优势物种而言,栎属树种的增加有利于常春藤的生长,但抑制了金星蕨的生长;杉木占比较高的样地高粱泡分布广,而荩草的重要值较低;檫木比例较高的样地,荩草和山马兰分布较广;亮叶桦和野樱桃比例则对各草本植物无明显影响(表4)。此外,从草本多样性与优势物种的相关性可知,金星蕨与物种多样性指数显著负相关(P<0.05),常春藤的增加则有利于增加物种多样性(P<0.05),且各物种之间存在一定的协同和竞争关系,其中,以麦冬和高粱泡之间竞争关系最为明显(P<0.05)(表5)。
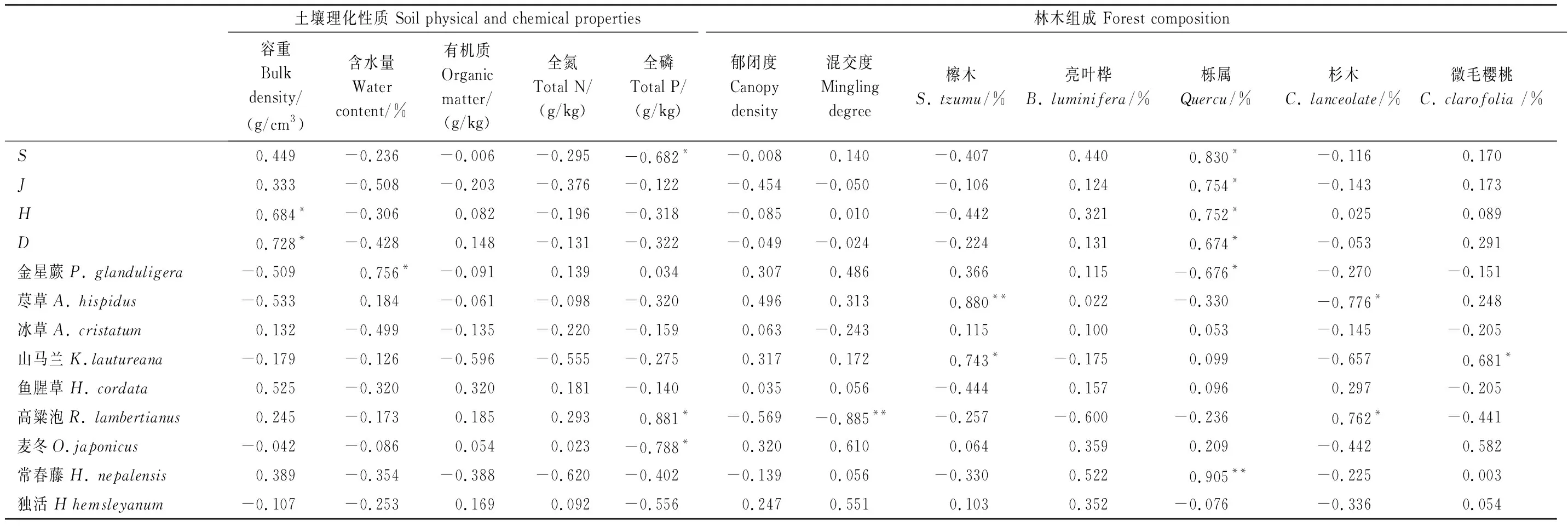
表4 林地特征与林下草本层物种多样性及优势物种的关系

表5 草本层物种多样性与优势物种间的关系
4 讨论
4.1 林分结构与生物多样性
合理的林分结构是提高森林生产力和森林生态系统服务功能的重要基础[11,27-28]。研究表明,相较于单一树种的纯林结构,针阔混交林对于改善林内微环境、提高土壤养分、增加生物多样性等方面具有重要优势[14,21]。通过阔叶化改造,林地易形成明显的复层林冠结构,提升空间资源的利用率,并且树种多样性的增加有利于增强林地抗逆性,进而提高生态系统的稳定性[14,27]。本研究中,选用了杉木纯林与杉阔混交林进行对比研究,发现混交林样地树种类型更为多样,冠层结构更为丰富,林地更新潜力大大提高(表2)。杉木是该地区主要的乔木树种,多年来被作为重要的用材林进行经营,如表1所示,为促进杉木生长,提高木材质量,缩短其成材年限,分别在2015年和2017年对杉木纯林进行了一次生长伐和生态疏伐。在该管理措施下,杉木的林分密度得到有效控制,生长条件得到改善,林木生长加快,杉木生物量迅速提高(表2)。相比而言,混交林样地仅进行了一次生态疏伐,其林木长势相对一般,且由于林地树种多样,存在一定的种间竞争,样地内林木树高、胸径均低于纯林样地,因而生物量较低。此外,研究区内纯林样地的杉木树种栽种整齐,长势均匀,但因其树种结构单一,杉木生态位占据绝对优势,其林下更新潜力略低于杉木-檫木混交林和杉木-檫木-亮叶桦混交林,仅为27.52%(表2)。针阔混交林同时具备了阔叶林和针叶林的林分特征,增加了生境的异质性,为更多林下草本的生长创造了条件[14]。檫木、亮叶桦等树种环境适应性强、生长速度快,通常被作为造林的先锋树种,且其通常在秋冬季落叶,一方面增加了样地凋落物的输入量,另一方面丰富了凋落物种类,可为林地土壤提供更为丰富的养分,进而促进林木生长。纯林样地较高的种植密度亦在一定程度上抑制了林下草本的生长。相比于混交林样地,纯林样地林下草本层的物种多样性相对较低,该结果与董卉卉等[14],郭琦等[21]的研究结果相似,而本研究中各样地之间物种多样性的差异并不显著。一方面,研究区位于中山地区,气候条件适宜,适宜各类草本植物的生长;另一方面,相较于其他地区,研究区具有更高的土壤肥力[29],给林下植被提供了良好的生存环境;再者,该地区林下草本植物多为多年生草本,在相似环境条件下,生长较为稳定,其演替过程相对缓慢[20,30];并且,在适宜的环境条件下,林下草本层宜形成特定的植物种群,进而维持稳定的生物多样性。
4.2 土壤理化性质与草本多样性
土壤是植被赖以生存和发展的基础,其物理化学性质对林木更新、林下草本生长等具有重要作用。如,田佳歆等[31]对长白山落叶松林下草本多样性的研究表明林下草本物种丰富度与土壤全磷、土壤有机质显著正相关;文海燕等[32]对科尔沁退化草地的研究亦得到相似的结果;而张荣等[33]、吕刚等[34]的研究结果正好相反,其结果表明林下草本物种丰富度与土壤全磷、土壤有机质显著负相关;此外,吕刚等[34]还指出物种优势度指数与土壤容重显著正相关,而与土壤有机质显著负相关;本研究的结果与吕刚等[34]相似,草本物种丰富度与全磷呈显著负相关,土壤容重与物种优势度指数呈显著正相关。这可能与研究区地处北亚热带中山地区,土壤磷含量相对较低有关[35],在一定程度上限制物种丰富度的增加;土壤容重的高低是土壤结构改良与否的重要指标,而较低的物种优势度则表明该生境能满足更多的物种生存。此外,对林内草本物种类型与土壤理化性质的分析又可知,金星蕨与土壤含水量显著正相关,高粱泡和全磷显著正相关,而麦冬与全磷显著负相关(表4)。金星蕨在研究区各样地内普遍存在,一方面与样地郁闭度较高(表2),适宜耐荫植物生长有关;另一方面,也在一定程度上反映了研究区土壤条件良好,适宜林下草本生长,从结果中可知,其在混交林样地中占据绝对优势,林地混交在一定程度上改变了土壤理化性质。
4.3 物种组成与群落演替
由于当前的研究缺少连续监测数据,对群落的演替进程尚不能给出确切的定论,但不同林地类型下草本植物物种组成的差异能为其提供一定参考。从以上结果可知,本研究中三种不同配置的样地具有相同的优势科,仅优先层次略有差异:如,杉木纯林样地中林下草本分属于27科38属38种,依次以蔷薇科、金星蕨科、禾本科和菊科为优势物种;杉木-檫木混交林22科36属40种,优势物种分属于禾本科、金星蕨科、蔷薇科和菊科;杉木-檫木-亮叶桦混交林25科38属40种,优势物种依次为禾本科、金星蕨科、菊科和蔷薇科。该研究中,蔷薇科植物多为低矮小灌木,如高粱泡、山莓、三花悬钩子等,因其长势矮小,因而归为草本植物进行分析。杉木纯林样地因其林地树种单一,与混交林相比冠层盖度和郁闭度较低,林内光照相对较为充足,适宜林下灌木的萌发和生长,因而蔷薇科植物相对较为丰富;然而,在研究区内,各样地的郁闭度均大于0.7(表2),林下接受光照的面积有限,在一定程度上限制了灌木层的生长,但同时也促进了金星蕨等耐荫物种的生长[20]。而禾本科和菊科成为主要优势科,可能是因为禾本科草本植物起源于热带森林或林缘开放的生境,对环境适应性强,且其颖果产生的特殊附属物有助于实现自身远距离传播;菊科植物则是利用自身结实量大、且种子小、密被冠毛、刺毛等附属物的优势实现远距离传播,进而扩大分布范围的目的[36]。由于区域内的地理环境较为相似,草本植物的种属差异本身并不明显,而在林地树种组成差异以及人为经营管理的影响下,优势物种的组成比例及分布随其所处的生境会有所改变。此外,已有研究表明,郁闭度较大的林分,林下草本层生态位重叠度较小,种间竞争强度小,在群落演替进程中,群落会逐渐处于相对稳定状态[20]。而且,研究发现,杉木-檫木混交林和杉木-檫木-亮叶桦混交林样地中多年生草本比例分别比纯林样地增加了6.18%和0.59%,混交林的群落相对较为稳定,这与之间的研究结果相一致,中等混交度的杉木林地(II)具有更高的群落稳定性[35]。
5 结论
本研究以三种不同类型的杉木人工林为研究对象,通过分析,发现混交林样地相比于杉木纯林样地,其树种类型更为多样,冠层结构更为丰富,林地更新潜力更高,同时亦具有更为丰富的林下草本植物。研究区地理环境优越,适宜林下植被生长,各类型之间林下草本多样性指数的差异并不明显,林地经过多年的生长,逐渐达到郁闭,林下草本群落在自然演替的进程中逐渐形成了特定的植物种群:杉木纯林样地以高粱泡、金星蕨和鱼腥草为优势种,杉木-檫木混交林样地以及杉木-檫木-亮叶桦混交林样地则以金星蕨、荩草和山马兰为优势种。林地草本以多年生草本植物为主,植物群落相对较为稳定。相比于高混交度的杉木-檫木-亮叶桦混交林样地,中度混交的杉木-檫木样地多年生草本占比更高,且其物种多样性适中。因而,从群落稳定性及可持续发展角度,对于杉木纯林的阔叶化改造,适宜的中度混交模式或许更为有利。
猜你喜欢
青年文学家(2021年16期)2021-08-05
今日中国·西班牙文版(2021年4期)2021-04-09
少儿美术(2020年8期)2020-11-06
共产党员·下(2018年6期)2018-12-05
现代园艺(2018年2期)2018-03-15
方圆(2018年23期)2018-01-07
中国民族医药杂志(2016年1期)2016-05-09
小说月刊(2014年1期)2014-04-23
