
腾格里沙漠东南缘固沙植被区生物土壤结皮及下层土壤有机碳矿化特征
2021-04-25 08:24李云飞李小军
生态学报 2021年6期
谢 婷,李云飞,李小军,*
1 中国科学院西北生态环境资源研究院沙坡头沙漠研究试验站, 兰州 730000 2 中国科学院大学, 北京 101408
植被恢复与重建是干旱沙区退化土地修复的有效途径,是该区生物土壤结皮(Biological Soil Crusts,BSCs)发生与发展的关键影响因素[1- 2]。固沙植被建立后,沙丘表面逐渐趋于稳定,以蓝藻为优势的生物土壤结皮开始拓殖,尔后随着植被恢复过程中生物和非生物环境的进一步改善,藻类结皮逐渐向着以荒漠藻类、地衣及藓类等为优势的生物结皮类型演变[3]。BSCs占干旱区地表活体覆盖的40%—70%,是该区地表景观的重要组成部分和土壤碳循环的主要参与者[4- 6]。已有研究表明,BSCs的年固碳量可达11.36—26.75g C/m2[7],而结皮呼吸释放的碳占其固碳量61%[8]。在半干旱草地生态系统中,BSCs呼吸可达土壤呼吸总碳释放量的43%,远大于植被和裸地斑块土壤呼吸的贡献[9]。因此,BSCs的拓殖和发育在干旱区碳循环过程中发挥重要作用。
土壤有机碳矿化是陆地生态系统到大气碳通量最大的组成部分,对维持土壤肥力和养分释放、缓解温室气体的增加及全球气候变化的影响至关重要[10- 11]。沙区固沙植被建立后,伴随着植被的演变,BSCs的物种组成、盖度、生物量、理化性质、微生物属性及水热因子等均发生了显著的变化[7,9,12- 15]。BSCs的这种变化使其本身参与的生态水文过程发生显著变化,进而可能影响其碳循环过程[5- 7,9,12]。因此,BSCs演变对碳矿化的影响逐渐受到关注。如杨雪芹[16]等研究发现,BSCs在由藻类为优势到经藻类和藓类混生向以藓类为优势的结皮演变过程中,其碳矿化速率由0.03 g kg-1d-1增加到0.04 g kg-1d-1,累积可矿化碳量的比例在4.7%—6.9%之间。李云飞等[17]通过对不同发育阶段的BSCs研究发现,有机碳的瞬时及最大矿化速率均表现为藓类结皮>地衣结皮>藻类结皮土壤。然而,已有研究主要侧重于BSCs自然演替过程中碳矿化的变化,而对固沙植被演替引发的BSCs生物和非生物过程演变对土壤碳矿化的研究较缺乏,这使得我们对沙区人工植被建设对土壤碳库动态变化及土壤质量的影响机理认识不足,在一定程度上制约对固沙植被的生态学效应的准确评价。
拟通过对腾格里沙漠东南缘不同恢复年限固沙植被区BSCs及其下层土壤碳矿化特征,及其与水分和土壤理化性质之间关系的研究,揭示固沙植被恢复过程中BSCs的拓殖和发育对土壤碳矿化过程的影响,阐明BSCs对荒漠生态系统碳循环的贡献及关键影响因素,为全面认识干旱区BSCs的生态功能提供科学依据。
1 材料与方法
1.1 研究区概况
实验区位于腾格里沙漠东南缘,中国科学院沙坡头沙漠研究试验站包兰铁路以北的人工固沙植被区(37°32′N,105°02′E)。该区为荒漠化草原向草原化荒漠的过渡带,也是沙漠和绿洲的过渡带。平均海拔1339 m,年均气温为9.6℃,最低气温-24.5℃,最高气温38.1℃;年均降水量约为186 mm,且80%的降水集中在5—9月;年潜在蒸发量为3000 mm,年平均风速为2.8 m/s,最大风速19 m/s,大于5 m/s的起沙风每年有200 d左右。
为了确保包兰铁路沙坡头沙漠地段的畅通无阻,科研人员于1956年开始在流动沙丘上设置1 m×1 m的麦草方格沙障并种植油蒿(Artemisiaordosica)、柠条锦鸡儿(Caraganakorshinskii)和花棒(Hedysarumscoparium)等旱生灌木,尔后在不同年份按照相同方法进行扩展,在包兰铁路两侧形成了南北宽约700 m,长16 km非灌溉人工固沙植被防护林体系[2,18]。该区的主要草本植物有:小画眉草(Eragrostisminor)、雾冰藜(Bassiadasyphylla)、刺沙蓬(Salsolaruthenica)、狗尾草(Setariaviridis)和茵陈蒿(Artemisiacapillaris)。人工固沙植被区建立后经过60多年的演变,植被—土壤系统发生了深刻的演变,BSCs也逐渐形成并发育,该区BSCs的主要优势生物成分为藻类、地衣和藓类[19]。不同恢复年限固沙植被区结皮层和下层土壤的理化性质见表1。
1.2 实验设计
2019年5月,分别在1956年(63a,恢复年限)、1973年(46a,恢复年限)、1987年(32 a,恢复年限)和2011年(8 a,恢复年限)建立的人工固沙植被区和流沙区(0 a,恢复年限)设置10 m×10 m的样方各3个。固沙植被区每个样方中随机选取10个采样点,分别用直径为5 cm的土钻采集结皮层及结皮下0—5 cm层土壤样品;流动沙丘采用同样的方法采集表层0—2 cm土壤和下层2—7 cm土壤。同一样方中同一土层不同采样点样品混合成一个样品带回实验室,共30个混合样。采集的土样在室内自然风干,去除残留的枯枝落叶后过2 mm筛。将风干土分为两份,一份用于测定土壤理化性质,另一份置于4℃冰箱,用于有机碳矿化的测定。
1.3 测定指标及方法
土壤pH值用pH计法(水∶土=5∶1)测定,土壤电导率采用电导仪(Cole-Parmer Instrument Company, Illinois, USA)测定,土壤颗粒组成采用采用激光粒度仪(Microtrac S3500, Microtrac, Montgom-eryville, USA)测定,利用压力膜仪(5bar压力膜仪,1600和15bar压力膜仪,1500F1,USA)测定水分特征曲线,并计算田间持水量。土壤全氮用FOSS凯氏定氮仪测定,土壤有机碳采用重铬酸钾氧化-外加热法测定[20]。
土壤有机碳矿化采用室内恒温培养-碱液吸收法测定。取出4℃条件下保存的样品,分别称取50 g置于250 mL的可密封广口瓶中,把广口瓶中土壤的水分用蒸馏水分别调节至土壤含水量的5%、10%和20%,再放入20℃的恒温培养箱中进行5天的预培养,预培养完成后即正式开始培养实验。分别在正式培养的第2、4、7、12、17、27、37、47天和第57天将盛有10 mL 0.2 mol/L NaOH溶液的25 mL的小烧杯置于广口瓶中,用于吸收有机碳矿化释放的CO2,广口瓶加盖密封24小时后取出装有NaOH溶液的小烧杯,用浓度为0.2 mol/L的HCl溶液进行滴定,计算培养过程中土壤碳矿化的累积释放量(mg C/kg)和矿化速率(mg C kg-1d-1)。
1.4 数据处理
采用重复测量方差分析法分析土层、恢复年限和水分对土壤有机碳矿化瞬时速率和累积释放量的影响,采用多因素方差分析法分析土层、恢复年限和水分对最大矿化速率和平均矿化速率的影响,利用最小显著差异法(LSD)进行显著性检验,显著性水平为0.05。土壤有机碳矿化最大速率、平均速率及累积释放量和土壤理化性质及土壤水分含量之间的相关性的冗余分析用Canoco 5.0软件实现。采用 SPSS 17.0进行数据统计分析。用 Origin 9.0拟合曲线并作图。
2 结果与分析
2.1 土壤有机碳矿化速率
整个培养期间,土壤有机碳矿化速率为培养前期快速下降,后期逐渐下降并趋于平缓。且同一土壤水分含量下,不同恢复年限固沙植被区碳矿化速率表现为63 a>46 a>32 a>8 a>0 a,同一植被区表现为BSCs >下层0—5 cm土壤(图1)。土壤水分含量的增加显著促进了土壤有机碳矿化的速率(表2,P<0.001),土壤水分含量从5%增加到20%时,不同恢复年限BSCs有机碳化的平均速率及最大速率分别增加了1.48—2.08倍和1.60—2.00倍,下层0—5 cm土壤增加了1.36—2.08倍和1.21—2.00倍(图1,图2)。
不同恢复年限、水分和土壤层次对土壤有机碳矿化的瞬时速率、最大速率影响显著(表2,P<0.001);且恢复年限、水分与土层的交互作用的影响也显著(表2,P<0.001)。
2.2 土壤有机碳矿化累积释放量
整个培养期间,不同恢复年限固沙植被区土壤有机碳矿化累积释放量为随培养时间的延长呈不断增加的趋势。同一土壤水分含量下,不同恢复年限固沙植被区表现为63 a>46 a>32 a>8 a>0 a,同一植被区表现为BSCs >下层0—5 cm土壤(图3)。土壤水分含量的增加显著促进了土壤累积释放量(表2,P<0.001),土壤水分含量从5%增加到20%时,BSCs累积碳矿化量增加了1.48—2.08倍;下层0—5 cm土壤累积释放量增加1.21—2.00倍(图3)。

表1 不同恢复年限固沙植被区BSCs及下层0—5 cm土壤理化性质
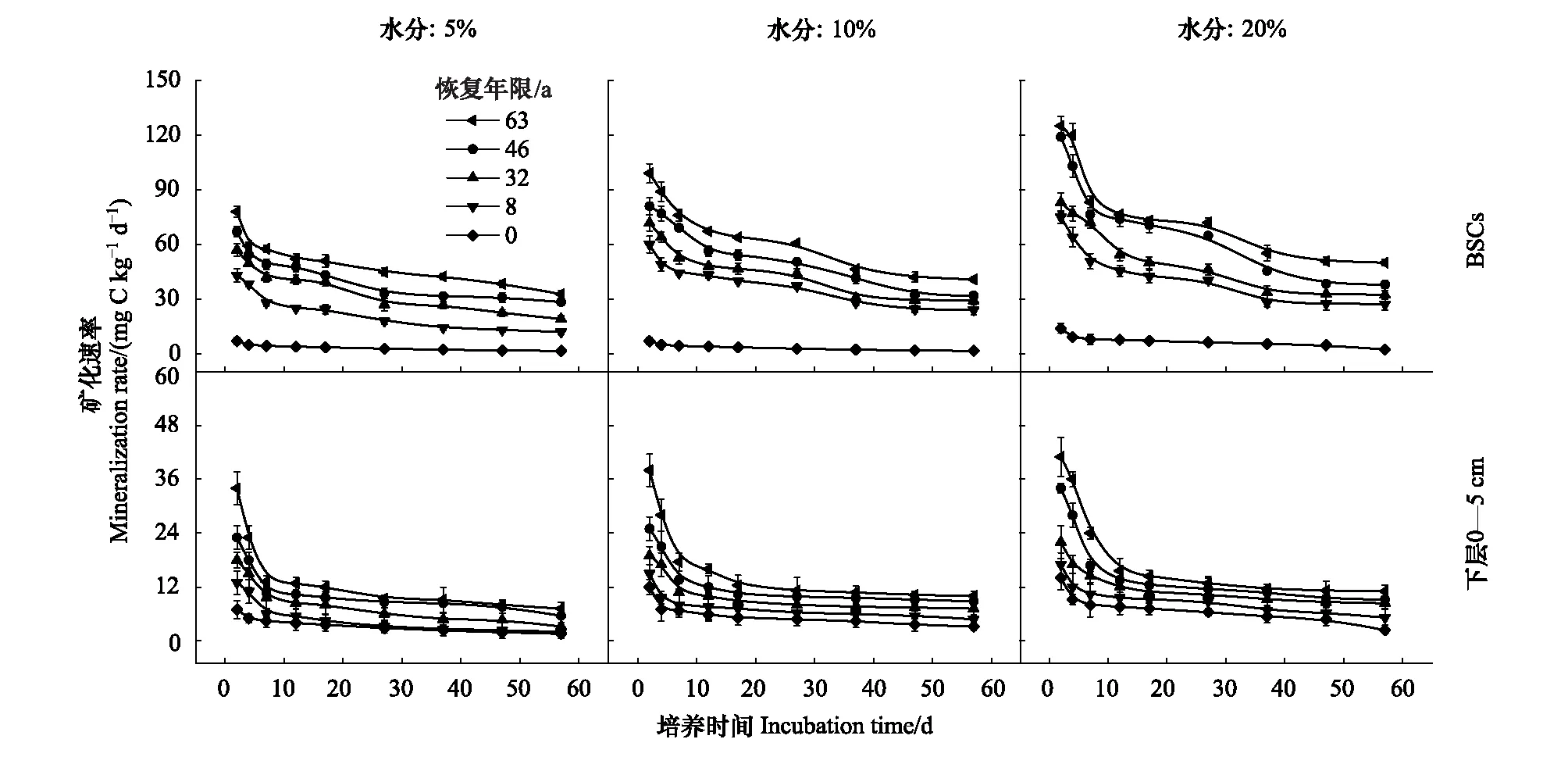
图1 不同培养时间和水分条件下生物土壤结皮和下层0—5 cm土壤有机碳矿化速率Fig.1 Soil organic carbon mineralization rate in the biological soil crusts (BSCs) and the 0—5 cm subsoil under different incubation time and water conditions
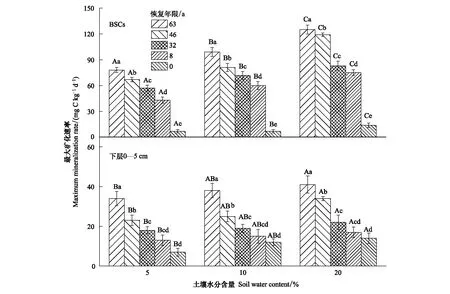
图2 不同土壤水分含量下BSCs和下层0—5 cm土壤有机碳矿化最大速率Fig.2 The maximum rate of soil organic carbon mineralization in the BSCs and the 0—5 cm subsoil under different soil water content不同大写字母表示不同土壤水分含量间差异显著(P<0.05),不同小写字母表示不同恢复年限间差异显著(P<0.05)
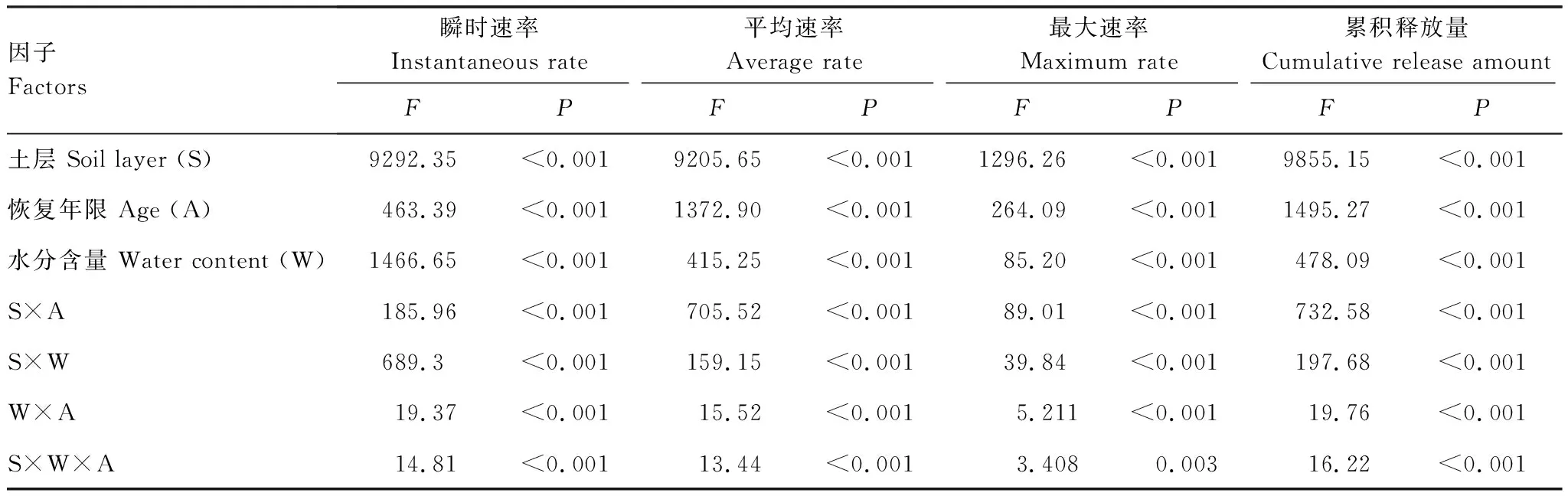
表2 土层、恢复年限和水分含量对土壤有机碳矿化瞬时速率、平均速率、最大速率和累积释放量的影响

图3 不同培养时间和水分条件下BSCs和下层0—5 cm土壤有机碳矿化累积释放量Fig.3 The cumulative release amount of soil organic carbon mineralization in the BSCS and the 0—5 cm subsoil under different incubation time and water conditions
2.3 BSCs和下层0—5 cm土壤有机碳矿化之间的关系
BSCs有机碳矿化的瞬时速率、平均速率、最大速率、累积释放量与下层0—5 cm土壤瞬时速率、平均速率、最大速率、累积释放量呈显著的正相关关系(图4,P<0.001)。

图4 BSCs和下层0—5 cm土壤在有机碳矿化的瞬时速率、平均速率、最大速率、累积释放量的关系Fig.4 Relationship between the instantaneous rate, average rate, maximum rate, and cumulative release amount of organic carbon mineralization in the BSCs and the 0—5 cm soil under crust
2.4 土壤有机碳矿化与土壤理化性质间的关系
冗余分析结果表明,有机碳矿化与土壤水分含量及土壤理化性质密切相关,不同因子对有机碳矿化的解释程度有着较大的差异,结皮层第一轴和第二轴分别解释了有机碳矿化变异的94.05%和1.56%,下层0—5 cm土壤分别解释了79.27%和3.76%(图5)。其中,结皮厚度、生物量、电导率、有机碳、全氮、粉粒、粘粒、全氮、田间持水量和土壤水分含量与有机碳矿化的最大速率、平均速率和累积释放量显著正相关(图5,P<0.01),而与pH和沙粒含量显著负相关。结皮层中生物量、电导率、厚度、黏粒含量和有机碳是影响有机碳矿化的主要因素,下层0—5 cm土壤中有机碳、电导率和黏粒含量是影响有机碳矿化的主要因素。土壤电导率、有机碳和黏粒含量是影响有机碳矿化的主要影响因素(图5)。

图5 BSCs和下层0—5 cm土壤有机碳矿化与土壤理化性质之间的相关性Fig.5 Correlation between soil organic carbon mineralization and soil physicochemical properties in the BSCs and the 0—5 cm subsoil
3 讨论
不同恢复年限固沙植被区BSCs和下层0—5 cm土壤碳矿化速率为培养前期快速下降,后期逐渐下降并趋于平缓(图1),表明微生物作用下土壤的有机碳矿化过程很大程度上取决于土壤中养分的供应[21- 22]。培养前期,土壤中大量可被微生物分解利用活性有机物质等逐渐消耗,随着培养时间延长,微生物开始利用较难分解的复杂有机物,但这种过程缓慢[22- 23],因此土壤有机碳矿化速率逐渐减小后趋于稳定。这种有碳矿化速率先快后慢的变化趋势在很多研究中均有报道[17,21- 23]。
同一土壤水分含量下,BSCs有机碳矿化的瞬时速率、平均速率、最大速率和累积释放量均显著大于下层0—5 cm土壤(表2,P<0.001),不同恢复年限固沙植被区土壤机碳矿化的瞬时速率、平均速率、最大速率和累积释放量均表现为63a>46a>32a>8a>0a(图1、图2和图3)。这与周玉燕等[23]和Su等[24]在旱地的研究结果相类似。说明随着恢复年限的延长,土壤的结构,所含养分含量等因素发生了变化[23]。有机碳矿化过程受植被类型及凋落物、根系分泌物的性质和数量、参与碳矿化过程微生物数量和活性的直接影响,同时也受土壤碳库的丰缺状况、pH值等的间接影响[25]。不同恢复年限下地表植被类型、盖度及多样性及土壤理化性质发生变化,从而使土壤有机质、土壤氮含量及微生物群落的结构及活性存在差异,进而影响土壤有机碳矿化速率。此外,本研究冗余分析表明土壤有机碳矿化的最大速率、平均速率和累积释放量与土壤的理化性质显著相关(图5,P<0.01),其中电导率、有机碳和黏粒含量是影响有机碳矿化的主要影响因素。
本研究认为植被恢复过程BSCs的演变也是其碳矿化差异原因主要表现在以下几个方面:(1)随着BSCs的演变,其盖度和粗糙度逐渐增大,捕获和富集大气降尘的能力逐渐增强,从而使结皮的厚度、土壤细颗粒物质和养分含量逐渐增大[26](表1),此外,BSCs还能够改善微生境土壤的水热因子,使得结皮层的代谢活性增强[9,12]。(2)随着BSCs的演变,其盖度和生物量显著增加[7,9,12](表1),从而使结皮层单位面积CO2交换速率显著提高[5,7,9,12]。(3)随着BSCs的演变,BSCs的类型(藻类、苔藓、地衣)的构成比例发生变化,从而影响碳矿化速率。通常与地衣和苔藓相比,藻类结皮具有较低的生物量和叶绿素含量[5,7,12],以及更加有限光穿透力[27],因此使得固碳能力相对降低[7,9,12]。比如,干旱区以具鞘微鞘藻为主的生物结皮的固碳速率约(1 μmol CO2m-2s-1)显著低于地衣和苔藓为主的BSCs的光合速率(10 μmol CO2m-2s-1)[28-29]。(4)BSCs的不同演替阶段具有不同的微生物组成和群落结构[30],对其碳矿化影响也不同。已有的研究报道了相似的研究结果[30- 31]。此外,本研究冗余分析表明结皮层生物量、电导率、厚度、黏粒和有机碳含量是影响有机碳矿化的主要因素(图5)。
结皮层和下层0—5 cm土壤的各种有机碳矿化特征呈显著的正相关关系(图4,P<0.001),说明下层土壤各种碳矿化特征随着BSCs各种碳矿化特征的增大而增大。BSCs和下层0—5 cm土壤有机碳矿化速率和累积矿化量变化趋势基本一致(图1、图3),但BSCs的碳矿化速率和累积矿化量均显著高于下层0—5 cm土壤(表2,P<0.001),其他类似研究中也发现了相似的规律[17]。究其原因主要有以下三点。首先,同一植被区,隐花植物的凋落物、分泌物、残体以及大气降尘等不断在土壤表层积累和分解,有效促进了生物土壤结皮有机质、养分含量的增加,使得结皮层及下层土壤有机碳含量存在一定的差异(表1),这也是生物土壤结皮层及下层0—5 cm土壤碳矿化差异的主要原因。此外,不同土层的微生物数量和活性是影响碳矿化差异的另一个原因。BSCs微生物量熵大于下层土壤[32],这显然为土壤碳矿化提供了微生物基础;另一方面,BSCs在获得充足的养分的同时,首先显著改善BSCs土壤的理化性质,这为微生物的生存提供了适宜的条件,而下层土壤,除了BSCs所提供的养分很难到达外,理化性质变差也可能是一个重要的因素。本研究(表1和图5)也证实了这一点。最后,BSCs是干旱沙区土壤碳和氮输入的重要来源,这有利于结皮层养分的聚集。研究发现,随着BSCs的发育,BSCs有机碳含量增加较为突出,10年以上的藓类结皮土壤有机碳含量可高达20.9 g/kg[33- 34],BSCs通过固定大气中的氮进而增加土壤氮输入量,达2—15 kg N hm-2a-1[35],这均为BSCs的碳矿化提供了物质基础,使得BSCs土壤和下层土壤的碳矿化过程存在一定的差异。
BSCs和下层0—5 cm土壤有机碳矿化瞬时速率、平均速率、最大速率和累积碳释放量均随土壤含水量的增大而增大(图1,图2,图3),其他旱地研究中发现了相似的规律[12,24]。表明土壤碳矿化过程主要与微生物的调节有关[36]。土壤含水量增加会使微生物的活性显著提高,进而加速了有机质的分解[37],另一方面,高的土壤水分能改变土壤养分的扩散速率,通过释放更多的土壤活性来提高土壤微生物基质的利用率[38]。此外,水分对不稳定有机碳组分含量和顽固性有机碳化率产生影响[39],土壤质地也可能影响微生物群落及细菌、真菌对土壤呼吸的贡献[40]。表明水分对碳矿化的调控作用不仅受到微生物活性的影响,也与土壤质地、有机碳组分等密切相关。因此,水分对有机碳矿化的作用机理是一个值得进一步探讨的问题。
4 结论
不同恢复年限固沙植被区土壤机碳矿化的瞬时速率、平均速率、最大速率和累积释放量均表现为63a>46a>32a>8a>0a,且同一植被区表现为BSCs >下层0—5 cm土壤,且其碳矿化过程主要受电导率、有机碳和黏粒含量的影响。土壤水分含量从5%增加到20%时,BSCs土壤有机碳矿化平均和最大速率及累计释放量分别增加了1.48—2.08倍、1.60—2.00倍和1.48—2.08倍,下层土壤分别增大了1.36—2.08倍、1.21—2.00倍和1.36—2.08倍,表明水分促进了固沙植被恢复过程中土壤有机碳矿化过程。由于本研究着重探讨的是不同恢复年限固沙植被区BSCs及下层土壤在不同水分条件下的碳矿化特征,为了更深入了解植被恢复过程中BSCs演变对土壤有机碳矿化过程影响,还需要BSCs的盖度、生物量及微生物属性等进一步研究。
猜你喜欢
农业工程学报(2022年8期)2022-08-08
北京大学学报(自然科学版)(2022年3期)2022-06-17
小猕猴学习画刊(2022年3期)2022-03-28
科技视界(2020年19期)2020-07-30
读者·校园版(2019年3期)2019-01-28
铁道标准设计(2018年11期)2018-10-20
少年文艺·开心阅读作文(2017年1期)2017-02-24
中国农业文摘-农业工程(2016年5期)2016-04-12
环球市场信息导报(2015年22期)2015-12-02
小天使·四年级语数英综合(2014年3期)2014-03-21
