
高寒草甸茎直黄芪物候对模拟季节性不对称增温的响应
2021-05-07 07:24尕藏加斯确多吉索南吉边巴普赤谢正新
草地学报 2021年4期
尕藏加, 斯确多吉, 索南吉, 边巴普赤, 南 吉, 谢正新
(1. 西藏大学理学院生物系, 西藏 拉萨 850000; 2. 中国科学院青藏高原研究所高寒生态重点实验室, 西藏 拉萨 850000;3. 中国科学院青藏高原地球科学卓越创新中心, 北京 100101; 4. 西藏大学-中国科学院青藏高原研究所那曲生态环境综合观测研究站, 西藏 那曲 852000; 5. 青海师范大学生命科学学院, 青海 西宁 810016; 6. 高原科学与可持续发展研究院, 青海 西宁 810016; 7. 中国科学院大学, 北京 100049)
植物物候是植物对季节性环境变化产生的节律性生理和生长响应特征,对其生存的环境变化特征十分敏感[1]。植物物候对环境变化的响应特征不仅可以用来指示植物对环境变化的适应性[2],而且植物物候变化可直接影响植物生长和繁殖[3],最终影响群落结构[4]以及生态系统功能[5]。
在过去的50年内,青藏高原气温每10年上升0.4℃左右,远远高于全球平均水平[6-7],导致青藏高原草地植被/植物物候发生变化[8]。大多数长期观测研究[9]和模拟增温控制试验研究表明[10-11]近30年来青藏高原的植物物候初始期的变化总体表现为返青和初花期提前、枯黄期延迟[9-12],而结实期则保持相对稳定[11,13],生长期长度延长[14]。但由于研究方法和地点以及研究对象等差异,这些植物物候变化的驱动因子还存在较大的不确定性,难以得出统一的物候研究结论。
现有大多数野外控制试验研究[8]是在仅生长季增温或全年恒定增温的条件下探讨气候变化对植物物候的响应特征[15],但气候变化呈现季节性不对称性,即冬季增温幅度高于生长季增温[16]。当冬季增温幅度高于生长季增温时,可能导致植物春化阶段受阻,植物返青物候延迟[3,17-19]。另外,模拟增温可导致土壤湿度降低,但关于温度和水分之间的交互影响研究鲜见。
茎直黄芪(AstragalusstrictusGrah. Ex Bunge)是青藏高原腹地纳木错区域典型的高寒草甸的常见的豆科黄芪属多年生草本植物,具备固氮能力,在草地生态系统氮循环过程中发挥重要作用[20]。茎直黄芪根系相对较深,具有较强的抗寒、耐旱、喜光等性状,对气候变暖相对敏感。故本研究以茎直黄芪为研究对象,利用开顶式模拟增温装置(Open top chambers,OTCs),在青藏高原腹地纳木错区域开展了季节性不对称增温野外控制试验,以期探讨季节性不对称气候变化特征对高寒植物物候的影响特征及其过程。
1 材料与方法
1.1 研究区概况
研究区位于西藏中部念青唐古拉山南麓纳木错湖附近典型高寒草甸草地发育区(30° 41.208′~30° 44.579′ N;91° 01.117′~91° 04.590′ E),海拔高度约4 875 m[18]。试验区的年平均气温在0℃左右,7月是气温最高的月份,12月是气温最低的月份,全年大约有4个月温度在0℃以上。研究区受季风、西风及其交互作用的影响[21-23],空气和土壤湿度在时间上分布不均匀,空气湿度和土壤湿度在1月最低,8月最高,年平均空气湿度约为52%。该区域年降水量约为407 mm[4],主要集中在6月到9月[18]。雨季通常在6月底或7月初开始,是该地区降水的主要来源。强降雪是该区的冬季主要降水来源。该区域主要优势植物为高山嵩草(KobresiapygmaeaC. B. Clarke),主要伴生种有紫花针茅(StipapurpureaGriseb)、矮嵩草(KobresiahumilisL)、茎直黄芪、雪白委陵菜(PotentillasaundersianaRoyle)等。
1.2 试验设计
本试验设全年增温(ANW)、仅生长季增温(GRW)和对照(Control,未经任何处理)共3个处理,生长季增温的增温罩仅在植物生长开始到枯黄期阶段(即4月末至10月末)放置,全年增温的增温罩则常年保留在对应处理样圆上。增温处理各重复4次,对照则重复了8次(便于选择,且提高数据质量),共计试验处理小区16个,每个小区直径为8 m。因在全年增温条件下冬季增温幅度高于夏季增温(OTCs自然增温效应所致),而在仅生长季增温条件下冬季不增温,所以实现了季节性不对称效应。生长季增温处理始于2008年,于2015年起又增设了全年增温处理,以探讨不同季节增温对高寒草甸草地生态系统的影响。为方便在同一处理下(每个处理小区内)开展不同目的的科学观测、样品采集等工作,以避免因样品采集等活动干扰其他长期观测内容(包括植物群落、物候、土壤温湿度等),同时为了有效实施OTCs增温处理,在每个处理小区内(直径8 m)又随机设立了5个(直径为1.5 m)处理小样圆。本研究中的植物物候观测是在每个处理小区内的其中一个长期不受任何取样干扰的指定样圆内进行开展。增温罩底部直径为1.5 m,顶部直径为0.75 m,高度为0.4 m(图1B)。
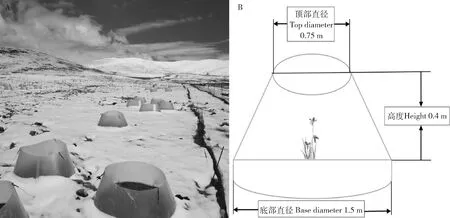
图1 纳木错样地OTCs模拟增温实验示意图
1.3 数据采集与整理
1.3.1温湿度数据 在每个处理小区内其中一个小样圆的正中央配置Decagon ECT,5TM和EC-TM传感器和EM50数据采集器(Decagon Devices,Pullman,Washington,USA)测量不同处理下的距离地面10 cm处的空气温度和地下10 cm处的土壤温度和土壤水分。所有传感器每15 min记录一次空气温度、土壤温度和土壤湿度数据。利用采集的数据整理生长季阶段(4月1日至10月30日),非生长季阶段(11月1日至第二年3月30日),白天(早晨8∶00至夜晚7∶45分钟),夜间(晚8∶00至第二天早晨7∶45分钟),午间(14∶00至15∶45),年均(全年)空气温度、土壤温度和土壤湿度数据。
1.3.2物候观测数据 在每个处理小区内的其中一个长期不受任何取样干扰的指定样圆正中央选择一个0.75 m×0.75 m观测样方,用铁钉固定其4角作为植物物候观测点。在每个固定样方内选择20株茎直黄芪个体(若部分样方内不足20株时,选择所有个体),每年观测返青物候,仅2019年增加了枯黄物候观测。利用采集的数据(国际儒略历Julian day)确定和计算返青期(每个处理样方内50%个体呈现返青的日期),枯黄期(每个处理样方内50%个体呈现枯黄的日期),生长期长度(每个样方内枯黄期减去返青期所得数值)。因研究区域地理特殊,交通不便和后勤等问题,仅2019年获取了枯黄物候以及生长期长度数据。
1.4 数据分析
我们使用广义线性混合效应模型(GLME)和方差分析(ANOVA)方法对数据进行分析[24]。在方差分析中,处理(Control,ANW,GRW),年份(2016—2019)及其相互作用(处理×年份)设定为自变量,茎直黄芪的返青期、枯黄期和生长期长度设定为应变量,分析不同增温处理以及年际差异对茎直黄芪物候的影响。同时我们利用GLME分析茎直黄芪返青、始黄期和生长期长度与生长季、非生长季、白天、夜间、午间、年均空气温度,土壤温度和土壤湿度之间的相关关系,设定变量“年”作为随机变量。所有分析均利用IBM SPSS 25.0版本和R 4.0.2版本进行。统计检验结果P<0.05和P<0.01分别表示显著和极显著。相关统计绘图由GraphPad Prism 8.0版本完成。
2 结果与分析
2.1 增温对空气温度、土壤温度和土壤湿度的影响
2.1.1增温对空气温度、土壤温度和土壤湿度的影响 2016—2019年在全年增温条件下,与对照比较,空气温度分别增加0.6℃,0.2℃,0.3℃,0.2℃ (图2A);土壤温度则分别增加2.5℃,1.9℃,1.6℃,0.9℃ (图2B);年均土壤湿度分别降低11%,4%,8%,2%(图2C)。
2016—2019年在生长季增温条件下,与对照比较,空气温度分别增加0.4℃,0.1℃,0.2℃,0.2℃ (图2A);土壤温度分别增加了1.7℃,1.7℃,0.8℃,0.6℃(图2B);年均土壤湿度分别降低3%,3%,5%,2%(图2C)。
2.1.2年际间微气候条件的变化特征 微气候条件在年际间差异显著。2016—2019年对照样方空气温度、土壤温度和土壤湿度结果表明:2017年空气温度和土壤温度最高,分别为0.24℃和2.58℃;2018年的土壤湿度最高,为20%(图2C)。
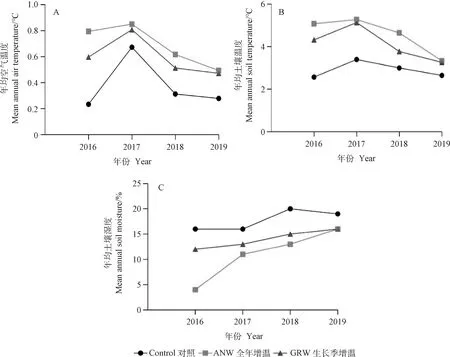
图2 对照、全年增温和生长季增温处理下年均空气温度、土壤温度和土壤湿度在年际间的变化(2016年-2019年)
2.2 返青期、枯黄期和生长期长度的变化特征
2.2.1返青物候 茎直黄芪的平均返青物候在年际间差异显著(表1,图3A)。与2017年基准年比较,2019年在所有处理条件下均显著提前了返青物候,与2019年比较,2017年和2018年的返青物候均显著延迟(图3A)。
2016年,与对照相比,全年增温处理提前了茎直黄芪的返青物候,提前幅度为5.7 d,生长季增温处理延迟了返青物候,延迟幅度为4.6 d。2017年,全年增温和生长季增温处理与对照比较均延迟了返青物候,延迟幅度分别为1.6 d,2.9 d。2018年与2016年相反,与对照相比,全年增温处理延迟了返青物候,延迟幅度为5.7 d,而生长季增温处理提前了返青物候,提前幅度为13 d。2019年在全年和生长季增温条件下与2017年的情况较接近,与对照相比均延迟了茎直黄芪的返青物候,延迟幅度为9.5 d和16.5 d(图3B)。
2.2.2枯黄物候和生长期长度 2019年的茎直黄芪在全年和生长季增温条件下的枯黄起始期分别为10月27日(DOY:300)和10月21日(DOY:294)(图4A),与对照比较分别推迟了7.7 d和1.9 d。
2019年茎直黄芪生长期长度在全年增温和生长季增温条件下分别为153.3 d和140.5 d,与对照相比分别缩短了1.7 d和14.5 d(图4B)。
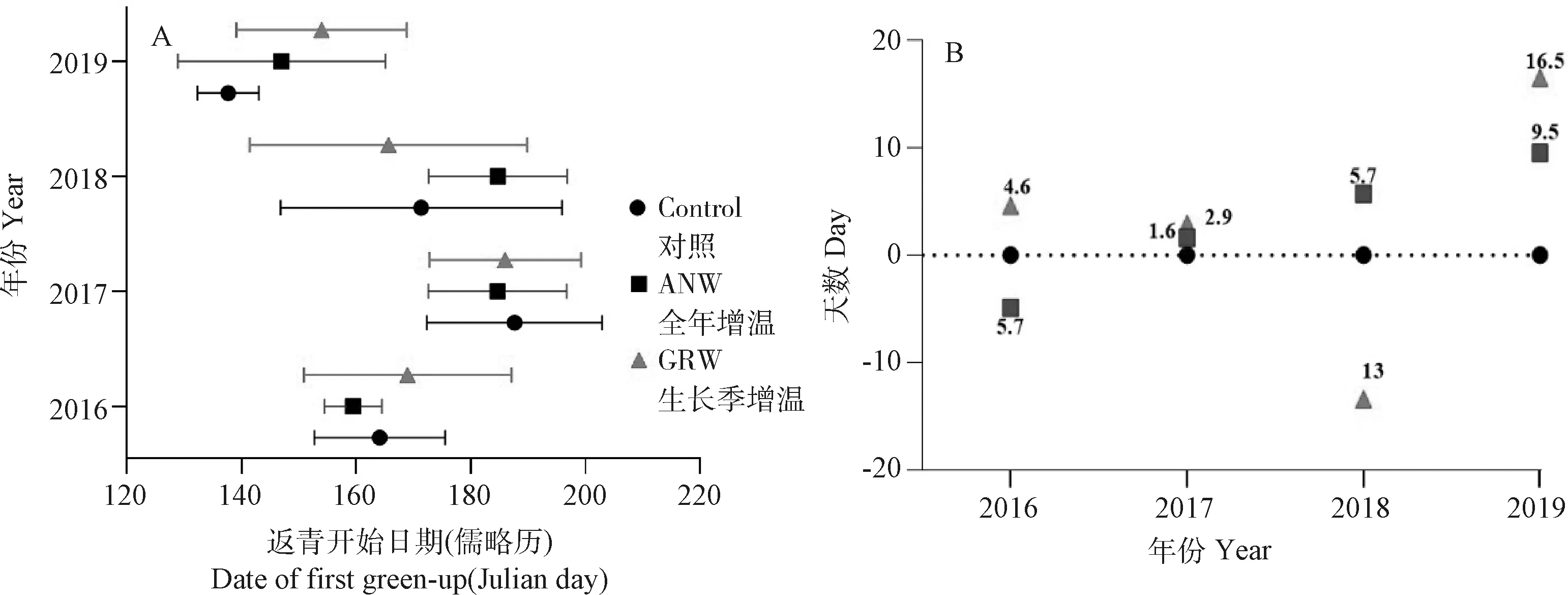
图3 对照、全年增温和生长季增温处理下西藏中部高寒草地植物黄芪的返青物候在年际间的差异
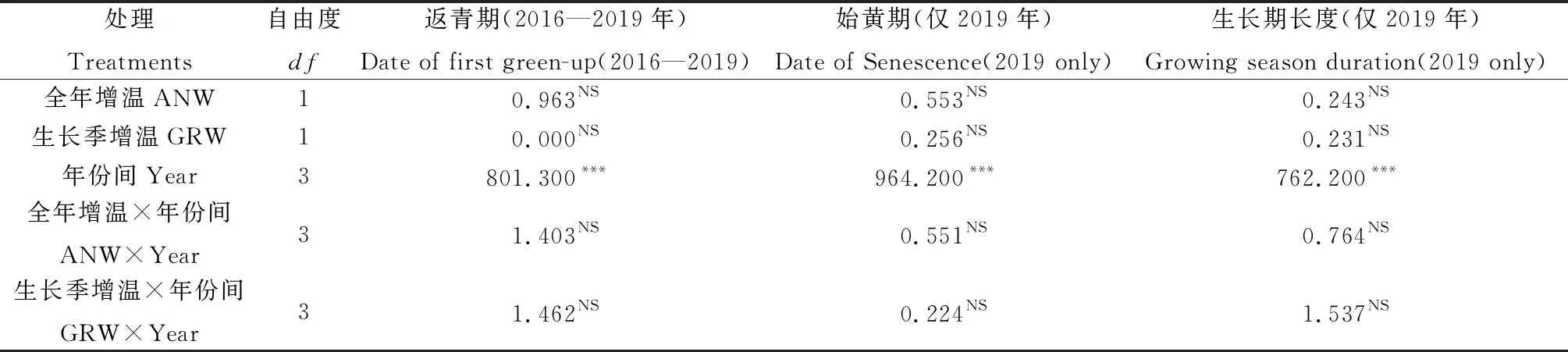
表1 全年增温、生长季增温、年份及其交互作用对西藏中部高寒植物茎直黄芪返青、始黄期和生长期长度的影响
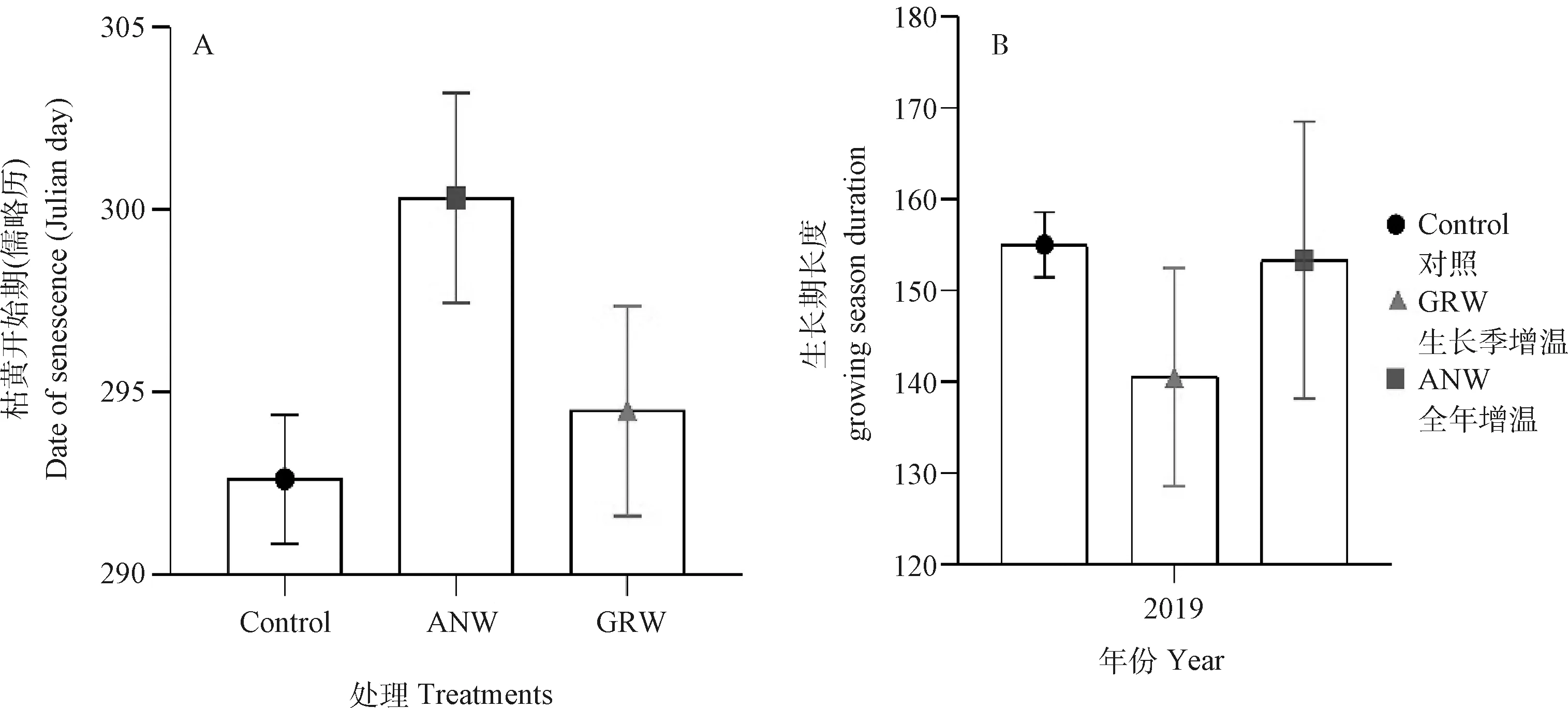
图4 2019年对照、全年增温、和生长季增温处理对茎直黄芪的枯黄期和生长期长度的变化
2.3 微气候条件与茎直黄芪返青物候的相关关系
茎直黄芪返青物候与年均空气温度和生长季空气温度呈显著负相关关系(P<0.05),但与年均、生长季、白天、夜间、午间土壤温度呈显著正相关关系(P<0.05)(表2)。返青物候与土壤湿度各变量无显著相关。基于2019年茎直黄芪的枯黄期和生长期长度与微气候各类变量之间的相关回归分析结果显示,枯黄期和生长期长度与各类微气候变量之间无显著相关关系。
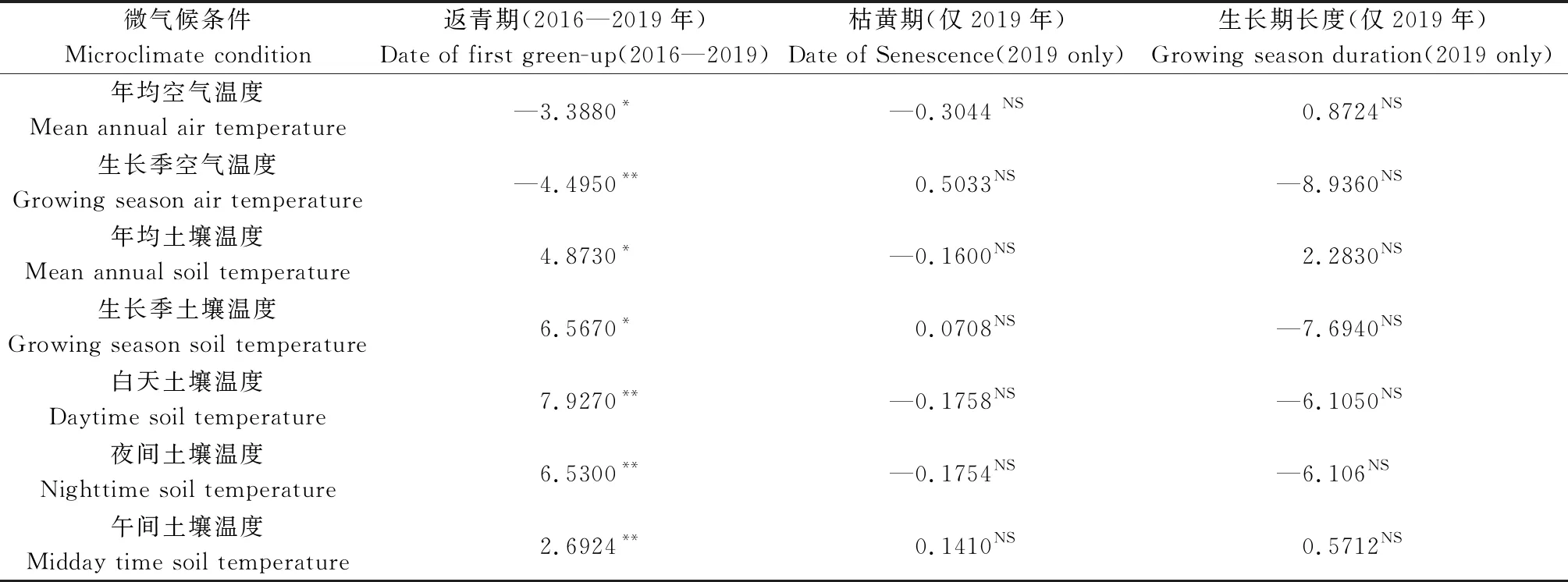
表2 增温处理下微气候条件对返青期、枯黄期和生长期长度的相关性
3 讨论
3.1 茎直黄芪返青物候的年际间差异特征及其机理
相关学者研究表明,温度是植物物候期变化的主要影响因子[25-28]。在气候变暖影响下,极端性干旱气候事件增多[29],导致水分成为限制因素,影响植物物候。本研究表明:茎直黄芪的返青物候在不同年份之间差异显著,总体来讲,2017年返青最晚,2019年则返青最早。这个结果可能与年际间的微气候条件有关。茎直黄芪是晚花植物,在自然情况下返青时间大致在5月下旬,此时自然环境温度已经基本满足其返青需求,但5月下旬雨季未至,因此返青的限制因素是水分而不是温度。2017年土壤和空气温度最高,土壤湿度最低,茎直黄芪因干旱胁迫导致返青延迟。而2019年则相反,由于土壤湿度较高,因此可能导致返青相对较早。以上结果表明,水分条件是影响茎直黄芪返青物候的主要影响因素。这与许多研究结果相似,如在青藏高原其它地区开展的研究结果表明,因气候变暖导致的水热蒸发增大和降水减少是西藏半干旱草甸草原植物返青推迟的主要原因[30];也有研究发现春季降水是青藏高原植物返青的主要影响因素[23];张钛仁等也认为青海环湖地区牧草返青早晚主要受水分条件的限制[31]。
3.2 不同增温处理下茎直黄芪返青与枯黄物候以及生长期长度变化特征及其机理
本研究结果显示,茎直黄芪的返青和枯黄物候在增温处理(包括全年和生长季增温)与对照相比均无显著差异,导致生长期长度也未发生显著变化。由于本试验研究开展于2008年,而本次探讨的茎直黄芪物候观测研究始于2016—2019年(枯黄物候仅2019年),此时处理样圆内的各类植物(包括茎直黄芪)已经经历增温处理8~11年,由于增温幅度较低(全年增温平均增温幅度较对照提高1.19℃,生长季增温较对照增温幅度提高1.15℃),已经逐渐适应了增温条件,导致其对增温效应不敏感。也有研究表明,青藏高原植物返青期对温度敏感性较弱可能与该地区温度波动较大有关,通常温度波动越大的地区,植物为了规避低温冻害,其春季物候对温度的敏感性也会下降[5,31]。另外,还有相关研究指出,随着气温不断升高,春季物候对温度响应的敏感性逐渐降低[32-33]。因此,植物经历了较长时间的增温会逐步适应相应的环境条件,使其对增温处理呈现不敏感趋势。当然,植物物候对气温升高的第一响应能否在较长时间尺度上出现变化还需要更深入的可靠验证。
4 结论
综上,茎直黄芪的植物物候和生长期长度变化特征与趋势主要受水分条件调控。在水分充足条件下温度提升会提前植物的返青物候,延长其枯黄物候,导致生长期延长。然而区域性环境温度较高,环境水分条件相对较低时,增温会使土壤环境的蒸腾加大导致土壤水分减少,植物物候返青延迟,枯黄期提前,生长期长度缩短。
猜你喜欢
农业工程学报(2022年12期)2022-09-09
热带作物学报(2022年6期)2022-07-08
天津农林科技(2020年3期)2020-08-13
水土保持研究(2019年6期)2019-10-19
电子制作(2019年15期)2019-08-27
西南农业学报(2017年5期)2017-06-23
辽宁林业科技(2017年4期)2017-06-22
河北林业科技(2016年5期)2016-11-08
浙江大学学报(工学版)(2016年2期)2016-06-05
高原山地气象研究(2016年4期)2016-02-28
