
基于宿主诱导的基因沉默技术创制抗拟轮枝镰孢玉米自交系
2021-05-12 15:08赫可伟陈甲法周子键吴建宇
中国农业科学 2021年9期
赫可伟,陈甲法,周子键,吴建宇,
基于宿主诱导的基因沉默技术创制抗拟轮枝镰孢玉米自交系
赫可伟1,陈甲法2,周子键2,吴建宇
1河南农业大学农学院,郑州 450002;2河南农业大学生命科学学院,郑州 450002
【】拟轮枝镰孢()是一种主要的病原真菌,侵染玉米可以导致穗粒腐病、茎腐病、苗期根腐病及引起种子腐烂。由拟轮枝镰孢引起的病害不仅影响玉米的产量和品质,而且其病原菌代谢过程中产生的伏马菌素等多种真菌毒素严重威胁了人畜安全。通过宿主诱导的基因沉默(host-induced gene silencing,HIGS)技术创制抗拟轮枝镰孢的玉米种质,为玉米抗病育种提供新的优异抗源。通过同源克隆方法克隆可能与拟轮枝镰孢生长发育相关的关键基因,并通过体外转录获得相应的dsRNA片段;将不同基因的dsRNA与拟轮枝镰孢的分生孢子悬浮液预混后,用于后续的体外RNA沉默试验;对感病玉米自交系西502的种子进行消毒与接种,在培养皿中28℃避光培养48 h,调查种子的发病程度;在混有dsRNA的孢子悬浮液中加入葡萄糖,25℃培养24 h后,在显微镜下观察孢子萌发与菌丝生长情况;将三叶期的西502幼苗转移至预混dsRNA的孢子悬浮液中进行培养,7 d后观察苗期根腐病发病状况;通过种子鉴定与苗期鉴定体系,逐步筛选具有显著抑制效果的沉默靶标基因;合成筛选出的重点靶标基因片段,构建沉默载体并转化感病玉米自交系西502;对转基因株系的种子接种鉴定,验证转化玉米株系的抗性;提取接种后转基因种子的总RNA,对拟轮枝镰孢的靶标基因进行荧光定量分析,确定HIGS株系的沉默效果。从拟轮枝镰孢中克隆出18个与其生长发育相关的候选基因;通过种子接种鉴定,发现11个候选基因被沉默后,种子的发病等级极显著降低;进一步筛选出6个沉默后影响拟轮枝镰孢的孢子萌发和菌丝生长的候选靶标基因、、、、和;通过苗期接种鉴定,最后筛选出3个在体外具有显著抑制效果的沉默靶标基因、和;进而将3个靶标基因的特异区段人工融合成一段序列并构建沉默载体,获得转基因植株;鉴定发现转基因植株的T2代种子对拟轮枝镰孢的抗性显著增强,且3个靶标基因的表达量均显著下降。拟轮枝镰孢基因、、与其生长发育密切相关,且沉默后能够显著提升玉米对拟轮枝镰孢的抗性。
玉米;拟轮枝镰孢;宿主诱导基因沉默;转基因;穗粒腐病
0 引言
【研究意义】由于拟轮枝镰孢引起的玉米病害(如穗粒腐病、茎腐病、苗期根腐病以及种子腐烂等)多数是通过空气与土壤传播,因而缺乏有效的防治措施[1]。拟轮枝镰孢引起的玉米穗粒腐病不仅直接影响玉米的产量和品质,而且其病原菌代谢过程中产生的伏马菌素等多种真菌毒素严重威胁人畜安全[2-3]。越来越多的证据表明,由拟轮枝镰孢属产生的真菌毒素如伏马菌素,可引起马脑白质软化症和猪肺水肿等牲畜疾病[4-5],且与中国部分地区高发的食道癌有关[6-8]。近年来,随着对食品安全问题的普遍关注,玉米穗粒腐病和种子腐烂等问题成为国内外研究的热点和难点问题。防治真菌病害的一个有效手段是克隆主效抗病基因并用于育种改良,ZUO等[9]和WANG等[10]在玉米丝黑穗病、茎腐病等遗传改良中取得了突破性进展,并为抗病育种树立了成功的典范。然而作为微效多基因控制的、由拟轮枝镰孢引起的玉米穗粒腐等病害,图位克隆并聚合抗病基因需要一个较长的时间和过程。因此,通过新的手段创制抗病种质并选育抗拟轮枝镰孢的玉米新品种,对国家的粮食生产安全具有重要意义。【前人研究进展】有关玉米对拟轮枝镰孢的抗性,国内外学者比较系统地研究了玉米穗粒腐病和苗期根腐病抗性。经典数量遗传学研究认为玉米穗粒腐病抗性是由微效多基因控制的数量性状,且以加性效应为主[11]。结合分子标记技术,PÉREZ-BRITO等[12]首次报道了由拟轮枝镰孢引起的玉米穗粒腐病抗性遗传研究成果,利用墨西哥育种材料组配的2个F2:3群体,分别鉴定出9个和7个QTL,其中3个是在2个群体中共同检测到的一致性QTL。北卡罗莱那大学玉米抗病课题组基于美国种质资源抗性鉴定的基础上,利用抗病自交系GE440与感病自交系FR1064构建的BC1F1:2群体及抗病自交系NC300与感病自交系B104构建的重组自交系(recombinant inbred line,RIL)群体,分别鉴定出7个和5个抗穗粒腐病QTL,单个QTL解释了4.4%—5.8%的表型贡献率[13];而意大利的Maschietto等[14]以毒素为鉴定指标,鉴定出一些与穗粒腐病抗性相关的QTL。Septiani等[15]利用401份多亲本高世代互交系(multiparent advanced generation inter-cross,MAGIC)群体,鉴定出10个苗期根腐抗病QTL,并结合转录组分析,预测了候选基因。张帆等[16]在四川省雅安市和绵阳市两地对抗病自交系R15和感病自交系掖478组配的F2群体进行抗性鉴定,在第1、2、3、4、6、7和9染色体上检测出10个抗穗粒腐病QTL,其中第6染色体上的QTL是环境一致性QTL。Ding等[17]利用抗病自交系87-1和感病自交系综3衍生的重组自交系群体,检测到6个抗穗粒腐病QTL,其中在第35染色体上分别检测到2个QTL,在第8和10染色体上分别检测到1个QTL,第3染色体上的和2个QTL是多环境稳定的QTL。近年来,随着基因组测序技术的应用,全基因组关联分析成为了鉴定作物抗病基因的重要手段。Stagnati等[18]利用230份自交系构建了关联群体并筛选出226 446个SNP标记,鉴定出164个与苗期根腐病抗性显著关联的SNP位点及25个相关的抗病候选基因。Zila等[19]利用1 687份美国玉米自交系建立的关联群体及200 978个SNP标记进行分析,鉴定出7个与抗性显著相关的SNP及6个抗病候选基因。Lanubile等[20]对抗病自交系CO441与感病自交系CO354接种拟轮枝镰孢后进行转录组分析,发现茉莉酸和乙烯合成途径与抗性相关,并诱导了多种次级代谢产物的合成。YAO等[21]利用关联分析法鉴定出34个与穗粒腐抗性紧密关联的SNP及69个抗穗粒腐病候选基因,其中包括一些植物激素相关基因及传统抗病蛋白。宿主诱导基因沉默技术是指通过宿主植物生成的RNA诱导病原体基因沉默的方法,是一种基于RNA干涉原理进行种质创新的基因工程技术。它根据病原菌生长或致病相关基因的特异性序列,设计RNAi载体,利用转基因宿主植物生成针对目标病原菌的效应RNA,从而通过跨物种的沉默抑制病原菌的生长达到抗病效果。HIGS技术最早应用在病毒病害的防治上[22-23],1995年在夏威夷证实了HIGS转基因番木瓜“UH-Rainbow”高抗环斑病毒。BAUM等[24]研究表明转V-ATPase A dsRNA的玉米植株可以显著降低玉米根虫病害的危害。这些试验证实了HIGS技术是一种目的性强、效率高的抗病种质创新手段。2007年,Khatri等[25]利用RNAi研究构巢曲霉多胺途径时发现,在构巢曲霉孢子萌发和生长时可以直接从培养基中摄取RNA导致自身沉默,这表明在非人工刺激条件下真菌也可以从生长环境中摄入外源siRNA导致自身沉默。2010年,TinOco等[26]将转入拟轮枝镰孢中,并将表达沉默的RNA序列载体转入烟草中,用转基因拟轮枝镰孢接种正常与转基因烟草叶片,发现只能在正常烟草叶片上检测到发出荧光的菌斑,证实了通过宿主植物表达的dsRNA可以沉默寄生病原菌的基因表达,也预示HIGS可能为抗真菌种质改良提供一种新的育种方案。实践上,Nowara等[27]在2010年通过沉默白粉病致病基因,获得抗白粉病的转基因小麦,证实了可以通过宿主诱导RNA干涉的方法使植物获得病原真菌的抗性。Koch等[28]通过体外RNA沉默试验,发现根据禾谷镰孢设计的一段791碱基的dsRNA序列可以抑制其生长发育,并以此为靶标获得了高抗禾谷镰孢的大麦转基因植株,该研究提出并验证了体外筛选病原真菌发育关键基因作为靶标创制HIGS株系的方案。2017年,THAKARE等[29]利用HIGS技术,成功将黄曲霉毒素合成过程关键基因中的3个片段人工合成在一起构建沉默载体并转入玉米中,第一次成功获得了抑制黄曲霉毒素的玉米植株,在玉米上证实了HIGS技术防治真菌病害的可行性。【本研究切入点】围绕玉米拟轮枝镰孢抗性已经鉴定出一些抗性QTL及候选基因,但多数鉴定的QTL效应较小,进行候选基因的图位克隆与功能研究以实现抗病多基因的聚合需要一个较长的过程,宿主诱导基因沉默技术(host-induced gene silence,HIGS)提供了创制抗病种质的另一种解决方案。【拟解决的关键问题】本研究通过筛选拟轮枝镰孢生长发育关键基因作为靶标,利用HIGS技术创制抗拟轮枝镰孢的新种质,为玉米抗病育种提供新的元件,并为创制优异抗源提供新方案。
1 材料与方法
1.1 拟轮枝镰孢生长发育相关基因的克隆
根据已报道数据,查找与作物致病真菌生长发育或致病性相关的关键基因,运用BLAST比对,确定在拟轮枝镰孢基因组中的同源基因共18个(表1),并克隆与拟轮枝镰孢发育相关的基因序列,作为HIGS沉默抗拟轮枝镰孢的候选靶标基因。
1.2 dsRNA的体外合成
与玉米基因组比对后,选择候选靶标基因内无连续超过20个碱基匹配的特异序列片段设计引物,并在序列两端分别加上T7转录酶的转录识别序列(电子附表1)。通过PCR扩增,获得两端带有T7启动子的体外转录模板。使用HiScribe™ T7 Quick High Yield RNA Synthesis Kit(NEB)试剂盒,对模板进行体外转录,获得候选靶标基因的dsRNA。
1.3 拟轮枝镰孢孢子悬浮液的制备
从自然发病的玉米籽粒上分离拟轮枝镰孢(),经单菌落分离纯化,经PDA培养基培养后,4℃保存备用。接种前将分离纯化的拟轮枝镰孢26℃活化。然后接种到PD培养基,200 r/min培养24 h。收集孢子悬浮液,并用无菌水稀释至1×106个/mL。参考Koch等[28]的体外沉默试验方法,在孢子液中分别加入候选靶标基因的dsRNA至终浓度10 ng·μL-1,用于后续的体外接种试验。
1.4 基于种子抗性鉴定的候选靶标基因筛选
西502玉米自交系是河南农业大学吴建宇课题组鉴定出的多组织多性状高感拟轮枝镰孢的塘四平头血缘自交系[43-44]。挑选整齐一致的西502种子,用75%的酒精表面消毒30 s,然后用3%次氯酸钠消毒15 min,无菌水冲洗3—4次后,将种子接入准备好的孢子液中,以接种水为空白对照,加入的dsRNA为阴性对照。接菌后将种子放在铺入湿滤纸的培养皿中,28℃避光培养。由于早期症状不明显,而后期RNA会在环境中降解,通过预试验确定,48 h后调查20粒种子的发病情况。单粒种子的评价标准为1:无明显菌丝;3:菌丝覆盖面积<25%;5:25%≤菌丝覆盖面积<50%;7:50%≤菌丝覆盖面积<75%;9:菌丝覆盖面积≥75%。
1.5 基于病原菌发育形态观察的候选靶标基因验证
将200 μL混有dsRNA的孢子液移入2.0 mL离心管中,并加入终浓度1%的葡萄糖。25℃培养24 h后,分别取样在显微镜下观察,检测沉默候选靶标基因对拟轮枝镰孢萌发和菌丝生长的抑制效果。
1.6 基于苗期抗性鉴定的靶标基因筛选
参考大田用的孢子悬浮液灌根接种法[45],挑选三叶期的西502幼苗,用纯净水将其根部清洗干净,转移至装有不同dsRNA的30 mL孢子液的三角瓶中进行培养,3个生物学重复,每重复3株。7 d后观察幼苗的苗期根腐病发病状况,进一步筛选高效的沉默靶标基因。根据叶片枯黄状况划分发病等级:1级:没有枯黄面积超过一半的叶片;3级:仅有一片叶片枯黄面积超过一半;5级:2个叶片的枯黄面积超过一半;7级:3个叶片的枯黄面积超过一半但植株未死亡;9级:植株完全枯死。
1.7 HIGS转基因株系的构建和鉴定
在筛选出的关键靶标基因上分别筛选200 bp左右的特异沉默序列,参考抗黄曲霉毒素HIGS株系创制的策略[29],将这些序列融合成一条连续的序列,合并的序列在金维智公司合成并连接到pFGC5941沉默载体上,然后转化感病玉米自交系西502,两代筛选后获得阳性株的T2代种子。参考JU等[46]的方法对筛选的野生型西502和HIGS转基因株系的种子进行接种鉴定,提取接种后3 d的野生型与转基因株系种子的RNA,通过qRT-PCR检测靶基因的表达情况。
2 结果
2.1 拟轮枝镰孢沉默靶标基因的初步筛选
由于拟轮枝镰孢与玉米互作的致病基因报道较少,参考体外筛选病原真菌发育关键基因确定标基因进而创制抗白粉病小麦种质的方案[28],从拟轮枝镰孢中克隆出18个已报道与病原真菌生长发育相关的关键基因(表1)。针对这18个沉默候选靶标基因,选择与玉米基因组相似度较低的沉默片段进行扩增,并在两端加上T7转录酶的启动序列。以扩增产物为模板,通过体外转录获得针对18个候选靶标基因的dsRNA(图1)。
为了筛选有效的沉默靶标基因,将拟轮枝镰孢的孢子液与不同候选靶标基因的沉默片段dsRNA混合后,对感病玉米自交系西502的种子进行接种鉴定。未加入dsRNA的空白对照组平均发病等级为6.3,沉默的阴性对照组平均发病等级为5.9,且与空白对照无显著差异。在加入不同dsRNA的18个处理组中,种子的平均发病等级为2.3—5.3,与对照相比,有5组种子受拟轮枝镰孢的侵染程度显著降低,11组极显著降低(图2)。其中,沉默的处理组种子发病程度最低,平均发病等级为2.3,降低至对照的37%。选择效应极显著的11个对照组的候选靶标基因作为初步筛选的靶标,进行后续试验。
2.2 候选沉默靶标基因的体外验证
利用1%葡萄糖溶液对拟轮枝镰孢孢子进行培养,并分别加入初筛出的11个靶标基因对应的dsRNA,观察孢子萌发及菌丝生长情况。培养24 h后,未加入dsRNA的对照组中拟轮枝镰孢孢子大部分萌发且菌丝生长良好,而11个处理组中有6组拟轮枝镰孢孢子的萌发率和菌丝生长速度明显受到抑制(图3)。初步确认、、、、和可能是参与拟轮枝镰孢的孢子萌发及菌丝生长的关键基因,选择它们进行后续的验证试验。
2.3 高效沉默靶标基因的最终确定
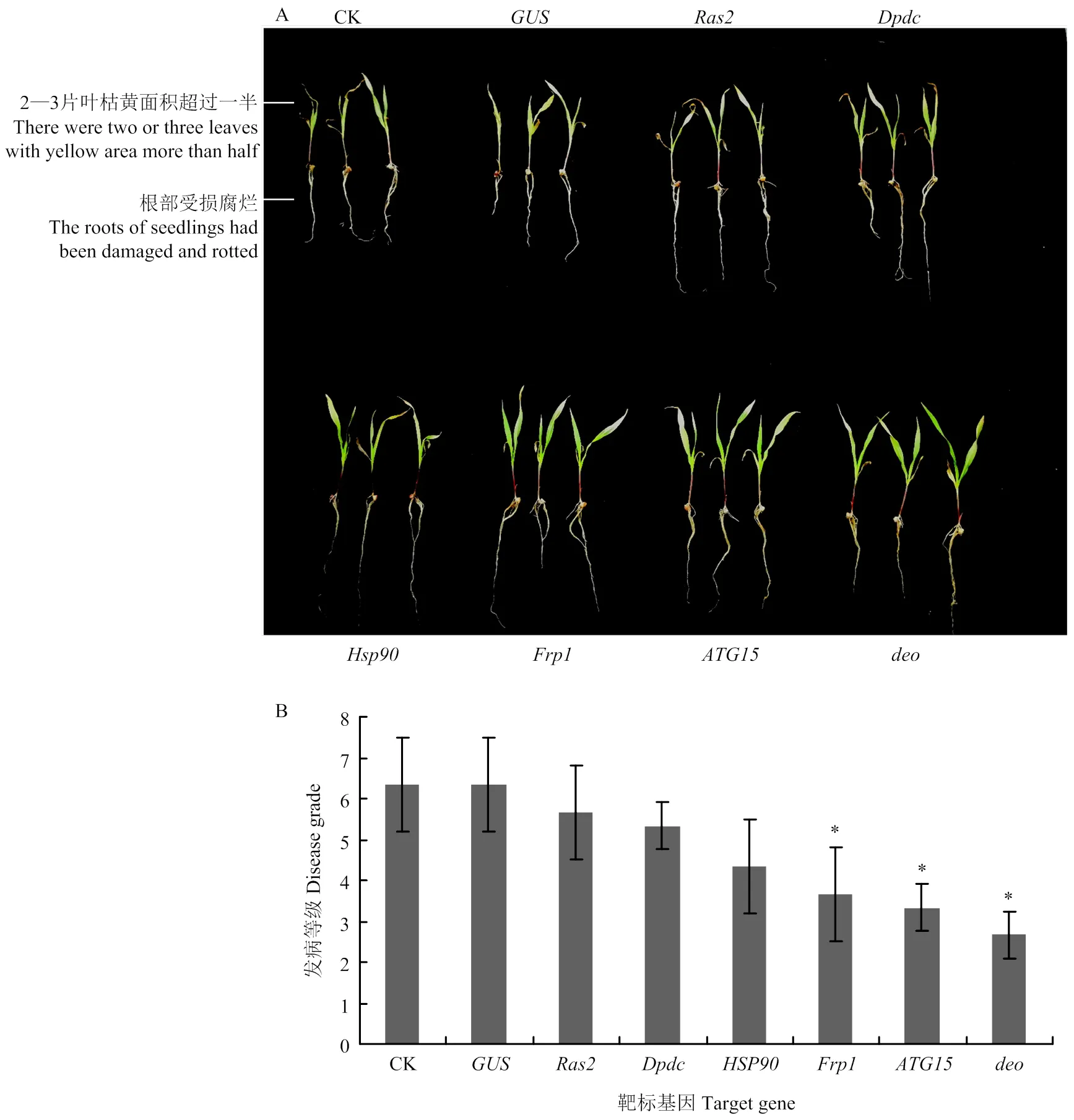
分别用筛选出的6个候选靶标基因及阴性对照的dsRNA与拟轮枝镰孢孢子液混合,并处理玉米幼苗,培养7 d后观察幼苗生长状况。未加入dsRNA的空白对照组和加入dsRNA的阴性对照组中幼苗发病最重,叶片枯死、根部腐烂且植株生长矮小,平均发病等级均为6.33;其他处理组发病状况由重至轻分别为、、、、和的沉默组。其中、和的沉默组中幼苗的长势良好,发病等级分别为3.67、3.33和2.67,发病程度较对照显著降低(图4)。因此,将、和确定为HIGS技术沉默拟轮枝镰孢的最终靶标基因。

*:P<0.05,**:P<0.01。下同The same as below
2.4 抗拟轮枝镰孢玉米转基因株系的构建及验证
分别从、和内选择200、222和200 bp的沉默区段,合并为622 bp长度的人工合成序列(图5)。将该序列连接至沉默载体pFGC5941,并转化感病玉米自交系西502,获得2个阳性转基因T0植株,经筛选两代后获得T2代转基因种子。
对野生型和2个转基因株系的种子进行接种鉴定。在接种拟轮枝镰孢7 d后,野生型种子平均发病等级为4.9,而2个转基因株系分别为3.4和2.8,均极显著低于野生型(图6)。对靶标基因的定量分析结果显示,3个靶基因在2个转基因株系中的表达量为野生型的38.1%—76.3%,均显著低于野生型,达到了宿主诱导的基因沉默效果。研究表明,HIGS转基因株系可以通过抑制拟轮枝镰孢关键基因的表达从而显著提高玉米对轮枝镰孢的抗性。
3 讨论
3.1 基于宿主诱导的基因沉默技术创制抗拟轮枝镰孢玉米种质的策略选择
玉米对拟轮枝镰孢的抗性是多基因控制的数量性状,目前,鉴定出的抗性QTL及候选基因抗病效应较小,通过图位克隆获得抗病基因并进行功能解析从而开发功能标记进行种质创新或抗病基因聚合存在一定难度。以病原菌关键致病基因或生长发育基因为靶标并通过宿主诱导基因沉默的方法创制抗病新种质是另一种值得探索的防治方案。HIGS的方法最早用于防治病毒和昆虫,但近年来也多有成功应用到真菌防治的报道。在此基础上,研究尝试通过HIGS技术获得了抗拟轮枝镰孢的玉米转基因株系,为玉米拟轮枝镰孢病害的抗性种质创制提供了新的方案。
A:接种7 d后的玉米幼苗外观。B:接种7 d后玉米苗期根腐病发病等级的统计

图5 人工合成的靶标序列

A:接种后7 d后野生型和HIGS株系种子的外观。B:接种7 d后野生型和HIGS株系种子的发病等级统计。C:接种3 d后3个靶标基因在野生型和HIGS株系中的相对表达量
HIGS所选用的沉默靶标基因通常为功能明确的病原基因。一般分为两类:一类是直接影响病原真菌致病或产毒能力的基因[27,29];另一类则是病原真菌自身生长发育所必需的基因[28]。在玉米抗黄曲霉毒素HIGS株系创制的研究中,没有检测到HIGS转基因株系的果穗内有黄曲霉毒素,但黄曲霉菌的数量并没有显著减少[29],也就是说抑制毒素合成基因虽然可以降低真菌毒素对人畜的危害,但可能无法降低病原菌对玉米产量造成的损失。由于玉米抗病基因与拟轮枝镰孢致病基因及其互作的研究均较少,因此,选择致病基因最为靶标基因较为困难。基于以上原因,试验选用了病原菌生长发育相关基因作为候选的沉默靶标。
3.2 关键沉默靶标基因的筛选方法
由于难以预测众多靶标基因沉默后的表型以及考虑到转基因在时间与经济上的高昂成本,因此,在创制HIGS转基因株系之前需要对候选靶标基因进行初筛。例如,在小麦的研究中,有时会利用病毒介导的基因沉默技术对靶标基因进行初步的筛选与验证[26]。然而,由于玉米具有丰富的遗传多样性,尚无稳定成熟的瞬时沉默体系可用。Khatri等[25]发现在构巢曲霉孢子萌发和生长时直接从培养基中摄取siRNA导致了自身沉默现象;而Koch等[28]通过dsRNA的体外沉默试验也发现,禾谷镰孢可以被体外的dsRNA沉默从而抑制其生长。这些研究证实了一些真菌可以吸收环境中的RNA并导致自身基因的沉默,为关键靶标基因的体外筛选带来了便利。通过体外合成dsRNA并进行沉默试验,相继从18个真菌生长发育相关基因中筛选出3个高效沉默靶标,并进一步成功创制了HIGS抗病株系。在此过程中,dsRNA在体外对拟轮枝镰孢的沉默效应通过形态学和致病性观察等得到验证,这与Koch等[28]的研究结果吻合。体外dsRNA沉默试验筛选靶标基因的初步探索,为后续的HIGS靶标确定提供了一个便捷的方法。然而,该方法是否适用于其他真菌,以及对dsRNA的长度与浓度有何要求和限制,尚需进一步试验。
3.3 通过同时沉默deo、Atg15和Frp1获得抗拟轮枝镰孢的玉米种质
本研究最终筛选到3个高效沉默靶标基因、和。其中编码一个脱氧辅蛋白合成酶,该基因被抑制后,禾谷镰孢生长减缓,而小麦及玉米对它的抗性显著提高[42]。编码一个与自我吞噬相关的脂肪酶,它被证实影响禾谷镰孢的菌丝生长[37]。在尖孢镰刀菌中编码一个与致病性相关的F-box蛋白,发现它与拟轮枝镰孢的生长可能相关[39]。由于这3个基因在体外沉默试验中均有较好的抑菌效果,因此,本研究探索了同时沉默这3个靶标基因的设计。RNA干扰的机制是匹配后的dsRNA在体内dicer酶或其同工酶的作用下切割成18 bp左右的小片段,进而沉默目标基因并产生二次扩散。因此,沉默载体中不需要表达完整的蛋白编码序列,仅需要表达目标基因的部分序列,转录出的RNA被切割成不同siRNA后共同沉默目标基因。以往研究发现,在同一个表达框中表达基因不同片段的人工合成序列,可以提高目标基因的沉默效率[29]。在此启发下,试验将3个靶标基因的部分序列人工合成在一段序列之中并构建沉默载体,成功创制了HIGS抗病株系,并同时检测到HIGS株系对这3个拟轮枝镰孢基因的沉默效果,为今后多基因沉默的转基因株系设计提供了方案。
4 结论
拟轮枝镰孢基因、和对拟轮枝镰孢的孢子萌发和生长发育至关重要,其转基因HIGS植株对玉米苗期根腐病、茎腐病、穗粒腐病有一定的抗性。
[1] 王晓鸣, 晋齐鸣, 石洁, 王作英, 李晓. 玉米病害发生现状与推广品种抗性对未来病害发展的影响. 植物病理学报, 2006, 36(1): 1-11.
WANG X M, JIN Q M, SHI J, WANG Z Y, LI X. The status of maize diseases and the possible effect of variety resistance on disease occurrence in the future. Acta Phytopathologica Sinica, 2006, 36(1): 1-11. (in Chinese)
[2] KAMLE M, MAHATO D K, DEVI S, LEE K E, KANG S G, KUMAR P. Fumonisins: Impact on agriculture, food, and human health and their management strategies. Toxins, 2019, 11(6): 328.
[3] Knutsen H K, Alexander J, Barregård L, Bignami M, Brüschweiler B, Ceccatelli S, Cottrill B, Dinovi M, Edler L, Grasl-Kraupp B, Hogstrand C, Hoogenboom L, Nebbia C S, Petersen A, Rose M, Roudot A C, Schwerdtle T, Vleminckx C, Vollmer G, Wallace H, Dall'Asta C, Eriksen G S, Taranu I, Altieri A, Roldán- Torres R, Oswald I P. Risks for animal health related to the presence of fumonisins, their modified forms and hidden forms in feed. EFSA Journal, 2018, 16(5): 5242.
[4] ROSS P F, RICE L G, PLATTNER R D, OSWEILER G D, WILSON T M, OWENS D L, NELSON H A, RICHARD J L. Concentrations of fumonisin B1 in feeds associated with animal health problems. Mycopathologia, 1991, 114(3): 129-135.
[5] MARASAS W F O. Fumonisins: Their implications for human and animal health. Natural Toxins, 1995, 3(4): 193-198.
[6] GELDERBLOM W C A, JASKIEWICZ K, MARASAS W F O, THIEL P G, HORAK R M, VLEGGAAR R, KRIEK N P. Fumonisins: novel mycotoxins with cancer-promoting activity produced by. Applied & Environmental Microbiology, 1988, 54(7): 1806-1811.
[7] YOSHIZAWA T, YAMASHITA A, LUO Y. Fumonisin occurrence in corn form high-risk and low-risk areas for human esophageal cancer in china. Applied and Environmental Microbiology, 1994, 60(5): 1626-1629.
[8] Ueno Y, Iijima K, Wang S D, Sugiura Y, Sekijima M, Tanaka T, Chen C, Yu S Z. Fumonisins as a possible contributory risk factor for primary liver cancer: A 3-year study of corn harvested in Haimen, China, by HPLC and ELISA. Food and Chemical Toxicology, 1997, 35(12): 1143-1150.
[9] ZUO W, CHAO Q, ZHANG N, YE J, TAN G, LI B, XING Y, ZHANG B, LIU H, FENGLER K A, ZHAO J, ZHAO X, CHEN Y, LAI J, YAN J, XU M. A maize wall-associated kinase confers quantitative resistance to head smut. Nature genetics, 2015, 47(2): 151-157.
[10] WANG C, YANG Q, WANG W, LI Y, GUO Y, ZHANG D, MA X, SONG W, ZHAO J, XU M. A transposon-directed epigenetic change inunderlies quantitative resistance to Gibberella stalk rot in maize. The New phytologist, 2017, 215(4): 1503-1515.
[11] LUNSFORD J N, FUTRELL M C, SCOTT G E. Maternal influence on response of corn to. Phytopathology, 1974, 65: 223-225.
[12] PÉREZ-BRITO S, JEFFERS D, GONZÀLEZ-DE-LEÓN D, KHAIRALLAH M, CORTÉS-CRUZ M, VELÀZQUEZ-CARDELAS G, AZPIROZ-RIVERO S, SRINIVASAN G. QTL mapping ofear rot resistance in highland maize, México. Agrociencia, 2001, 35: 181-196.
[13] ROBERTSON-HOYT L A, JINES M P, BALINT-KURTI P J, KLEINSCHMIDT C E, WHITE D G, PAYNE G A, MARAGOS C M, MOLNÁR T L, HOLLAND J B. QTL mapping for Fusarium ear rot and fumonisin contamination resistance in two maize populations. Crop Science, 2006, 46(4): 1734-1745.
[14] MASCHIETTO V, COLOMBI C, PIRONA R, PEA G, STROZZI F, MAROCCO A, ROSSINI L, LANUBILE A. QTL mapping and candidate genes for resistance to Fusarium ear rot and fumonisin contamination in maize. BMC Plant Biology, 2017, 17(1): 20.
[15] SEPTIANI P, LANUBILE A, STAGNATI L, BUSCONI M, NELISSEN H, MARIO ENRICO P, DELL’ACQUA M, MAROCCO A. Unravelling the genetic basis of Fusarium seedling rot resistance in the MAGIC maize population: novel targets for breeding. Scientific reports, 2019, 9(1): 5665.
[16] 张帆, 万雪琴, 潘光堂. 玉米抗穗粒腐病QTL定位. 作物学报, 2007, 33(3): 491-496.
ZHANG F, WAN X Q, PAN G T. Molecular mapping of QTL for resistance to maize ear rot caused by. Acta Agronomica Sinica, 2007, 33(3): 491-496. (in Chinese)
[17] DING J Q, WANG X M, CHANDER S, YAN J B, LI J S. QTL mapping of resistance to Fusarium ear rot using a RIL population in maize. Molecular Breeding, 2008, 22(3): 395-403.
[18] STAGNATI L, LANUBILE A, SAMAYOA L F, BRAGALANTI M, GIORNI P, BUSCONI M, HOLLAND J B, MAROCCO A. A genome wide association study reveals markers and genes associated with resistance toinfection of seedlings in a maize diversity panel. Genes, Genomes, Genetics, 2019, 9(2): 571-579.
[19] ZILA C T, OGUT F, ROMAY M C, GARDNER C A, BUCKLER E S, HOLLAND J B. Genome-wide association study of Fusarium ear rot disease in the U.S.A. maize inbred line collection. BMC Plant Biology, 2014, 14: 372.
[20] LANUBILE A, FERRARINI A, MASCHIETTO V, DELLEDONNE M, MAROCCO A, BELLIN D. Functional genomic analysis of constitutive and inducible defense responses toinfection in maize genotypes with contrasting ear rot resistance. BMC Genomics, 2014, 15(1): 710.
[21] YAO L, LI Y, MA C, TONG L, DU F, XU M. Combined genome-wide association study and transcriptome analysis reveal candidate genes for resistance to Fusarium ear rot in maize.Journal of Integrative Plant Biology, 2020, 62(10): 1535-1551.
[22] SUDARSHANA M R, ROY G, FALK B W. Methods for engineering resistance to plant viruses. Methods in Molecular Biology, 2007, 354: 183-195.
[23] WATERHOUSE P M, FUSARO A F. Viruses face a double defense by plant small RNAs. Science, 2006, 313(5783): 54-55.
[24] BAUM J A, BOGAERT T, CLINTON W, HECK G R, FELDMANN P, ILAGAN O, JOHNSON S, PLAETINCK G, MUNYIKWA T, PLEAU M, VAUGHN T, ROBERTS J. Control of coleopteran insect pests through RNA interference. Nature Biotechnology, 2007, 25(11): 1322-1326.
[25] Khatri M, Rajam M V. Targeting polyamines ofsiRNA specific to fungal ornithine decarboxylase gene. Medical Mycology, 2007, 45(3): 211-220.
[26] TINOCO M, BÁRBARA D, DALL'ASTTA R, JOÃO P, ARAGÃO F.trans-specific gene silencing in fungal cells by in planta expression of a double-stranded RNA. BMC Biology, 2010, 8: 27.
[27] NOWARA D, GAY A, LACOMME C, SHAW J, RIDOUT C, DOUCHKOV D, HENSEL G, KUMLEHN J, SCHWEIZER P. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen. The Plant Cell, 2010, 22(9): 3130-3141.
[28] KOCH A, KUMAR N, WEBER L, KELLER H, IMANI J, KOGEL K H. Host-induced gene silencing of cytochrome P450 lanosterol C14 -demethylase-encoding genes confers strong resistance to. Proceedings of the National Academy of Sciences of the USA, 2013, 110(48): 19324-19329.
[29] THAKARE D, ZHANG J, WING R A, COTTY P J, SCHMIDT M A. Aflatoxin-free transgenic maize using host-induced gene silencing.Science Advances, 2017, 3(3): e1602382.
[30] BLUHM B H, ZHAO X, FLAHERTY J E, XU J R, DUNKLE L D.regulates growth and pathogenesis in. Molecular Plant-Microbe Interactions, 2007, 20(6): 627-636.
[31] JIANG J, LIU X, YIN Y, MA Z. Involvement of a velvet protein FgVea in the regulation of asexual development, lipid and secondary metabolisms and virulence in. PLoS ONE, 2011, 6(11): e28291.
[32] ADÁM A L, KOHUT G, HORNOK L., a hog-type map kinase gene, is involved in multistress response in. Journal of basic microbiology, 2010, 48(3): 151-159.
[33] COLABARDINI A C, BROWN N A, SAVOLDI M, GOLDMAN M H S, GOLDMAN G H. Functional characterization of, a homologue of the mammalian kinase SGK. PLoS ONE, 2013, 8(3): e57630.
[34] CHOI Y E, SHIM W B. Functional characterization of, a gene encoding a putative protein phosphatase 2A catalytic subunit. Microbiology, 2008, 154(1): 326-336.
[35] EATON C J, CABRERA I E, SERVIN J A, WRIGHT S J, COX M P, BORKOVICH K A. The guanine nucleotide exchange factor RIC8 regulates conidial germination through Gα proteins in. PLoS ONE, 2012, 7(10): e48026.
[36] VIJAI B, SABINE B, ALBERT V, GOPALAN S, WEI Y D. Alanine: glyoxylate aminotransferase 1 is required for mobilization and utilization of triglycerides during infection process of the rice blast pathogen,. Plant Signaling & Behavior, 2012, 7(9): 1206-1208.
[37] NGUYEN L N, JÖRG B, LE G T, STÄRKEL C, SCHÄFER W. Autophagy-related lipase FgATG15 ofis important for lipid turnover and plant infection. Fungal Genetics and Biology, 2011, 48(3): 217-224.
[38] ZHANG H, GUO J, VOEGELE R T, ZHANG J, DUAN Y, LUO H, KANG Z. Functional characterization of calcineurin homologs pscna1/pscnb1 inf sp tritici using a host-induced rnai system. PLoS ONE, 2012, 7(11): e49262.
[39] DUYVESTEIJN R G E, WIJK R V, BOER Y, REP M, HARING M A. Frp1 is af-box protein required for pathogenicity on tomato. Molecular Microbiology, 2005, 57(4): 1051-1063.
[40] LAMOTH F, JUVVADI P R, FORTWENDEL J R, STEINBACH W J. Heat shock protein 90 is required for conidiation and cell wall integrity in. Eukaryotic Cell, 2012, 11(11): 1324-1332.
[41] ZHAO P B, REN A Z, XU H J, LI D C. The gene, encoding a camp-dependent protein kinase catalytic subunit homolog, is required for hyphal growth, spore germination, and plant infection in. Journal of Microbiology & Biotechnology, 2010, 20(1): 208.
[42] WORIEDH M, HAUBER I, MARTINEZ-ROCHA A L, VOIGT C, MAIER F J, SCHRÖDER M, MEIER C, HAUBER J, SCHÄFER W. Preventing fusarium head blight of wheat and cob rot of maize by inhibition of fungal deoxyhypusine synthase. Molecular Plant-Microbe Interactions, 2011, 24(5): 619-627.
[43] CHEN J, DING J, LI H, LI Z, SUN X, LI J, WANG R, DAI X, DONG H, SONG W, CHEN W, XIA Z, WU J. Detection and verification of quantitative trait loci for resistance to Fusarium ear rot in maize. Molecular Breeding, 2012, 30(4): 1649-1656.
[44] MU C, GAO J, ZHOU Z, WANG Z, SUN X, ZHANG X, DONG H, HAN Y, LI X, WU Y, SONG Y, MA P, DONG C, CHEN J, WU J. Genetic analysis of cob resistance to: another step towards the protection of maize from ear rot. Theoretical and applied genetics, 2019, 132(4): 1049-1059.
[45] DOLORES B R, FERNANDO C G, CARLOS L G A, ANTONIO R H, VICTOR H A R, JOSE L C S. Responses of maize landrace seedlings to inoculations ofspp. Open Access Library Journal, 2017, 4(6): 1-14.
[46] JU M, ZHOU Z, MU C, ZHANG X, GAO J, LIANG Y, CHEN J, WU Y, LI X, WANG S, WEN J, YANG L, WU J. Dissecting the genetic architecture of Fusarium verticillioides seed rot resistance in maize by combining QTL mapping and genome-wide association analysis. Scientific Reports, 2017, 7: 46446.
resistant maize inbred line development using host-Induced gene silencing technology
HE Kewei1, CHEN Jiafa2, ZHOU Zijian2, WU Jianyu1,2
1College of Agronomy, Henan Agricultural University, Zhengzhou 450002;2College of Life Sciences of Henan Agricultural University, Zhengzhou 450002
【】() is a common pathogen, which can cause ear rot, stalk rot, seedling blight, and seed rot in maize. These diseases caused bynot only affected the yield and quality of maize, but also seriously threatened to the safety of human and livestock by a variety of fungal toxins such as fumonisin which produced during the metabolic process of the pathogen. So far, there is no report about the major resistance gene cloned and utilized forin maize. Using host-induced gene silencing technology provides a new strategy for resistance breeding in maize. 【】 Key genes associated with thedevelopment were cloned using homologous gene sequence method, and the dsRNA were produced by in-vitro transcription (IVT) assay. The dsRNA for different genes was premixed with suspension spores ofused for RNA silencing experiment in vitro. For investigate the degree of disease, the seeds of the susceptible inbred line Xi502 were sterilized and inoculated, and then were cultured in a petri dish at 28℃ in the dark for 48 h. For investigate the incidence of the seeds after inoculation, glucose was added to the spore suspension mixed with dsRNA, then spore germination and mycelia growth were observed under the microscope after 25℃ culture for 24 h. The Xi502 seedlings of the trifoliate stage were transferred to the spore suspension with premixed dsRNA for culture, and the incidence of seedlings blight was observed after 7 days. In order to select the target gene for HIGS, combine the seed morphological observation result after inoculation and seedling inoculation result. Then, the silent vector about these key target genes were constructed and transferred into the susceptible inbred line Xi502. The transgenic seeds were evaluated by artificial inoculation. The total RNA of the transgenic seeds after inoculation was extracted, and the relative expression of target genes inwas analyzed by qRT-PCR to determine the silencing effect of HIGS line. 【】Eighteen candidate genes related to growth were cloned by homologous cloning method in. It was found that the disease level of seeds was significantly reduced after 11 candidate genes silencing by seed inoculation experiments. Furthermore, six of the 11 candidate target genes,,,,,, and, were found that response to the spore germination and mycelium growth after gene silencing. Finally, based on the results of seedling inoculation, 3 silencing target genes,andwith significant inhibitory effect in vitro were selected. Then the silencing vector was constructed by combine three specific segments from the three target genes, transgenic plants were obtained. It was found that the resistance level was highly increased in T2-generation seeds compared to the none-transgenic plants. As well as the expression levels of all the three target genes were significantly decreased in. 【】Three genes,,and, are important for development of. By constructing transgenic HIGS plants for target gene,and, the increase the resistance toin maize.
maize (L);; host-induced gene silencing; transgenic; ear rot

10.3864/j.issn.0578-1752.2021.09.002
2020-03-16;
2020-05-21
国家自然科学基金(31761143009,U1704105)
郝可伟,E-mail:544220111@qq.com。通信作者周子键,E-mail:zhouzijian19900601@163.com。通信作者吴建宇,E-mail:wujianyu40@126.com
(责任编辑 李莉)
猜你喜欢
今日农业(2022年4期)2022-11-16
中国种业(2022年8期)2022-08-19
种子(2022年6期)2022-08-03
火力与指挥控制(2022年3期)2022-04-27
北京农学院学报(2022年1期)2022-03-10
种子(2021年6期)2021-07-16
军民两用技术与产品(2021年10期)2021-03-16
上海师范大学学报·自然科学版(2020年5期)2020-12-18
江苏农业科学(2020年14期)2020-08-28
师道·教研(2019年7期)2019-08-13
