
条斑紫菜的种质创新与应用*
2021-05-24 11:39胡传明陆勤勤许广平杨立恩田翠翠邓银银
广西科学院学报 2021年1期
周 伟,胡传明,陆勤勤,许广平 ,杨立恩,田翠翠,邓银银**
(1.江苏省海洋水产研究所,江苏南通 226007;2.江苏省农业种质资源保护与利用平台,江苏南京 210014)
0 引言
紫菜(Pyropia)隶属于红藻门(Rhodophyta)红藻纲(Rhodophyceae)红毛菜亚纲(Bangiophycidae)红毛菜目(Bangiales)红毛菜科(Bangiaceae),是一种广温性海藻,分布于南、北半球的寒带至热带地区[1]。紫菜一般生长于沿海潮间带的岩礁上,或附生在其他海藻上[2]。据报道,世界上约有134种紫菜。目前我国已记载有24个种或变种,其中主要栽培种类为北方的条斑紫菜(Pyropiayezoensis)和南方的坛紫菜(Pyropiahaitanensis)。紫菜的大规模人工栽培不仅具有重要的经济价值,而且还可以保持近海养殖生态平衡及碳循环,创造重要的生态效益[3]。但是,近年来由于全球气候变化以及栽培密度过大等问题,条斑紫菜栽培产业的稳定性和可持续发展受到影响。而优良种质是条斑紫菜规模化产业健康发展的基石,因此,本文对条斑紫菜良种培育及其应用进行回顾与分析,并探讨了其种质创新的新思路。
1 紫菜栽培现状
紫菜是世界上最重要的人工栽培海藻之一,目前在全球范围内有20多亿美元的产业规模[1]。中国、日本和韩国是紫菜的主要栽培国家。根据联合国粮食及农业组织(Food and Agriculture Organization of the United Nations,FAO)统计的2000-2017年紫菜生产数据,中国紫菜年产量逐年增加,至2017年年产量达到1.7×106t,年产值达到1.1×106美元,是目前最大的紫菜生产国。韩国近几年的紫菜产量和产值也在逐步增加,至2017年年产值达到5.4×105美元。近二十年里,日本紫菜年产量十分稳定(约3×105t),年产值高达8.3×105美元,为单价最高的紫菜生产国(图1)。

图1 中国、日本和韩国在2000-2017年紫菜的产量和产值(数据来自FAO)
条斑紫菜(Pyropiayezoensis)作为中国、日本和韩国的主要紫菜栽培品种,其产品的加工具有较高的经济价值[2]。在我国,条斑紫菜栽培生产是我国长江以北人工栽培海藻中最具经济价值的产业[3]。条斑紫菜品质优良,年产量6 000 t左右,其产值占我国紫菜总产值的一半左右。江苏省作为条斑紫菜主产区,历经三十多年的发展过程,无论是栽培面积、产量,还是配套产业规模都已占据全国的95%以上,其中栽培面积约2.1×104hm2(年平均递增率1.6%),育苗室面积约3.2×105m2,各类作业船500艘,拖拉机、收获机等专业机械1 000台以上,各类加工机组600多套(台),干紫菜加工机组523台,二次加工生产线100条以上,从业人员5万多人,行业总产值30亿元左右[4]。
随着生产规模的扩大,条斑紫菜产业目前面临许多问题,急需研究解决,而条斑紫菜栽培产业的健康稳定发展有赖于良种的选育与应用。因此,中国、日本和韩国为满足条斑紫菜产业规模化栽培和应用,根据不同的育种目标需求,通过一定的育种手段培育出一些具有优良性状的条斑紫菜新品种。
2 育种目标
紫菜育种是筛选培育出具有优良性状的、且能稳定遗传的紫菜新品种。随着栽培生产和市场发展对条斑紫菜产品的多样化需求,其育种目标也随之发生改变,大致可分为4个目标,下文详细介绍。
2.1 以产量为目标
紫菜的良种培育始于20世纪70年代,研究者发现栽培的紫菜种质质量性状退化,所以产量是当时的主要育种目标[5]。在品种选育中,紫菜叶状体的生长速度是鉴定品种产量的一个重要指标。中国科学院黄海水产研究所专家培育出一种新的长叶形条斑紫菜品种,海区试验显示其比原有养殖品种生长快,增产幅度约为15%[6]。Lee等[7]和Ma等[8]分别分离出生长速度快的新品系PyE2和E,在实验室条件下,两者生长速度均显著高于各自对应的野生型,且该性状稳定遗传。Park等[9]对一株快速生长的品系Py2K进行转录组分析,发现紫菜生长速度加快很可能是由于藻体对氮源的吸收能力增加而引起的。
此外,无性生殖的单孢子是条斑紫菜栽培不可缺少的重要种苗来源。据观察,人工采苗附着的壳孢子苗下海后会因种种影响因素不断脱落,最终在栽培网上形成稳定幼苗群体的主要是单孢子苗群[10]。所以条斑紫菜的产量不仅取决于藻体的生长速度,而且与是否形成合理的单孢子苗群息息相关。例如,我国的条斑紫菜新品种“苏通1号”和“苏通2号”单孢子放散条件较宽,易形成合理的幼苗种群结构,比传统栽培品系增产15%以上,显示出高产、稳产的特点[11,12]。
2.2 以品质和营养为目标
随着中国、日本和韩国紫菜产业竞争激烈,市场对紫菜产品的品质和营养提出了更高的要求[13]。因此,藻体的品质和营养成分含量成为鉴定条斑紫菜优异种质的又一关键指标。条斑紫菜品质的好坏主要取决于3种光合色素和蛋白(叶绿素a、藻红蛋白和藻蓝蛋白)的含量[14],是条斑紫菜优良品系重要的筛选指标[15-17]。紫菜的主要营养成分有可溶性蛋白、游离氨基酸和脂肪酸等[18],因此在条斑紫菜新品种“苏通1号”和“苏通2号”的初期选育中,藻体的可溶性蛋白、游离氨基酸、总脂肪酸、不饱和脂肪酸成分和含量是营养成分的筛选指标[11,12];在不同海区的栽培试验中,通过品质和营养成分分析可筛选出合适条斑紫菜“黄优1号”生长的栽培海区[19]。
2.3 以抗逆性为目标
近十几年来,受全球气候变暖的影响,紫菜栽培海区的水温随之升高,紫菜的萌发与生长受到影响,严重时会出现大规模掉苗和烂苗现象,给紫菜生产带来巨大的经济损失[20]。因此,培育耐高温紫菜品种对紫菜产业的可持续发展具有重要意义。Shin等[21]、黄文等[22]分别培育出耐高温品系HTR-1、T-17,在高温条件下,藻体的生长、色素含量、光合效率以及单孢子释放量都显著优于野生型。Ding等[23]分离出一株耐高温条斑紫菜品系YZ-4,其F1代的藻体在高温下的生长速度显著高于野生型。Lee等[24]分离出一株耐高温品系Py500G,通过蛋白组学研究发现,在高温条件下耐高温品系的光合作用、能量代谢和氧化还原稳态蛋白质水平增加,从而使得紫菜抗逆性能力增强。
2.4 以抗病害为目标
由于海洋酸化、污染以及栽培密度不合理等原因,条斑紫菜在栽培期易受到细菌、真菌的感染,导致赤腐病、壶状菌病和绿斑病等3种主要病害频发,严重影响条斑紫菜的产业发展[25]。2013年,在韩国Seocheon的紫菜养殖区大规模爆发了一种由壶状菌(Olpidiopsisspp.)引起的壶状菌病,造成大约160万美元的损失,占该地区2013-2014年度紫菜总销售额的10.7%[26]。由腐霉菌(Pythiumchondricola)引起的赤腐病和由PyroV1病毒(Pyropia-infecting virus 1)引起的绿斑病,也会给紫菜产业带来巨大损失[27,28],因此培育抗病害的优异种质迫在眉睫。近期,韩国藻类研究者Park等[29]从感染赤腐病的条斑紫菜中分离出存活的营养细胞,成功培育出两株抗赤腐病的新品种AP1和AP2,这两个新品种在实验室条件下对腐霉菌产生部分稳定的抗性,有望成为抗赤腐病病害的优异种质。
3 种质创新方法
紫菜育种技术是通过人为的方式,固定或改变紫菜的遗传性状,培育出能稳定遗传且具有优良性状的新品种。条斑紫菜育种技术主要有3种,包括选择性育种、诱变育种和杂交育种。
3.1 选择性育种
选择性育种技术,是通过从野生群体或栽培群体中选取具有优良性状的个体作为亲本,通过连续几代的栽培,重复选择性状优良的个体,直至形成稳定遗传的良种[30]。20世纪70年代,日本最早开展条斑紫菜的选育研究,在紫菜栽培群体中筛选出长约40 cm、宽15-20 cm的个体,经过数年反复筛选获得具有稳定遗传性状的栽培品种“奈良轮条斑紫菜” (P.yezoensisf.narawaensis),对日本紫菜产业的发展起到重要推动作用[31,32]。我国条斑紫菜的人工栽培始于20世纪70年代,早期栽培生产和良种选育的主要种源均来自于山东青岛海域采集的条斑紫菜自然种群。20世纪80年代初,中国科学院黄海水产研究所专家根据前述日本学者的方法选育出1个长叶形条斑紫菜新品种[6]。近期,韩国藻类研究者Park等[29]通过选择性育种方法培育出两株抗赤腐病的新品种AP1和AP2。
在条斑紫菜选择性育种中,藻体表型性状易于观察和挑选,且选育过程在海区进行,能表现出较好的海区适应性,在早期可以较为快速地选育出适宜生产栽培的新品种。
3.2 诱变育种
诱变育种技术是指利用物理或化学等诱变方法,诱发植物组织细胞产生基因突变,再从突变体中筛选出在藻体生长和生理生化等方面具有优势的个体过程[33]。关于紫菜人工诱变育种所使用的诱变剂主要有化学诱变剂N-甲基-N′-硝基-N-亚硝基胍(MNNG)、甲基磺酸乙酯(EMS),以及物理诱变剂γ-射线、重离子束等。
在化学诱变方面,许璞[34]和王金峰等[35]利用MNNG处理条斑紫菜的叶状体、营养藻丝和壳孢子,均可产生色素突变体,而这些色素突变体可用于后续的育种工作。Lee等[7]和Ma等[8]均采用EMS对条斑紫菜的叶状体进行诱变处理,分别分离出一种生长速度快的品系。在物理诱变方面,γ-射线因穿透力强、诱发突变率高、周期短等优点而被广泛应用。Wang等[36]证实60Co-γ射线对条斑紫菜叶状体的诱变效果较好。随后利用该技术,江苏省海洋水产研究所联合常熟理工学院研究培育出中国第一、第二个条斑紫菜新品种“苏通1号”和“苏通2号”。另外,Niwa等利用重离子束诱变条斑紫菜并成功获得色素突变体[37],随后利用重离子束进一步诱导该突变体,获得可以萌发形成嵌合体的单孢子[38]。
诱变育种随机性大,诱变方向和性质不能控制,较难实现有目的的调控,但能在短时间内获得大量突变体,将其与人工诱变、逆境胁迫相结合,也可较容易地选育出紫菜抗逆品系。
3.3 杂交育种
杂交育种技术是将不同基因型的紫菜品系杂配,从杂交种中选取在生长、生化方面具有优势的新品种的方法。该技术的优势是能有目的地将亲本的优良性状集中到杂交后代中,发挥杂交重组优势,产生在性状上超过亲本的类型。日本藻类学家Miura等[31]利用色素突变体杂交培育新品种,成功培育出色泽优、产量高的第一个人工紫菜新种质(日本国品种登录名“晓光”),成为20世纪90年代中后期日本最为盛行的栽培品种。刘美君等[15]将野生型条斑紫菜和红色突变体杂交选育,分离出1个耐高温的重组优良品系(HW-4)。唐隆晨等[16]和江灏等[17]将条斑紫菜红色突变体和绿色突变体杂交,在实验室分别选育出一种生长速度快、品质优的新品系(LC-14和A-18),有望在生产中应用。
4 种质应用
中国、日本和韩国作为条斑紫菜栽培的主产国,各自培育了许多条斑紫菜新品种,用以满足不同海区、栽培方式、市场等各方面的发展。20世纪90年代以来,我国藻类学工作者在紫菜育种学研究方面取得多项科技成果。特别是1999年农业部批准的“国家级紫菜种质库”的建设运行,在总结我国紫菜育种的技术与成果的基础上,建立了紫菜种质制备、保存与应用的技术方法和规范,并获得了一批稳定的育种材料。2014年与2015年,由江苏省海洋水产研究所和常熟理工学院联合研发的条斑紫菜“苏通1号”与“苏通2号”分别成为我国第一、第二个经国家审定批准的条斑紫菜新品种。在相同栽培条件下,与普通条斑紫菜品种相比,新品种不仅产量提高15.0%以上,而且藻体制品品质优良,销售价格比市场平均价高出10.0%[11,12]。
近年来,韩国利用选择性育种方法培育出6个具有优良性状且已广泛应用于生产的条斑紫菜新品种[39]。日本目前也有13个条斑紫菜新品种在其国家农业部注册认证,虽然新品种的来源、育种方式等未知,但大多数已应用于实际生产中[39]。条斑紫菜新品种的推广应用使得紫菜产业摆脱对野生种质的过度依赖,从根本上促进条斑紫菜产业稳定、健康发展(表1)。
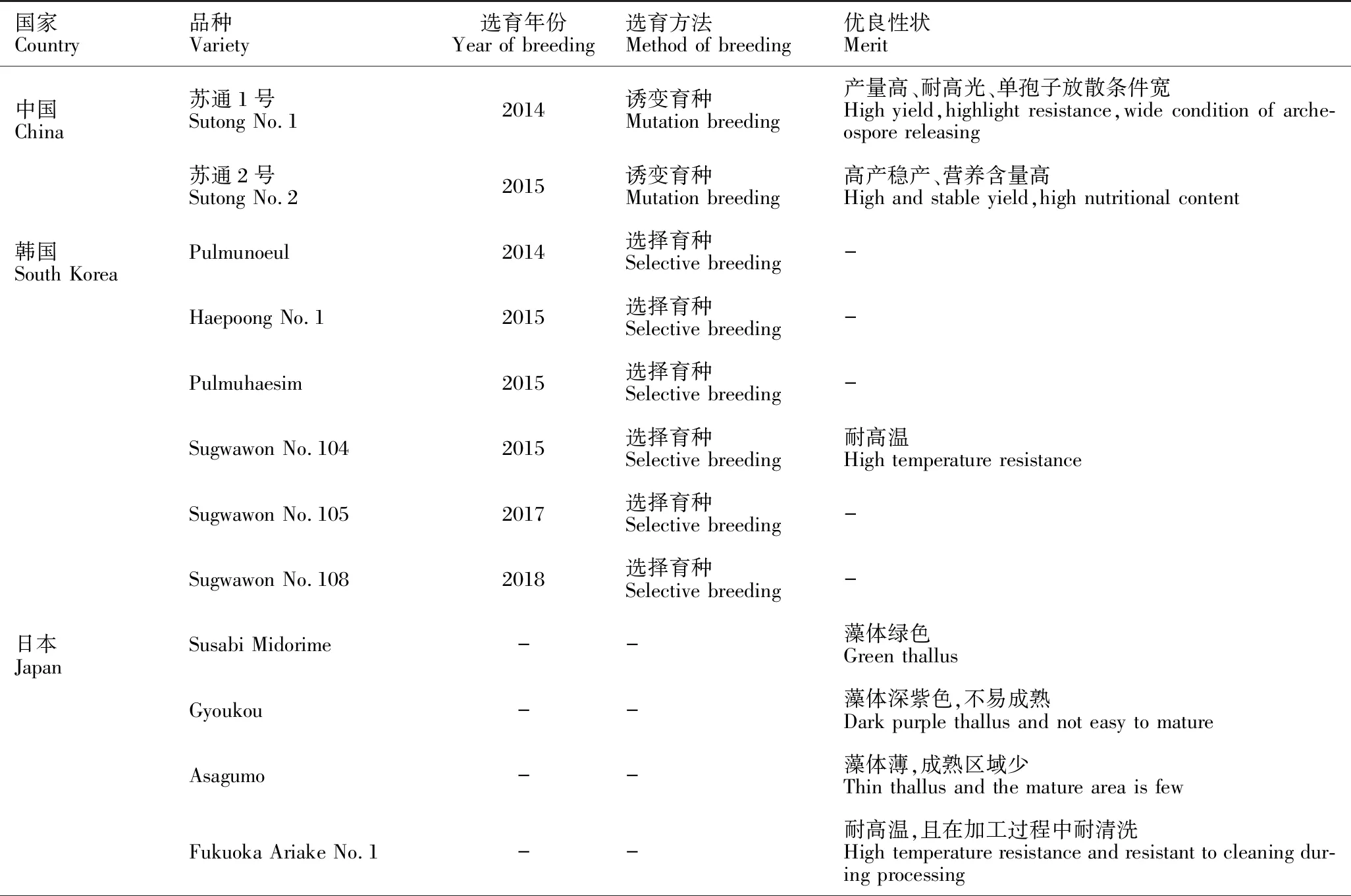
表1 中国、日本和韩国已认证的条斑紫菜新品种
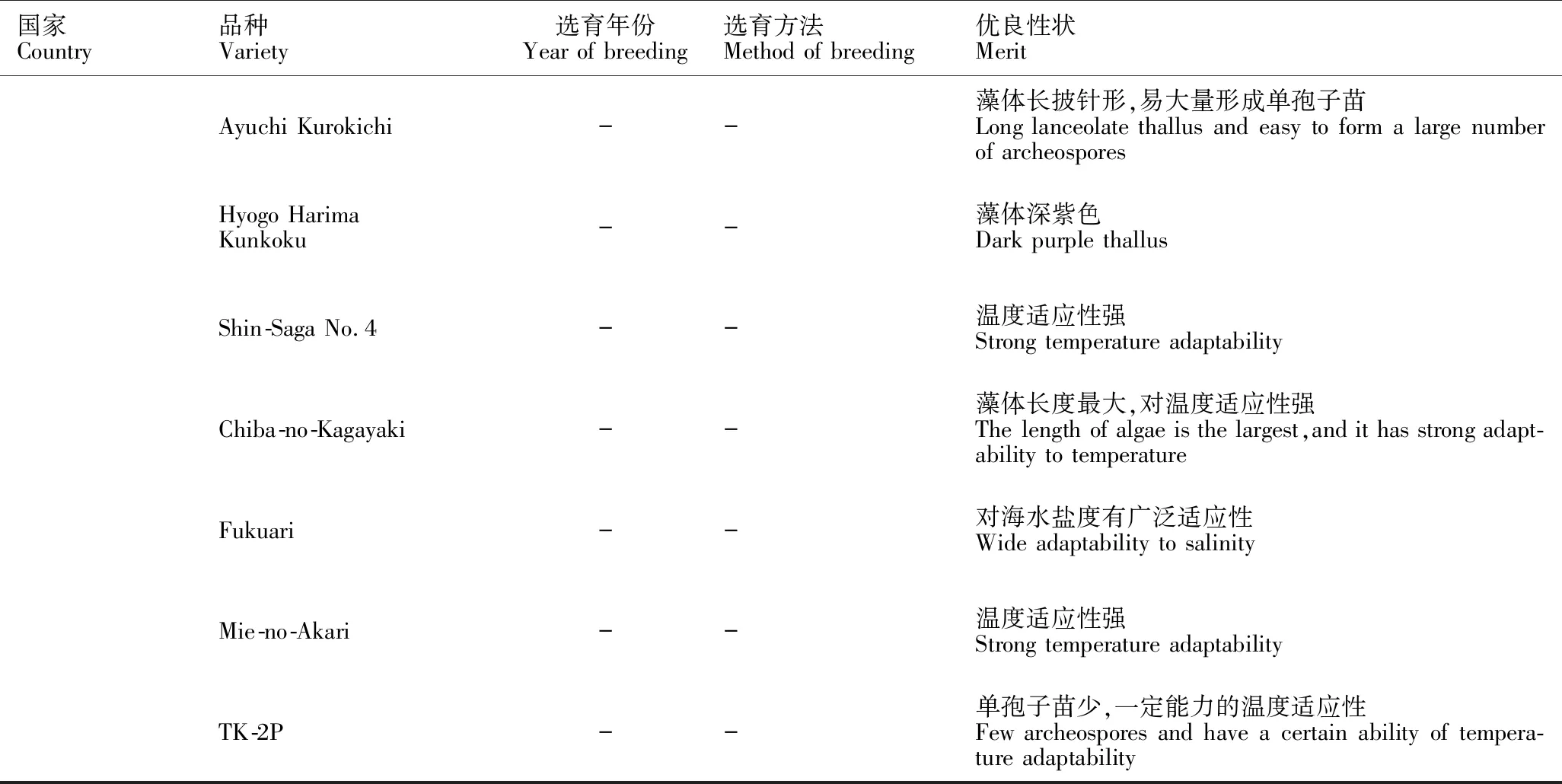
续表1
5 展望
5.1 育种技术有待创新
在条斑紫菜种质创新和应用的实践中,一般需要将多个选育手段结合起来。如对野生条斑紫菜丝状体进行γ-射线和强光胁迫处理,经过连续4代的群体选育,筛选出生长速度快、色泽好及抗高光胁迫的“苏通2号”[12];将条斑紫菜经过γ-射线和高温胁迫处理,最终筛选出抗高温的条斑紫菜新品种“Sugwawon No.104”(韩国)[21]。现有研究基本上是先诱变育种和抗逆性胁迫处理,再经过海区筛选,最终获得性状优良且产量稳定的新品种,这是目前能较快获得条斑紫菜新品种的选育方法。与其他作物相比,紫菜的育种技术还处于较初级阶段,下一步可以考虑从基因水平出发,利用先进的测序技术及分子生物学手段,进行紫菜基因型与表型的筛选,缩短育种周期,提高育种的精准性。
5.2 种质混杂问题
随着条斑紫菜产业发展与主产区北移,即由江苏南通沿岸海域北移至江苏北部(连云港)、山东威海以及渤海湾等沿岸海域[40],产业上应用的种质混杂情况日趋严重。种质来源模糊、种质特性不清楚,对生产上的育苗效率和栽培效果影响较大[41]。建立不同品系单独的栽培系统,对不同品系进行性状评价并掌握其种质特性是育种工作的重要部分。在此基础上,根据不同海区的特点合理应用种质才能达到较好的应用效果。
5.3 种质适应性
适应性是紫菜栽培良种选育的重要指标之一。无论采取何种育种方法,所获得的良种往往存在难育的瓶颈问题,只有通过海区生产中试,研究出不同种质所配套的育苗和栽培技术,才能实现良种大规模推广应用。
条斑紫菜新品种的推广,促进了中国、日本和韩国紫菜产业的健康发展,产生巨大的经济效益和社会效益。但与整个产业发展规模相比,条斑紫菜的新品种良种性状单一,特别是高值化优良性状的栽培品种尚未形成,难以满足产业规模化、多样性的栽培和生产要求。下一步需加大育种创新和应用的研究力度,在育种技术研究、特色性状种质开发、良种性状评估技术等方面进一步突破,以提高我国条斑紫菜优异种质的数量和质量。
猜你喜欢
孩子(2021年1期)2021-02-02
食品与发酵工业(2020年23期)2020-12-22
蔬菜(2020年7期)2020-12-16
幽默大师(2019年10期)2019-10-17
生命科学研究(2018年1期)2018-05-29
婚姻与家庭·婚姻情感版(2018年4期)2018-04-09
农学学报(2018年2期)2018-03-03
安徽医科大学学报(2016年12期)2017-01-15
生态毒理学报(2016年4期)2016-12-02
天津医科大学学报(2015年2期)2015-12-22
