
镉胁迫对不同叶用莴苣品种生长及生理特性的影响
2021-05-26 19:25康育鑫廖水兰兰婕王涛黄语燕陈永快
江苏农业科学 2021年7期
关键词:生理特性
康育鑫 廖水兰 兰婕 王涛 黄语燕 陈永快

摘要:为了鉴选出对镉有较强耐受性的叶用莴苣品种,为镉毒害植物原理提供理论依据,以6种叶用莴苣品种四季、绿神、绿雅、小叶苦苣、紫雅、紫艳为供试材料,在盆栽基质试验条件下,用含有50 mg/L CdCl2·2.5H2O的营养液浇灌进行胁迫处理,探讨了在镉胁迫下不同品种叶用莴苣生理特性的变化情况。结果发现,在一定浓度Cd2+(50 mg/L)胁迫下,对镉耐受程度不同的叶用莴苣品种,在其生物量、叶绿素含量、抗氧化酶活性、丙二醛(MDA)含量、脯氨酸(Pro)含量等生理特性方面产生明显差异。在生物量方面,绿神表现最优;在叶绿素和可溶性蛋白方面,紫艳表现最佳,紫雅、绿神次之;在脯氨酸含量、POD活性方面,绿雅表现最好;在SOD、CAT活性方面,紫雅表现最好;在MDA含量方面,绿神含量最少,紫雅次之,紫艳含量最多。得出结论,绿神、绿雅、紫雅叶用莴苣品种对重金属镉的抗性高于其他3种叶用莴苣品种。
关键词:叶用莴苣;镉胁迫;生理特性
中图分类号:S636.201 文献标志码: A
文章编号:1002-1302(2021)07-0149-06
收稿日期:2020-09-17
基金项目:福建省农业科学院一般项目(编号:A2018-5、Sfjd1941)。
作者简介:康育鑫(1987—),男,福建泉州人,硕士,主要从事设施农业、数字农业研究。E-mail:kangyuxin369@163.com。
随着现代工业的快速发展,重金属在土壤中广泛分布,造成大面积土壤污染,并严重危害人体健康[1-2]。土壤污染可以诱导形成许多活性氧自由基,引起细胞膜脂不同程度过氧化及蛋白质等生物大分子物质变性,进而严重侵害植物生长发育[3]。镉(镉)作为毒性最强、危害较大、污染最普遍的重金属之一,不但能影响植物正常生长发育,而且还能经过营养链方式损害生命健康[4-6]。大量研究证实,重金属镉毒害首要引起植物体内的抗氧化系统失衡,增加细胞内活性氧含量,从而造成植物细胞受损甚至死亡[7]。研究表明,重金属胁迫不仅影响植株生长及生理特性[8],还可间接反映环境中污染物的存在。
葉用莴苣别称生菜,原产地在欧洲,我国很多地区均有栽培,富含花青素、膳食纤维、类胡萝卜素等多种有益成分[9],具有降血压、降胆固醇、预防贫血及心律紊乱等多种保健功能[10-13]。目前国内外对叶用莴苣本身在镉胁迫下各生理指标的变化及镉胁迫对不同品种叶用莴苣生理生化指标影响的差异研究较少。综合预试验及相关研究成果,本试验选择中等镉污染浓度(50 mg/L Cd2+)进行镉胁迫处理,研究6个叶用莴苣品种的生物量、叶绿素含量、抗氧化酶活性、MDA含量、Pro含量等生理特性方面的差异性,从而鉴选对镉有较强耐受性的品种,为更好地了解重金属毒害植物原理提供一定理论支撑。
1 材料与方法
1.1 供试材料
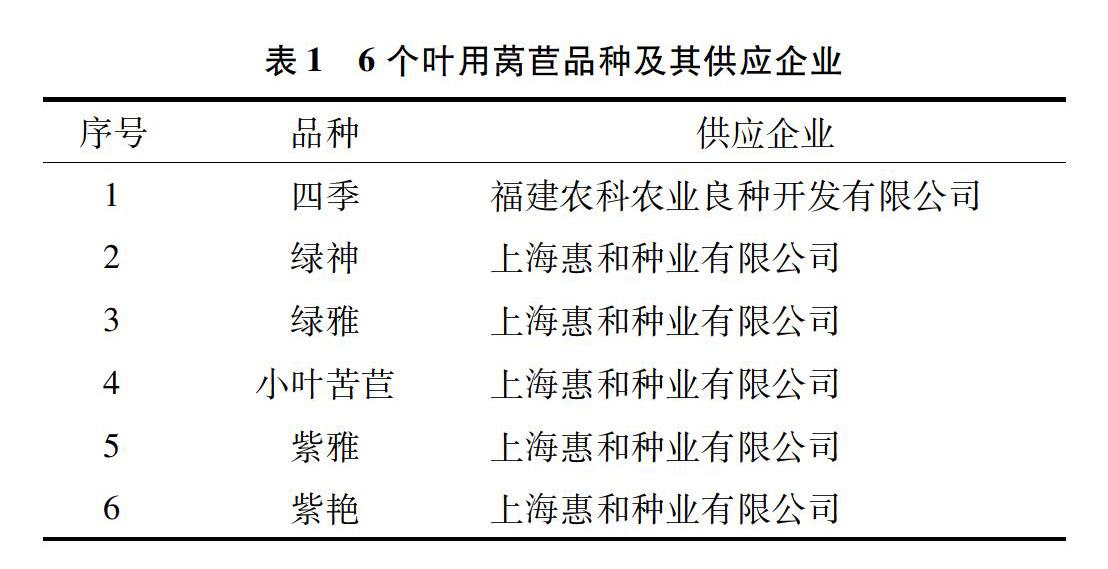
以6个叶用莴苣品种四季、绿神、绿雅、小叶苦苣、紫雅、紫艳为试验材料,详见表1。
1.2 试验设计
试验在福建省农业科学院海峡农业现代示范园薄膜温室和玻璃温室生理生化实验室进行。
1.2.1 育苗 2018年10月30日开始,利用自动播种机播种,选取72孔规格穴盘和丹麦进口草炭基质来育苗,并用蛭石洒在基质上面同时浇透水,后期做好水肥管理工作。
1.2.2 盆栽试验 盆栽试验于12月10日在海峡农业现代农业示范园薄膜温室进行。将供试土壤(草炭 ∶珍珠岩体积比=2 ∶1),装入聚乙烯塑料盆(15 cm×13 cm)。当叶用莴苣幼苗长至4~5张真叶,根系底部形成根坨时,开始在聚乙烯塑料盆中定植,每盆定植1株。用正常营养液预培养7 d后,镉胁迫处理使用含有50 mg/L CdCl2·2.5H2O的营养液浇灌,非镉胁迫处理使用正常营养液浇灌(对照),每天浇灌1次,每个处理3个重复。正常营养液配方A:添加Ca(NO3)2 80 g/L,KNO3 30 g/L;配方B:KH2PO4 15 g/L,MgSO4 50 g/L,EDTA-Fe 2.3 g/L,MnSO4 0.6 g/L,ZnSO4 0.3 g/L,CuSO4 0.02 g/L,NH4 Mo 7.5 mg/L,H3BO3 0.3 g/L。生长35 d后收集全部植株,测定各项生理生化指标。
1.3 测定指标及方法
1.3.1 生物量测定 根据植株地上部和地下部鲜质量测量生物量。选取长势匀称的植株,用自来水反复冲洗,最后用去离子水洗净,用滤纸吸干多余水分,记录植株地上部和地下部鲜质量。测量数据选用平均值。
1.3.2 生理生化指标测定 叶绿素采用丙酮-乙醇混合液提取测定[14];可溶性蛋白含量测定采取考马斯亮蓝法[15];脯氨酸(Pro)含量测定采取磺基水杨酸法[16];丙二醛(MDA)含量测定采取硫代巴比妥酸法[17];超氧化物歧化酶(SOD)活性测定采取氮蓝四唑(NBT)法[18];过氧化物酶(POD)活性测定采取愈创木酚法[19];过氧化氢酶(CAT)活性测定采取紫外吸收比色法[20]。测量数据选用平均值。
1.4 数据处理
数据采取 Excel、DPS软件进行统计和分析。
2 结果与分析
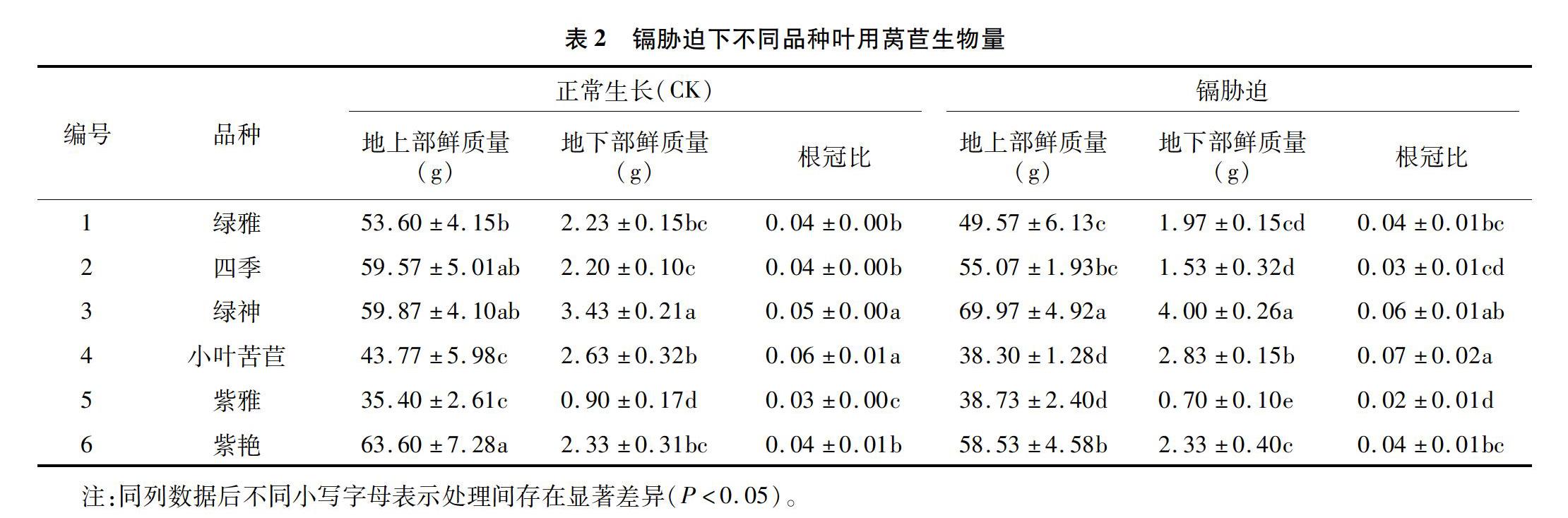
2.1 镉胁迫下叶用莴苣采收后生物量情况
采用50 mg/L Cd2+处理不同种类叶用莴苣,采收后植株地上部、地下部鲜质量及根冠比均产生较明显的差异(表2)。地上部鲜质量表现为,正常生长(CK)以紫艳最高,达63.60 g,紫雅最低,仅为35.40 g;镉胁迫下绿神最高,为69.97 g,比正常生长(CK)增长了16.87%,小叶苦苣最低,为 38.30 g。镉对绿雅、四季、小叶苦苣、紫艳生长产生一定的抑制作用,生物量明显降低;而绿神、紫雅几乎不受镉影响。地下部鲜质量表现为,正常生长(CK)以绿神根系生长最为旺盛,高达3.43 g,而紫雅质量最低,仅有0.90 g;镉胁迫下绿神最高,为4.00 g,比正常生长(CK)增长了16.62%,紫雅最低,为0.70 g。关于地下部生长,镉对绿雅、四季具有一定的抑制作用,而绿神、小叶苦苣、紫艳几乎不受镉影响。在根冠比方面,除了四季、紫雅受到镉影响,其他品种叶用莴苣几乎不受影响。
2.2 镉胁迫对不同品种叶用莴苣叶绿素含量的影响
叶绿素水平能够衡量光合作用强弱,也可以揭示在胁迫下植物被侵害的情况。由图1可知,在 50 mg/L Cd2+胁迫下,除了紫艳,其他叶绿素含量均明显下降;四季下降最多,达40.02%,绿雅下降23.53%,小叶苦苣下降22.57%,绿神下降10.87%,紫雅下降最少,仅8.43%。绿雅、四季、紫雅叶绿素含量之间差异不显著,绿神、小叶苦苣之间差异也不显著。Cd2+被叶用莴苣吸收后,能够抑制合成叶绿素中几种酶的活性,阻碍叶绿素合成,从而进一步造成了植物光合作用低效,对叶用莴苣生理生化代谢产生巨大的毒害作用。综上所述,绿神、紫雅叶绿素含量降低得最为缓和,而紫艳在镉胁迫下合成更多叶绿素,提高光合作用效率,更能抵抗镉毒害。
2.3 镉胁迫对不同品种叶用莴苣可溶性蛋白含量的影响
可溶性蛋白影响植物耐受性,其含量增多会使植物适应性增强。由图2可知,在50 mg/L Cd2+胁迫下,所有叶用莴苣品种的可溶性蛋白含量都有不同幅度的提高,表现为一定刺激作用。其中紫艳含量最高,为14.79 mg/g,与绿神差异不显著,与其他品种之间存在显著性差异。而与对照相比,紫艳提高了2.51倍,紫雅提高了2.00倍,绿神提高了1.31倍,小叶苦苣提高了64.80%,四季提高了53.84%,绿雅提高了46.50%。结果表明,50 mg/L浓度Cd2+会导致不同品种叶用莴苣的可溶性蛋白含量提高,诱导镉结合蛋白降低镉的毒性,从而抵抗镉毒害。其中紫艳对镉的适应性最强,绿神、紫雅次之。
2.4 镉胁迫对不同品种叶用莴苣脯氨酸含量的影响
脯氨酸能起到对细胞质渗透平衡的调节作用。在胁迫条件下,植物体内脯氨酸含量大量积累,可以降低重金属毒害,从而提高植物体的抗逆能力。
由图3可知,在50 mg/L Cd2+胁迫下,绿雅脯氨酸含量最高,为12.20 μg/g,比对照提高48.80%;紫雅次之,为11.11 μg/g,比对照下降了3.56%;其他品种叶用莴苣脯氨酸差异不显著。正常生长时,紫艳脯氨酸含量最多,高达22.36 μg/g;而镉胁迫下,其脯氨酸含量下降幅度最大,为51.75%。结果表明,绿雅抗逆性最强,而紫艳最容易受镉伤害。
2.5 镉胁迫对不同叶用莴苣品种丙二醛含量的影响
丙二醛(MDA)在一定程度上反映了膜脂过氧化情况,从而可以明晰膜系统受伤害的情况。植物体内MDA积累的量越多,膜脂过氧化的程度就越大,细胞膜受到的伤害就越严重。由图4可知,无论是正常生长还是受到50 mg/L Cd2+胁迫,紫艳的MDA含量都是最高,紫雅次之,与其他品种之间存在显著差异。镉胁迫下,紫艳MDA含量分别是紫雅、小叶苦苣、四季、绿雅、绿神的1.61倍、3.71倍、4.02倍、4.19倍、4.37倍。与对照相比,四季MDA含量增加幅度最大,达91.49%;绿雅、小叶苦苣、紫艳、紫雅为84.62%、47.97%、47.82%、14.11%;绿神最低,为11.85%。结果表明,四季、绿雅最容易发生細胞膜脂过氧化,在镉胁迫下其细胞膜受到严重伤害,而绿神、紫雅MDA增加量远远低于其他品种,耐受性较强。
2.6 镉胁迫对不同叶用莴苣品种抗氧化酶活性的影响
当受到镉胁迫时,植物体内形成大量活性氧,而抗氧化酶系统能通过消灭活性氧来减少损害植物。抗氧化酶系统主要由SOD、POD、CAT构成,植物体内形成的自由基在SOD作用下生成H2O2,而H2O2在POD和CAT作用下进一步催化分解为无毒无害的H2O和O2。由图5、图6、图7可知,50 mg/L Cd2+胁迫下,绿雅的SOD、POD、CAT都发生不同幅度的提高。在SOD活性方面,绿雅、四季、紫雅表现为上升,较对照分别增长了3.90%、9.80%、14.87%;绿神、小叶苦苣、紫艳表现为下降。在POD活性方面,与对照相比,绿雅提高最多,为84.62%;绿神次之,为42.86%;小叶苦苣为5.36%;而紫艳活性下降幅度最大,为50.00%;四季、紫雅分别下降了8.33%、5.26%。在CAT活性方面,除了小叶苦苣,其他品种在重金属镉胁迫下的CAT活性都有不同程度的提高。紫雅增幅最大,为66.00%;绿雅、紫艳、绿神分别增加57.89%、53.23%、18.18%;四季增幅最小,为15.19%。结果表明, 在镉胁迫下,紫雅、绿雅可以通过发挥抗氧化酶系统作用,从而减少镉侵害自身。
3 讨论
3.1 镉胁迫下不同叶用莴苣采收后生物量情况
生物量测定方面,以绿神、紫艳表现俱佳,抵御镉胁迫能力较强,这与贾茵等的研究结果[21-22]相仿。生物量受影响,主要是由于与光合作用、呼吸作用相关的酶会失去活性,甚至出现变性的现象,从而使合成相关酶的活性降低,导致植物的光合作用、呼吸作用的速率呈现下降趋势,使得植物体无法正常生长发育[23]。本试验结果说明了不同品种叶用莴苣耐镉性和生物量幅度的变化均有明显的差别。
3.2 镉胁迫下不同品种叶用莴苣生理生化指标的影响
叶绿素总量高低决定了光合作用的强弱。在50 mg/L Cd2+胁迫下,除了紫艳品种,其他叶用莴苣品种叶绿素含量均明显下降,绿雅、四季、紫雅之间差异不显著,绿神、小叶苦苣之间差异也不显著,这与牟祚民等的研究结果[24]类似,也与刘燕等研究得到的镉胁迫下油菜叶绿素总量有下降趋势的结论[25]相仿。分析其原因,可能是一定浓度Cd2+被不同品种叶用莴苣吸收后,能够通过降低酶活性来降低植物光合作用效率,并影响叶用莴苣生理生化代谢。本试验中,紫艳可以合成更多叶绿素,提高光合作用效率,更能抵抗镉毒害。
可溶性蛋白含量可以反映植物的抗逆性强弱。本试验中,紫艳可溶性蛋白含量最高,增长最快,与绿神差异不显著,与其他叶用莴苣品种之间存在显著差异,该研究结果与刘文英等研究[26]类似。可能的原因是一定浓度Cd2+进入不同品种叶用莴苣后,提高可溶性蛋白含量,刺激蛋白结合镉而降低镉的毒性,因而能抵抗镉毒害。其中紫艳对镉的适应性最强,绿神、紫雅次之。
脯氨酸在一定程度上能够衡量植物抗逆性,脯氨酸含量越高抗性越强[27]。脯氨酸含量的增加可以降低重金属对机体的毒害,从而提高植物体的抗逆能力。本试验结果表明,绿雅抗逆性最强,而紫艳容易受镉影响,这与王阳阳等的研究[28]相仿。原因可能是,在50 mg/L Cd2+胁迫下,绿雅能显著增加脯氨酸含量,具有较强的活性氧清除能力,减少膜脂化程度,抵御镉的毒害。
丙二醛含量是植物膜脂过氧化的一个重要指标,积累量较多将严重伤害细胞膜。本试验结果表明,所有叶用莴苣品种的细胞膜均受到一定伤害。四季、绿雅更易发生细胞膜脂过氧化,细胞膜受到严重伤害;而绿神、紫雅MDA的增加量远远低于其他品种,耐受性较强,该研究结果与刘文英等的研究[26,29]相似。主要是因为MDA是镉胁迫膜脂过氧化产物之一,能反映膜脂过氧化的程度[30-32]。
植物體内进化形成的抗氧化酶系统可以在一定程度上清除植物在逆境胁迫下细胞内产生的活性氧,从而可以抵抗逆境胁迫毒害作用[33]。本试验中,绿雅可以通过提高SOD、POD、CAT抗氧化酶活性来清除活性氧,从而减轻镉对自身伤害,而其他品种也可以通过提高其中某个抗氧化酶活性来提升整体抗逆性。这是因为抗氧化保护酶系统主要有SOD、POD、CAT,可以很好地控制活性氧代谢活动。这一结论与Shah等的研究的结果[32,34]相似。SOD、POD活性增加,证明SOD、POD共同作用消灭自由基与过氧化物,提升了抗氧化水平,或许是通过调整消灭系统来顺应活性氧增加,以减少伤害植物。绿雅、紫雅的SOD、CAT活性增长率较其他品种更大,表明绿雅、紫雅抗逆性更强。该结果和郑世英等的研究结论[35-36]类似。除了小叶苦苣,其他品种叶中的CAT活性均有所增加,而绿雅、紫雅品种CAT活性增加幅度最大,证实过氧化氢酶可以消灭自由基;而小叶苦苣CAT活性降低,Shah等表示其与酶活性降低和活性氧较多等密切相关[32]。本试验中,50 mg/L Cd2+胁迫下,绿雅、紫雅膜脂过氧化程度相对较低,其抗氧化酶的清除能力强于其他品种。
4 结论
镉含量会影响植物代谢活动和生长发育[37]。本研究表明,在一定浓度Cd2+(50 mg/L)胁迫下,6种叶用莴苣品种对镉的耐受能力存在一定差异,综合表现为绿神、绿雅、紫雅对镉的耐受性高于其他3种叶用莴苣品种,可为重金属毒害植物原理提供一定理论支撑。
参考文献:
[1]Doyurnm S,Celik A C. Removal from aqueous solutions by olive cake[J]. Journal of Hazardous Materials,2006,138(1):22-28.
[2]王 维. 水稻CdCl2吸收的区域模型及其调控研究[D]. 南京:南京林业大学,2012.
[3]江行玉,赵可夫. 植物重金属伤害及其抗性机理[J]. 应用与环境生物学报,2001,7(1):92-99.
[4]刘 莉,钱琼秋. 影响作物对镉吸收的因素分析及土壤镉污染的防治对策[J]. 浙江农业学报,2005,17(2):111-116.
[5]Pelfrêne A,Waterlot C,Mazzuca M,et al.Assessing Cd,Pb,Zn human bioaccessibility in smelter-contaminated agricultural toposils(northern France)[J]. Environmental Geochemistry and Health,2011,33(5):477-493.
[6]陈 会,任艳芳,陈秀兰,等. Cd胁迫下不同耐性水稻植株幼苗生长和抗氧化酶的变化[J]. 江西农业大学学报,2012,34(6):1099-1104.
[7]尹永强,胡建斌,邓明军. 植物叶片抗氧化系统及其对逆境胁迫的响应研究进展[J]. 中国农学通报,2007,23(1):105-110.
[8]王启明. 重金属Cr6+胁迫对玉米幼苗生理生化特性的影响[J]. 河南农业科学,2006,35(8):37-40.
[9]刘 蕊,王晓茜,韩莹琰,等. 北京地区春茬栽培的结球生菜品种筛选[J]. 北京农学院学报,2018,33(2):53-57.
[10]Heimler D,Vignolini P,Arfaioli P,et al. Conventional,organic and biodynamic farming:differences in polyphenol content and antioxidant activity of Batavia lettuce[J]. Journal of the Science of Food and Agriculture,2012,92(3):551-556.
[11]Pérez-López U,Miranda-Apodaca J,Muoz-Rueda A,et al. Lettuce production and antioxidant capacity are differentially modified by salt stress and light intensity under ambient and elevated CO2[J]. Journal of Plant Physiology,2013,170(17):1517-1525.
[12]Petroni K,Pilu R,Tonelli C. Anthocyanins in corn:a wealth of genes for human health[J]. Planta,2014,240(5):901-911.
[13]Zhang Y,Butelli E,Martin C. Engineering anthocyanin biosynthesis in plants[J]. Current Opinion in Plant Biology,2014,19:81-90.
[14]高俊凤. 植物生理学试验指导[M]. 北京:高等教育出版社,2006:142-143.
[15]焦 洁. 考马斯亮蓝 G-250 染色法测定苜蓿中可溶性蛋白含量[J]. 农业工程技术,2016,36(17):33-34.
[16]张志良,瞿伟菁,李小方. 植物生理学试验指导[M]. 北京:高等教育出版社,2009.
[17]赵世杰,许长成,邹 琦,等. 植物组织中丙二醛测定方法的改进[J]. 植物生理学通讯,1994,30(3):207-210.
[18]沈文飚,徐朗莱,叶茂炳,等. 氮蓝四唑光化还原法测定超氧化物歧化酶活性的适宜条件[J]. 南京农业大学学报,1996,19(2):101-102.
[19]李忠光,龚 明. 愈创木酚法测定植物过氧化物酶活性的改进[J]. 植物生理学通讯,2008,44(2):323-324.
[20]李 玲. 植物生理学模块试验指导[M]. 北京:科学出版社,2009.
[21]贾 茵,刘才磊,兰晓悦,等. Cd胁迫对小报春幼苗生长及生理特性的影响[J]. 西北植物学报,2020,40(3):454-462.
[22]白瑞琴,孟海波,周 爽. 重金属Cd对两个马铃薯品種生长发育的影响[J]. 华北农学报,2012,27(1):168-172.
[23]王焕校. 污染生态学基础[M]. 昆明:云南大学出版社,1990:71-148.
[24]牟祚民,姜贝贝,潘远智,等. 重金属胁迫对天竺葵生长及生理特性的影响[J]. 草业科学,2019,36(2):434-441.
[25]刘 燕,彭钟山,蒋光霞. 重金属胁迫对甘蓝型油菜叶绿素的影响[J]. 贵阳学院学报(自然科学版),2007,2(1):46-49.
[26]刘文英,张东旭,张永芳,等. 不同浓度Cd胁迫对3种芥菜型油菜幼苗生理特性的影响[J]. 山西大同大学学报(自然科学版),2014,30(4):45-48.
[27]戴高兴,彭克勤,萧浪涛,等. 聚乙二醇模拟干旱对耐低钾水稻幼苗丙二醛、脯氨酸含量和超氧化物歧化酶活性的影响[J]. 中国水稻科学,2006,20(5):557-559.
[28]王阳阳,任艳芳,周国强,等. Cd胁迫对不同抗性水稻品种幼苗生长和生理特性的影响[J]. 中国农学通报,2009,25(24):450-454.
[29]韩承华. Cd、Pb、Cu、Zn对三种水生叶菜生长的影响以及Se缓解效果的研究[D]. 扬州:扬州大学,2017.
[30]Sanità di Toppi L,Gabbrielli R. Response to cadmium in higher plants[J]. Environmental and Experimental Botany,1999,41(2):105-130.
[31]邵国胜,Hassan M J,章秀福,等. Cd胁迫对不同水稻基因型植株生长和抗氧化酶系统的影响[J]. 中国水稻科学,2004,18(3):239-244.
[32]Shah K,Kumar R G, Verma S,et al. Effect of cadmium on lipid peroxidation,superoxide anion generation and activities of antioxidant enzymes in growing rice seedlings[J]. Plant Science,2001,161(6):1135-1144.
[33]徐文君,程江峰. 土壤邻苯二甲酸二丁酯对白菜生长和抗氧化酶系统的影响[J]. 江苏农业科学,2020,48(7):142-147.
[34]肖美秀,林文雄,陈冬梅,等. Cd胁迫对耐性不同的水稻幼苗膜脂过氧化和保护酶活性的影响[J]. 中国生态农业学报,2006,14(4):256-258.
[35]郑世英,王丽燕,张海英. Cd胁迫对两个大豆品种抗氧化酶活性及丙二醛含量的影响[J]. 江苏农业科学,2007(5):53-55.
[36]Wu F B,Zhang G P,Dominy P. Four barley genotypes respond differently to cadmium:lipid peroxidation and activities of antioxidant capacity[J]. Environmental and Experimental Botany,2003,50(1):67-78.
[37]李 晔,李玉双,孙丽娜,等. 重金属Cd胁迫对不同玉米品种生理生化指标的影响[J]. 安徽农业科学,2011,39(5):2627-2628,2657.
猜你喜欢
云南中医中药杂志(2017年1期)2017-02-17
山东农业科学(2016年12期)2017-01-21
现代园艺(2016年9期)2016-07-16
现代园艺(2016年2期)2016-03-15
