
不同林龄马尾松人工林土壤酶活性及其生态化学计量特征
2021-06-05 01:23梁月明潘复静靳振江马姜明杨章旗零天旺李明金
桂林理工大学学报 2021年1期
李 臻, 梁月明, 潘复静, 靳振江, 马姜明, 杨章旗, 零天旺, 李明金
(1.桂林理工大学 a.环境科学与工程学院;b.广西环境污染控制理论与技术重点实验室, 广西 桂林 541006; 2.中国地质科学院岩溶地质研究所/自然资源部、广西壮族自治区岩溶动力学重点实验室, 广西 桂林 541004; 3.珍稀濒危动植物生态与环境保护教育部重点实验室, 广西 桂林 541004; 4.广西壮族自治区林业科学研究院, 南宁 530002; 5.广西壮族自治区横县镇龙林场, 南宁 530300)
0 引 言
我国现有的人工林已经占据大多数宜林造林的优越地段, 但是人工林长期进行纯林经营, 会导致其层次结构简单、生态系统脆弱[1]。其中, 马尾松人工林占据人工林很大的比例,马尾松(Pinusmassoniana)具有耐干旱、耐瘠薄、喜光等特点, 是我国松类中分布最广、数量最多的造纸材树种。在广西, 马尾松大面积种植, 且马尾松人工林是广西主要的三大用材林(松、杉、桉)之一, 然而马尾松人工林普遍存在林分结构不合理、生产力低、生态功能差、物种单一等问题[2-3]。尽管广西马尾松人工林具有较高的碳汇功能, 但是其生长依然受到很大的局限性, 如养分限制(N、P等)、经营管理效率低等[4-5]。
土壤养分是影响森林生态系统生产力、结构和功能的重要因素, 充足且均衡的土壤养分供应是植物稳定快速生长的需要[6]。目前, 作为森林资源关键组分之一的人工林, 其发展过程中遇到的土壤质量与生产力等问题受到了普遍关注[7-8]。土壤养分库大小可以反映土壤的资源水平, 但对植被生产力起关键作用的是可利用性养分, 即土壤中易被植物吸收同化的养分元素或化合物的数量[9-11]。氮和磷是对植物生长影响最大的矿质元素, 影响着森林地上和地下的生产力及碳积累[12-13]。人工林养分循环一般具有低归还、低循环速率的特点, 这些问题都严重限制了林木的健康和快速生长[14-16]。土壤酶的催化作用对土壤中元素(包括C、N、P、S)循环与迁移有着重要影响, 其活性也可以作为指示土壤肥力的指标[17]。杨涛等[18]的研究表明樟子松人工林的土壤养分与酶活性之间存在密切关系, 当土壤地力衰退、养分含量降低时, 酶活性也随之降低; 李志辉等[19]的研究表明桉树人工林的土壤氧化氢酶和脲酶有利于土壤养分的转化, 增强其活性有利于林地养分可利用性的提高, 促进林木生长; 陈彩虹等[20]对杉木、枫香、樟树和湿地松等4种人工林的研究表明土壤中过氧化氢酶、脲酶、磷酸酶、脱氢酶和纤维素酶的活性与土壤养分因子间均存在极显著或显著的正相关关系。以上结果表明, 人工林土壤酶活性与土壤养分之间存在着紧密的联系。
生态化学计量学是近年来新兴的一个生态学研究领域, 集中探讨了C、N、P三种主要元素的化学计量关系, 是一种研究生态系统中的多种元素间平衡协调情况的新方法[21]。土壤酶生态化学计量是指微生物获取C、N、P元素后所分泌的酶的活性的比值, 它是生态计量理论和生态代谢理论的交叉点。已知,β-葡萄糖苷酶(β-Glucosidase,βG)与碳循环相关;β-乙酰葡糖胺糖苷酶(β-N-acetylglucosaminidase, NAG)与氮循环相关; 土壤酸性磷酸酶(Acid phosphatase, ACP)、酸性磷酸酶(Alkaline phosphatase, ALP)与磷循环相关[22]。以βG∶(NAG+LAP)作为酶活性C∶N值、βG∶AP作为酶活性C∶P值、(NAG+LAP)∶AP作为酶活性N∶P值[23], 来反映微生物对养分需求差别。 微生物可以通过调节土壤养分的循环过程, 对植物土壤的养分协调产生重要影响,且其综合了生物学、化学、物理计量学等基本原理, 可以更深入地探究生物之间的养分循环内在机理, 因此土壤酶活性生态化学计量特征可以指示土壤中养分的可利用情况以及养分受限情况[23-25]。已有的研究发现土壤酶活性生态化学计量值会受到人工林类型、林下植被类型、林分和土壤理化性质等因素的影响[22], 但是基于马尾松人工林的土壤酶活性生态化学计量学理论来探讨不同林龄、不同土壤深度的土壤酶活性与土壤养分之间关系的研究仍然较少, 该理论可以深入探究马尾松人工林的养分限制及循环情况, 进而为本地区马尾松人工林的经营提供更加科学的建议, 最终达到兼顾其经济效益和生态效益的目的。
本研究基于长期定位试验, 选择马尾松人工林的4种龄林作为研究对象, 收集了0~10、10~20和20~30 cm的土壤样品, 详细分析了土壤β-葡萄糖苷酶(βG)、β-乙酰葡糖胺糖苷酶(NAG)、酸性磷酸酶(ACP)和碱性磷酸酶(ALP)酶活性与土壤的C、N和P的含量之间的关系, 主要目的在于:(1)摸清不同林龄马尾松人工林在不同土壤深度下土壤酶活性和养含量的变化; (2)计算土壤酶活性生态化学计量比值, 分析马尾松人工林的养分限制情况。通过以上研究, 进一步提高对马尾松人工林的养分限制及其循环的认识, 为马尾松纯林的经营管理工作提供科学的理论依据。
1 研究区概况
研究区位于广西壮族自治区南宁市横县北部的镇龙林场, 东经108°48′—109°37′, 北纬22°08′—23°30′, 海拔400~700 m, 以低山丘陵地形为主。横县东连贵港市, 南接灵山县、浦北县, 西接青秀区, 北壤宾阳县。横县镇龙林场经营总面积达到6 069.9 hm2, 主要种植树种为巨尾桉(Eucalyptusgrandis)、马尾松和杉木(Cunninghamialanceolata)等, 水资源丰富, 年平均降雨量1 450 mm; 土壤类型主要是酸性或微酸性的赤红壤[48]。
2 研究方法
2.1 样地设置
2018年7月(生长季), 选取4种林龄(分别为6、17、32和58 a)的马尾松人工林作为研究对象, 分别为幼龄林(AF1)、中龄林(AF2)、成熟林(AF3)和过熟林(AF4)。在每个林分中, 选取立地条件、土层厚度、土壤质地和林木长势较一致的区域, 各设立3个20 m×20 m的标准样方, 共设置样方12个。不同林龄马尾松人工林的基本特征见表1。
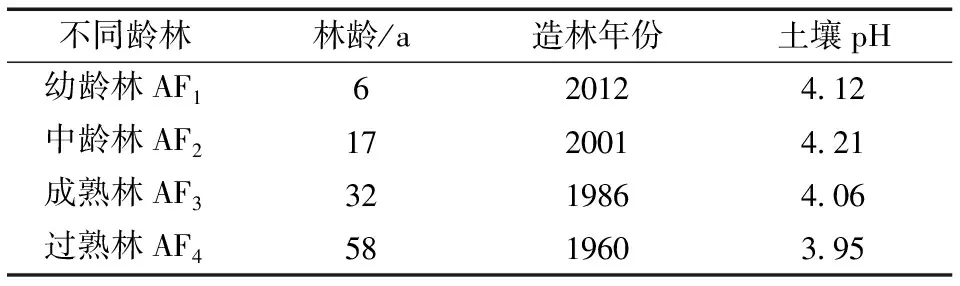
表1 不同龄林马尾松人工林的基本概况Table 1 Basic overview of Pinus massoniana plantationsin different age stands
2.2 土壤样品采集
将每个样方分成4个10 m×10 m的子区域, 在样方中心点和4个子区域的中心点采集不同深度土壤样品(0~10、10~20、20~30 cm), 将5个采样点的同层土样混合均匀(每个样方得到3种深度的混合土样各1份), 去除其中植物残体并过2 mm(10目)筛,然后将其分成两等份, 一份装进塑料封口袋并放入4 ℃冰箱保存, 用于检测土壤酶活性;另一份,风干后一半过0.85 mm筛,一半过0.15 mm筛,分别用于土壤N与C、P的测定。
2.3 样品分析
2.3.1 土壤酶活性的测定β-葡萄糖苷酶(βG)、β-乙酰葡糖胺糖苷酶(NAG)、酸性磷酸酶 ( ACP)、碱性磷酸酶( ALP)活性采用MUB荧光光度法测定。称取新鲜土壤 0.4 g于100 mL灭菌带盖玻璃瓶中, 加入50 mL 灭菌冷却后的醋酸钠(碱性磷酸酶)缓冲液, 用磁力搅拌器将土壤悬液搅拌均匀(边搅拌边加样), 加入到96孔微孔板中, 将微孔板置于黑暗条件下20 ℃培养4 h后, 每个孔加入10 μL 1 mol/L的NaOH溶液使反应终止, 用纸巾擦净酶标板底部(去除凝结物防止影响读数), 用多孔板检测仪或酶标仪测定荧光值, 激发波长365 nm, 发射波长450 nm。经过负控制和淬灭校正后, 土壤酶活性单位用nmol·g-1·h-1[25]。
2.3.2 土壤养分的测定 土壤有机碳(SOC)含量采用KCr2O7+H2SO4氧化法测定; 土壤全氮(TN)含量采用凯氏定氮法并用流动注射仪(FIAstar 5000, FOSS, Hillerod, Denmark)测定; 土壤铵态氮和硝态氮先用2 mol/L KCL溶液浸提,过滤后上清液用AA3流动注射仪(seal,德国)测定。土壤全磷(TP)含量采用加NaOH后放入马弗炉高温消煮, H2SO4+HCl清洗后以钼蓝显色液进行显色, 分光光度计进行测定; 土壤有效磷(AP)用NaHCO3溶液浸提后以钼蓝显色液显色, 最后用分光光度计进行测定。
2.4 统计与分析
用Excel进行数据整合, 土壤养分含量和酶活性大小均为平均值。用SPSS 24.0进行数据处理和统计分析(p<0.05), 利用单因素方差分析(one-way ANOVA)及多重比较分析(LSD)比较马尾松人工林不同林龄和土壤深度的养分含量和酶活性的差异。对土壤酶活性进行对数转换后, 再进行比值计算以表示酶活性的生态化学计量特征:用lnβG∶ln NAG表示土壤酶活性C∶N值; lnβG∶ln(ACP+ALP)表示酶活性C∶P值; ln NAG∶ln(ACP+ALP)表示酶活性N∶P值[23]。利用皮尔森相关分析方法(Pearson correlation analysis)和RDA方法(冗余分析)分析了马尾松人工林土壤酶活性化学计量比值与土壤养分的相关性和其受土壤环境因子影响的大小。
3 结果与分析
3.1 土壤养分的特征
在不同土壤深度中, 马尾松人工林同一龄林的SOC和 TN的含量随着土壤深度的增加而逐渐降低, TP含量变化较小, AP含量在幼龄林、成熟林和过熟林都表现为: 随土壤深度增加而逐渐降低, 在中龄林时则随土壤深度增加先降低后小幅度升高。同一土壤深度下, 不同龄林的SOC、TN和AP含量无显著差异,但是TN含量随着林龄的增长逐渐降低, AP含量在幼龄林时最高; TP含量差异显著,幼龄林最低, 中龄林最高; 土壤氨氮含量没有显著差异(20~30 cm成熟林与过熟林除外), 土壤硝氮含量在20~30 cm土壤层表现出显著差异: 幼龄林>成熟林>过熟林>中龄林(表2)。
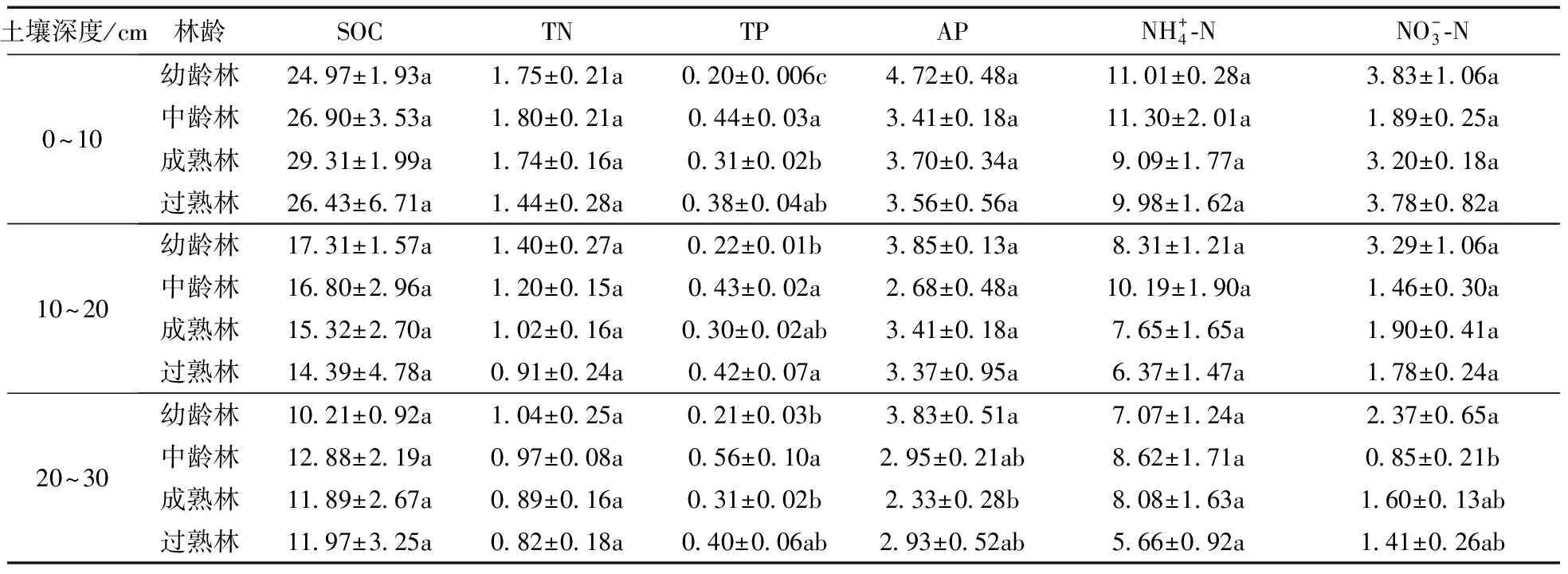
表2 不同龄林马尾松人工林土壤养分差异性Table 2 Differences of soil nutrients in different Pinus massoniana plantations agewB/(g·kg-1)
3.2 土壤酶活性及其比值的变化
ACP酶活性在不同林龄间没有显著差异,βG、NAG和ALP酶活性在不同林龄间有显著性差异, 但4种酶活性都表现为中龄林最高、成熟林最低, 且βG酶活性和NAG酶活性的变化趋势为中龄林>过熟林>幼龄林>成熟林, ACP和ALP酶活性变化趋势为中龄林>幼龄林>过熟林>成熟林(图1, 其中酶活性为平均值, 下同)。4种酶活性在0~10 cm土层最高。在0~10 cm土壤层中, 中龄林和过熟林的βG、NAG酶活性高于幼龄林和成熟林; 幼龄林和中龄林的ACP、ALP酶活性高于成熟林和过熟林。在10~20 cm土壤层中, 4种龄林的βG和ALP酶活性与在0~10 cm土层内的变化趋势大致一致, 中龄林和成熟林的NAG、ACP酶活性高于幼龄林和过熟林。 在20~30 cm的土壤层中, 幼龄林和中龄林的NAG、ACP、ALP酶活性高于成熟林和过熟林; 中龄林的βG酶活性最高, 过熟林最低(图2)。

图1 不同龄林马尾松人工林土壤酶活性差异Fig.1 Differences of soil enzyme activities at four age stands in Pinus massoniana plantations注: 不同小写字母表示马尾松人工林不同林龄间酶活性存在差异显著(p<0.05), 下同
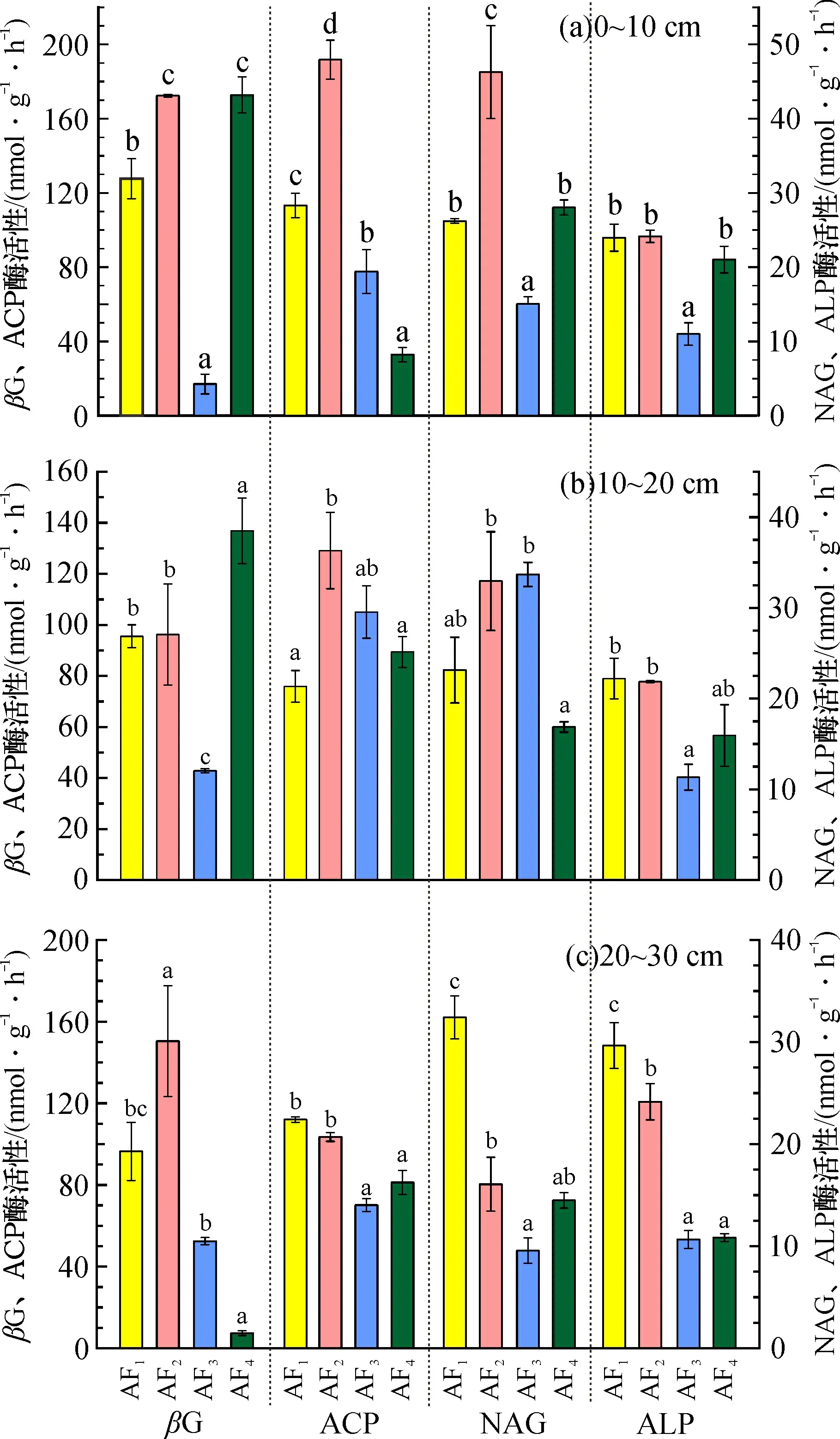
图2 不同土壤深度马尾松人工林酶活性的差异Fig.2 Differences of enzyme activities at different soil depths in Pinus massoniana plantations
土壤酶活性对数转化后, 4个龄林的土壤酶活性C∶N 值分别为1.21、1.17、1.16、1.20;土壤酶活性C∶P 值分别为1.00、1.06、0.86、1.09;土壤酶活性N∶P 值分别为0.76、0.84、0.71、0.83(表3)。

表3 不同龄林马尾松人工林酶活性生态化学计量比值特征Table 3 Characteristics of ecological stoichiometry ratio of enzyme activity in different Pinus massoniana plantations age stands
3.3 土壤酶活性及其比值与土壤养分的关系
Pearson 相关分析表明:βG、NAG、ACP、ALP酶活性皆与SOC、TN含量呈显著正相关关系(r分别为0.295、0.644、0.462、0.332, 0.305、0.595、0.463、0.420), 同时βG、NAG酶活性还与TP含量呈显著正相关关系(r分别为0.437和0.324);酶活性N∶P 值与SOC、TN极显著正相关(r为0.454和0.431); 酶活性C∶P 值与TP显著正相关(r为0.295); 酶活性C∶N 值与SOC、TN、AP呈负相关关系(表4)。
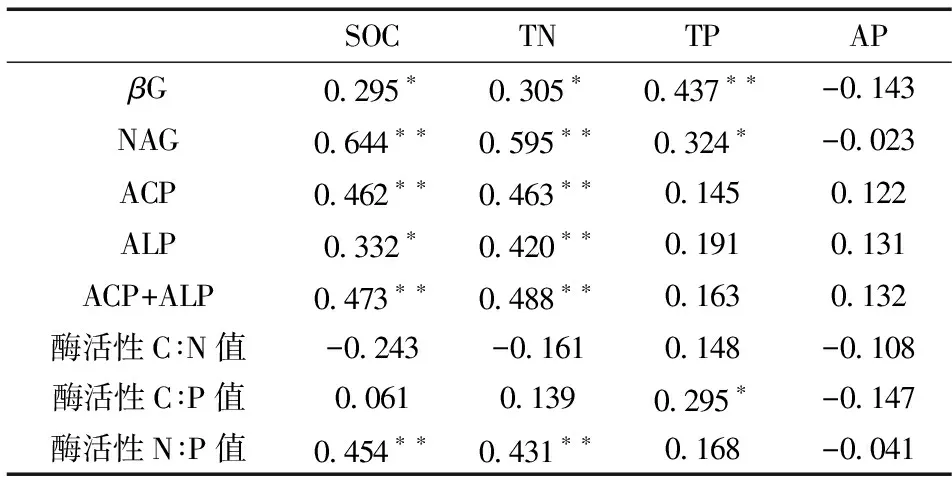
表4 马尾松人工林土壤养分与酶活性相关性
RDA 分析表明: 前两个排序轴保留了土壤酶活性数据总方差的55.92%, 累计解释率分别为26.95%和28.97%, 即土壤环境的SOC、TN、TP、AP四个因子累计解释土壤酶活性及其生态化学计量比的55.92%(图3)。其中,TP和TN分别对解释率贡献了55.2%和41.6%(p<0.05)(表5)。
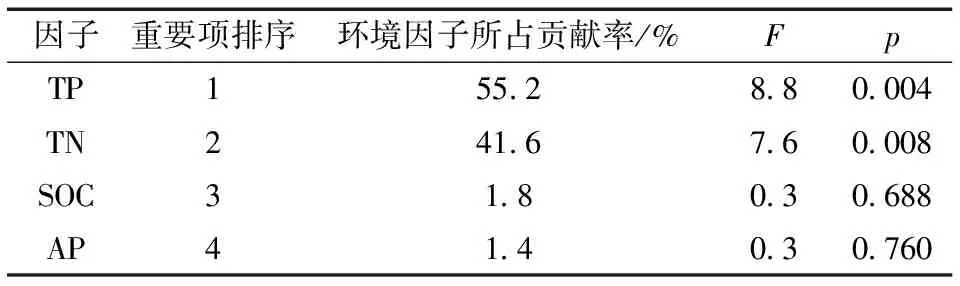
表5 环境因子解释的重要性排序和显著性检验结果
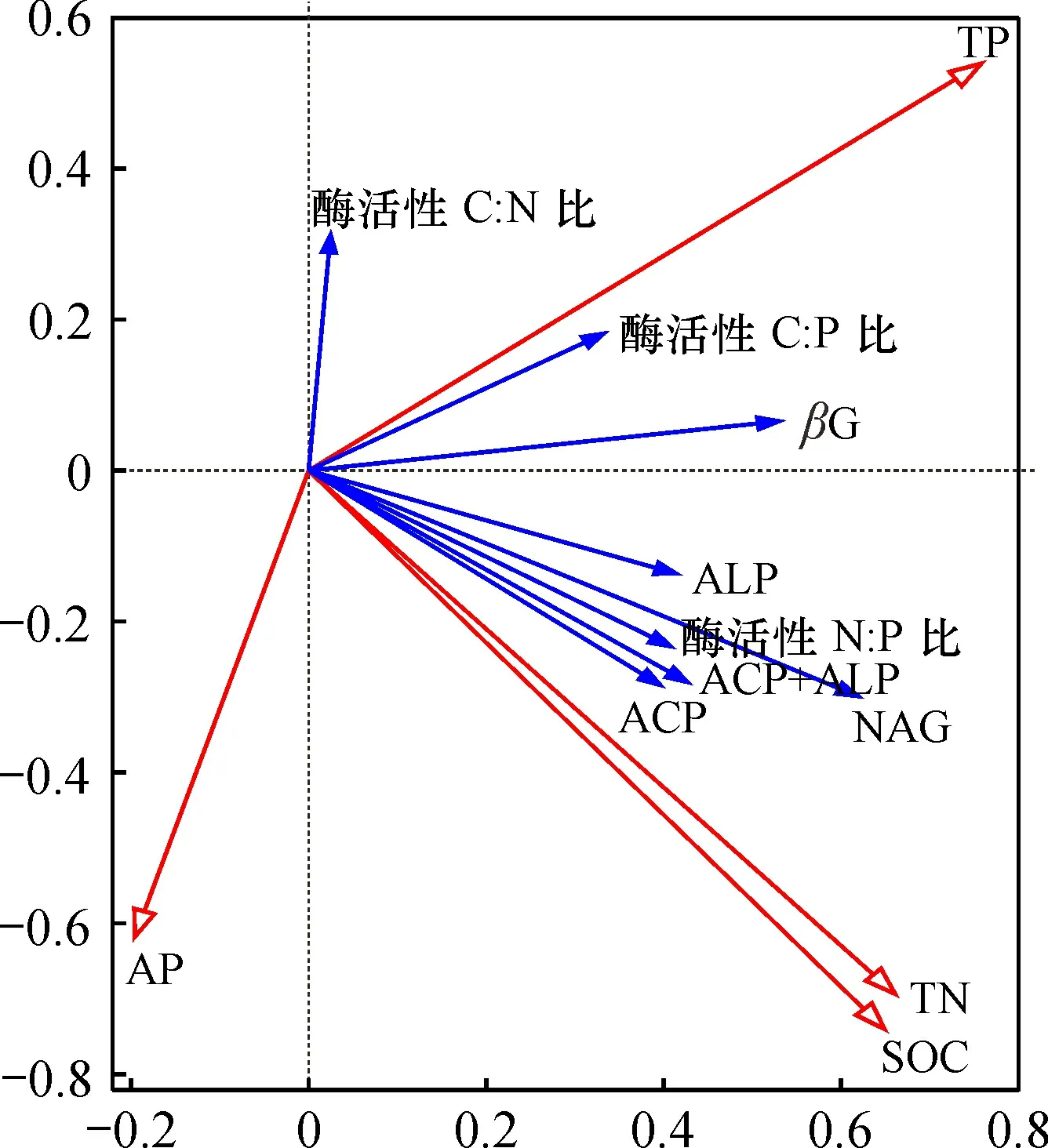
图3 马尾松人工林土壤酶活性及其化学计量比与土壤环境因子的 RDA 分析Fig.3 Soil enzyme activities and stoichiometric ratios of Pinus massoniana plantations and RDA analysis of soil environmental factors
4 讨 论
4.1 不同龄林土壤酶活性与养分的变化和联系
土壤酶在森林生态系统中的物质循环和能量流动过程中扮演着重要的角色, 监测人工林土壤酶活性是研究其土壤质量的重要内容, 这对人工林的经营和管理而言具有重要意义[26-28]。在本研究中, 0~30 cm的土壤深度对养分和酶活性会产生影响, 土壤营养元素循环与土壤剖面结构息息相关、不同土壤酶活性对土壤层次的响应也不同[29]。参考庞威[29]和杨瑞等[30]的研究, 出现这一结果可能是由于土壤表层的动植物残体和微生物数量最多、根系分布密集, 而土壤酶的主要来源是微生物、动植物残体及植物根系分泌物, 因此在0~10 cm土壤层中, 其分泌的酶多, 酶的活性相应就高, 而随着土壤深度的增加, 这种活性减弱, 土壤腐殖质含量减少、土壤养分含量也逐渐降低。因此0~10 cm土壤层的酶活性和养分含量会高于10~20 cm和20~30 cm。养分含量和土壤酶活性的变化也说明了在同一土壤深度中, 土壤中有机质转化能力相近, 土壤肥力变化不大。
相对于土壤深度, 林龄对养分和酶活性的影响更为显著, 不同龄林的βG和NAG酶的活性、ACP和ALP酶的活性变化趋势分别为: 中龄林>过熟林>幼龄林>成熟林、中龄林>幼龄林>过熟林>成熟林, 同时在不同龄林时βG、NAG和ALP酶活性有显著差异, ACP酶活性没有显著差异; 随着龄林的增长, SOC含量变化不大, TN含量逐渐降低, TP含量随着龄林的增长呈现升高—降低—升高的趋势, 即幼龄林和中龄林的TN含量较高, 且中龄林和过熟林的TP含量大于幼龄林和成熟林, AP含量的变化趋势与TP相反。表明不同的龄林对土壤酶活性有影响, 尤其是对βG、NAG和ALP酶活性影响显著, 且4种土壤酶活性均表现为在中龄林最高、在成熟林最低; 同时, 不同龄林对土壤养分也有影响, 成熟林的SOC含量最高、幼龄林的有效P含量最高, 中龄林和过熟林的总N含量和总P含量较高。
土壤与森林的生长是相互影响, 相互制约的关系[31]。研究表明, 马尾松人工林在快速生长的幼龄林和中龄林时期, 需要大时的P素[32-33]。因此在幼龄林时, 土壤N、P含量较高, NAG酶和磷酸酶活性也较高。结合梅杰[34]的研究, 中龄林的根系活力最强, 分泌物最多, 土壤微生物最活跃, 有利于有机物分解转化, 因此在中龄林时, 土壤酶活性和土壤养分含量均处在较高水平。成熟林时生长已趋于稳定, 郁闭度增大、林下植被盖度下降, 土壤微生物活性降低从而使地表枯落物分解变慢, 土壤接收的有机质含量减少, 氮、磷含量降低, 酶活性随之也降低[35]。过熟林时马尾松已近衰亡, 这时凋落物量较大, 而凋落物是土壤有机质的最主要来源, 因此过熟林的有机质含量较高, 同时凋落物和大量死根的腐解归还会给土壤提供碳源, 微生物活性增高从而过熟林的βG酶和NAG酶活性也较高[36]。氮、磷元素作为植物体的营养元素, 生长初期在植物体内都有较高的积累, 但是经植物生长旺盛期的波动变化后, 土壤中氮、磷含量降低, 特别是速效养分消耗较大[37-38],这与本研究的结果一致, 马尾松人工林土壤在幼龄林时具有较高的AP含量, 但是经过快速生长期后, 中龄林的AP含量低于其他3种龄林, 成熟林的马尾松人工林生长趋于稳定, AP含量逐渐提升。
4.2 土壤酶活性比值与马尾松人工林养分限制的关系
根据Sinsabaugh等[23]研究, 酶活性C∶N、C∶P和N∶P值全球平均值分别为1.41、0.62、0.44。土壤酶活性比值关系对土壤养分有至关重要的影响, 它们不仅推动着土壤 C、N、P等养分的转化与循环, 同时对 C、N、P 元素的平衡协调情况也产生显著的影响[39-40]。酶活性生态化学计量特征的研究为环境养分提供了可利用性信息, 进而对土壤的养分限制提供了指示[41]。酶活性N∶P值高于全球均值, 表明研究地区微生物代谢受到N限制, 而酶活性C∶P值高于全球均值时表明受P限制[42-43]。本研究中, 4种龄林的酶活性C∶N值分别为1.21、1.17、1.16、1.20, 均小于全球平均值(1.41); 酶活性C∶P和N∶P值分别为1.00、1.06、0.86、1.09和0.76、0.84、0.71、0.83, 均大于平均值(0.62和0.44)。林诚等[44]研究发现酶活性生态化学计量特征受土壤C、N、P 资源有效性的调控, 本研究结果发现TP、TN含量对土壤酶活性生态化学计量比值的影响显著, 具体表现为:TN含量与酶活性N∶P值极显著正相关, TP含量与酶活性C∶P值显著正相关。结合本研究地区的土壤酶化学计量比值可知, 该地区马尾松人工林的微生物代谢往往对N源酶的投入低于P源酶, 而微生物会根据自身养分的需求分泌酶来获取养分, 因此它们将分配更多资源以获取最为限制的养分[39]。同时, 本研究地区的马尾松人工林随着龄林的增长, 其土壤SOC含量变化不大、TN含量、TP含量、酶活性N∶P值和酶活性C∶P值的变化趋势均为升高—降低—升高, 说明该地区马尾松人工林C含量充足, N含量较低, P限制较大。
综上, 该研究地区的马尾松人工林受到P养分的限制。这与陈建会等[45]、Dessaux等[46]和樊纲惟等[47]的研究结果一致, P是热带和亚热带地区最难溶、最难移动的养分限制因子, 亚热带常绿阔叶林区域土壤受磷限制情况严重。除此之外, 结合4种龄林的酶活性N∶P值和酶活性C∶P值的大小发现, 本研究地区受P养分的限制程度随着龄林的增长呈降低—升高—降低趋势, 表明幼龄林、中龄林和过熟林受到P养分限制更为严重, 成熟林时养分含量相对稳定, 与其他研究结果相一致[48]。从生物学角度而言, 森林从幼龄林阶段到成熟林阶段是其生物量不断累积的过程[49]。因此, 可在幼龄林和中林龄时适当施加磷肥以促进其生长发育。
5 结 论
(1)马尾松人工林的土壤βG、NAG和ALP酶活性在不同龄林间具有显著差异, ACP酶活性在不同龄林间的差异不显著; 4种土壤酶活性均表现为在中龄林最高, 在成熟林最低。土壤酶活性在不同土壤深度具有差异, 在上层显著大于下层。
(2)土壤酶活性及其化学计量值受影响的土壤养分因子由大到小分别为TP、TN、SOC和AP, 表明较低的土壤P和N含量显著影响土壤酶活性及其生态化学计量特征的变化。
(3)马尾松人工林土壤酶活性C∶N值小于全球均值, 且随着林龄增大先降低后升高; 土壤酶活性C∶P和N∶P值大于全球均值, 随着林龄增大表现为升高—降低—升高趋势。说明马尾松受到P养分的限制, 在幼龄林和中龄林受限制程度更为严重, 成熟林养分含量相对稳定。
猜你喜欢
江西农业大学学报(2022年1期)2022-02-04
林业资源管理(2021年2期)2021-06-07
江西农业大学学报(2021年2期)2021-05-16
福建农林大学学报(自然科学版)(2020年6期)2020-11-26
锦绣·下旬刊(2020年12期)2020-01-03
农业与技术(2018年16期)2018-11-28
农村经济与科技(2018年15期)2018-05-14
森林工程(2018年5期)2018-05-14
现代农业科技(2017年10期)2017-07-12
广西林业科学(2016年2期)2016-03-20
