
菌酶协同发酵豆粕替代鱼粉对大口黑鲈生长性能、血清生化、免疫和抗氧化指标及肝脏组织形态的影响
2021-06-05 02:58陈晓瑛陈绍坚王国霞吴浩敏曹俊明孙育平
动物营养学报 2021年5期
陈晓瑛 陈绍坚 黄 文 王国霞 黄 敏 吴浩敏 曹俊明* 孙育平**
(1. 广东省农业科学院动物科学研究所,农业农村部华南动物营养与饲料重点实验室,广东省畜禽育种与营养研究重点实验室,广州 510640;2.广州飞禧特生物科技有限公司,广州 510642;3.广东海纳川生物科技股份有限公司,佛山 528500)
蛋白质是饲料中最关键的营养组成,也是决定饲料品质和价格的重要因素。鱼粉因蛋白质含量高、氨基酸种类组成平衡、营养丰富、诱食性好、消化率高等特点,一直以来都是水产饲料最重要的蛋白质来源[1]。然而,随着水产养殖规模的不断扩大,鱼粉供给已成为制约水产养殖健康可持续发展的制约因子。因此,水产饲料中鱼粉替代物的寻找及其高效利用备受行业关注。
大口黑鲈俗称加洲鲈,为一种典型温水肉食性鱼类,具适应性强、生长快、肉质鲜美和病害少等优点,已成为我国重要的淡水养殖品种。生产实践表明,一般大口黑鲈实用饲料蛋白质需求>45%,其中鱼粉用量为35%~55%,对鱼粉依赖性强。在鱼粉供需严峻的形势下,开展鱼粉替代物研究有利于大口黑鲈养殖业的健康持续发展。生产实践表明,微生物发酵后豆粕品质不稳定,此外单一微生物发酵法的产酶量较低,无法满足实际生产需求。菌酶协同发酵豆粕(FSBE)即在酶解工艺的处理下加入一定量的乳酸菌、酵母菌、芽孢菌、霉菌等益生菌进行发酵。益生菌在发酵过程中会产生多种香味物质,可对豆粕产品的苦味发挥调节作用,同时克服单独利用微生物发酵产酶不足的问题。近年来研究表明,FSBE效果较微生物发酵法和酶单独处理要好,其可缩短抗原削减周期,利于工业化生产;利用豆粕中糖代谢生成乳酸及其他各种有机酸,降低pH,从而发挥抑菌作用,延长豆粕产品保质期,此外发酵产生的多种有机酸能增加豆粕的营养性和适口性,这对于豆粕高效利用和饲料制备具有重大意义[16]。目前,关于豆粕替代大口黑鲈饲料中鱼粉的研究报道主要集中于去皮豆粕、酶解豆粕等方面[12-13,17-20],而有关FSBE替代饲料中鱼粉的研究尚未见报道。为此,本试验使用FSBE替代饲料中不同比例鱼粉,并设置普通豆粕替代30%鱼粉为负对照组,研究其对大口黑鲈生长性能、血清生化、免疫和抗氧化指标及肝脏组织形态的影响,旨在评估FSBE替代鱼粉的可行性,为FSBE在大口黑鲈配合饲料中的应用提供参考。
1 材料与方法
1.1 试验材料
FSBE制备所需原料和发酵工艺均由佛山某生物科技有限公司提供。在风干豆粕中加入水、酶制剂(中性蛋白酶活性>1 000 U/g),并接种微生物(1 000 CFU/g枯草芽孢杆菌和500 CFU/g植物乳杆菌),搅拌均匀后将湿度保持约40%,35 ℃恒温条件下密封持续发酵48 h。烘干、粉碎,过60目筛后,封装于塑料自封袋中于-20 ℃冰箱保存备用。FSBE呈浅棕黄色,色泽一致,且无结块霉变、氧化酸败、异臭味等现象。鱼粉、豆粕均由广东某饲料公司提供。鱼粉、FSBE和豆粕的营养组成见表1。
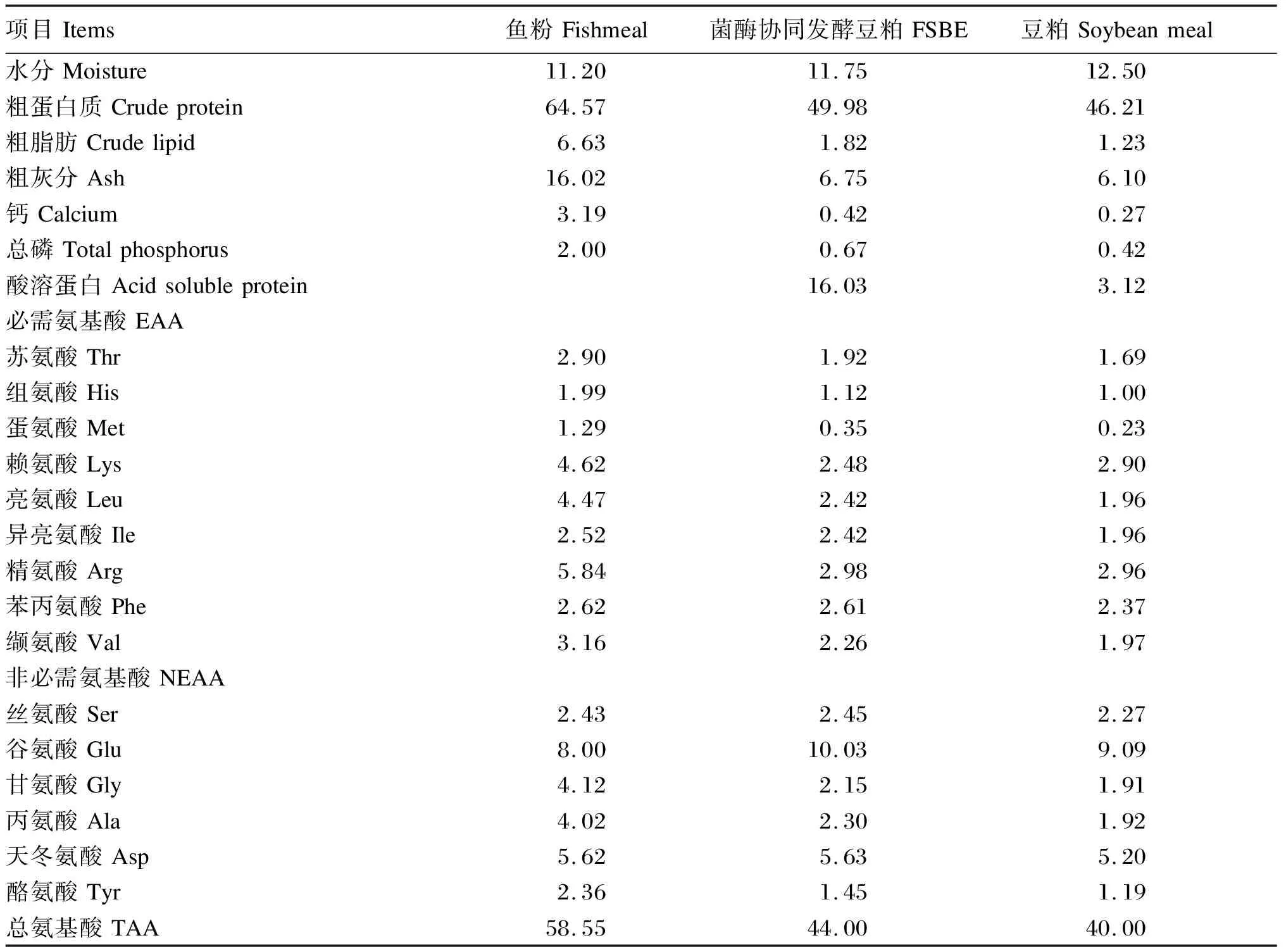
表1 鱼粉、FSBE和豆粕的营养组成(干物质基础)
1.2 试验设计与饲料组成
根据大口黑鲈营养需求,对照饲料(T0)以鱼粉、豆粕、玉米蛋白粉、血粉及鸡肉粉为蛋白质源;鱼油、豆油及大豆卵磷脂为脂肪源;α-淀粉为糖源。采用等氮(50%粗蛋白质)等脂(10%粗脂肪)原则,在对照组基础上用FSBE分别替代10%(FSBE10)、20%(FSBE20)、30%(FSBE30)和40%(FSBE40)鱼粉,并用豆粕替代30%鱼粉为负对照饲料(SB30),在除T0组外的各组饲料中补充赖氨酸、蛋氨酸,共配制6种试验饲料。各组饲料组成及营养水平见表2,饲料氨基酸组成见表3。将所有饲料原料粉碎过60目筛,根据饲料配方按逐级扩大法逐级混合后,再置于V型混合机混匀,然后转入油脂捏合机(NH-10型,华南理工大学)中加入鱼油、豆油及大豆卵磷脂进行充分捏合、混匀。将混合均匀的原料转入饲料搅拌机(B20型,广州市番禺力丰食品机械厂)中,加入适量水充分搅拌混合10 min。用双螺杆挤压机(SLX-80型,华南理工大学)制成粒径为2.5 mm的颗粒饲料,55 ℃烘干,自然冷却过20目筛后放入密封塑料袋中,置于-20 ℃冰箱保存备用。
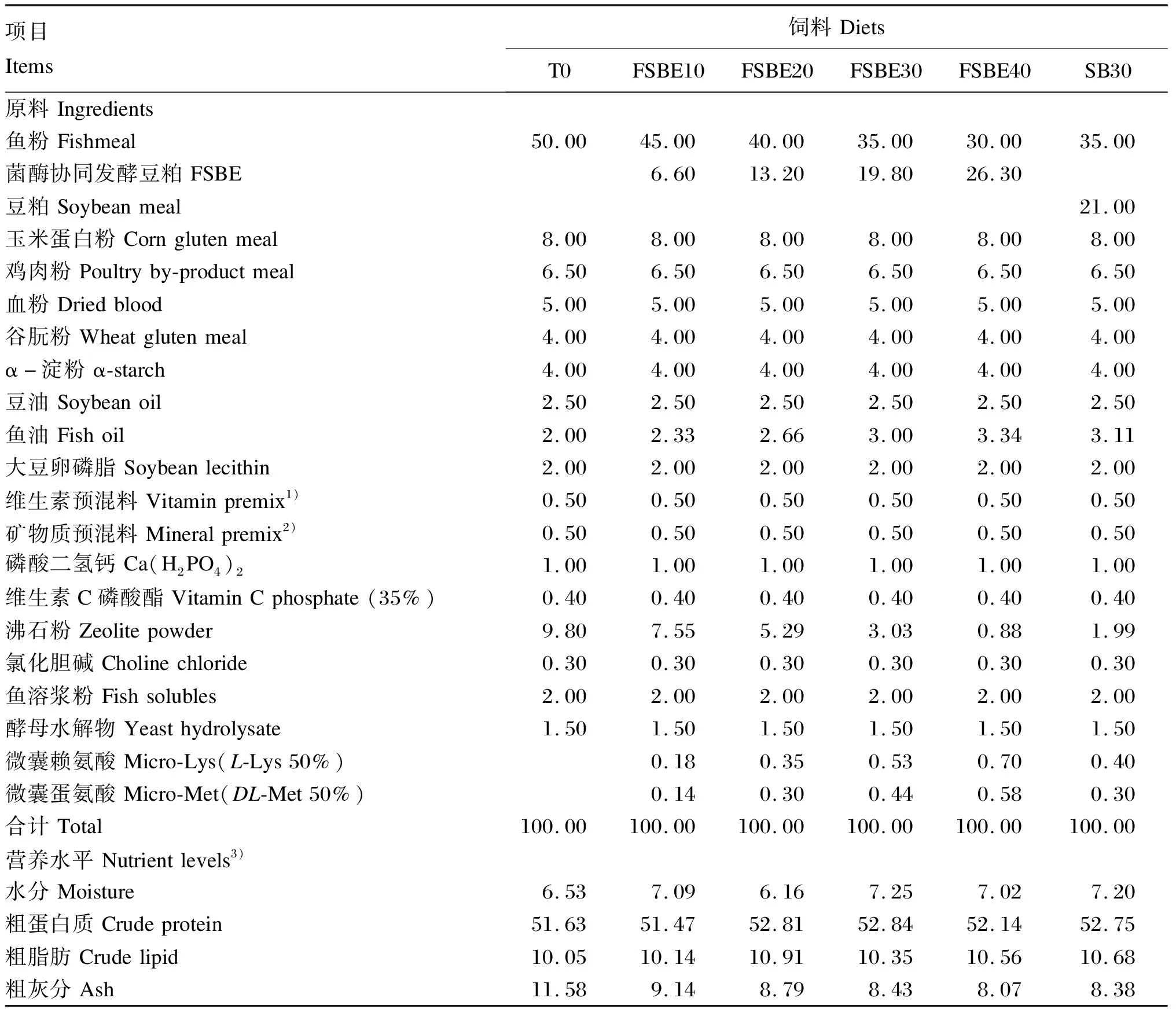
表2 饲料组成及营养水平(干物质基础)

表3 饲料氨基酸组成(干物质基础)
1.3 试验鱼及饲养管理
大口黑鲈购自广东省阳山利阳水产科技有限公司,养殖试验在广东省农业科学院动物科学研究所室内循环水养殖系统中进行。室内循环水系统中暂养2周,并每天(08:30和18:30)饱食投喂商用配合饲料(50%粗蛋白质,11%粗脂肪)。试验开始前大口黑鲈禁食24 h,挑选体质健壮、活泼、体表完好、平均体重为(5.08±0.01) g的大口黑鲈幼鱼600尾,随机分为6组,每组4个重复,每个重复25尾,放养于由24个容积为350 L的圆柱形玻璃纤维钢桶(直径80 cm,高70 cm,实际水容量为300 L)组成的室内循环水养殖系统中,分别投喂6种试验饲料。每天表观饱食投喂试验饲料2次(08:00和18:30),投喂1 h后收集残饵并烘干。试验期间养殖用水为经砂滤、曝气后自来水,进水速率为1.3~1.5 L/min。养殖过程中连续充气,并视水质情况换水,每次换水量为1/3。每天观察大口黑鲈健康与摄食状况,并记录投饲量、水温及死亡情况,并定期检测水质指标。养殖期间水温变化范围为25.0~30.0 ℃,溶氧含量>5.0 mg/L,pH 6.8~7.2,氨氮含量<0.10 mg/L,亚硝酸盐含量<0.05 mg/L,自然光照。饲养周期为8周。
1.4 样品采集和指标测定
养殖试验结束后饥饿24 h,依次捞取每个桶中的鱼,称重、计数,计算生长性能指标。从每桶中随机捞取接近平均体重的15尾鱼,放入含MS-222(40 mg/L)的水溶液中麻醉。其中3尾鱼保存在-20 ℃冰箱中,用于全鱼体成分测定;12尾鱼称重、测体长后,采用1 mL无菌注射器从尾静脉处取血,并转入5.0 mL离心管收集,室温静置2 h后离心(4 000 r/min,10 min),收集上清,于-80 ℃冰箱保存,用于血清生化、免疫和抗氧化等指标测定。采血后,按顺序每桶随机选择9尾鱼解剖,称量内脏团、肝脏及腹腔脂肪重,计算形体指标。每桶随机取3尾鱼,分离肝脏用Bouin’s液固定,用于组织切片制作。
试验饲料、全鱼水分含量采用105 ℃常压干燥法测定(GB/T 5009.3—2016);粗蛋白质含量采用凯氏定氮法(N×6.25)测定(GB/T 6432—2018);粗脂肪含量采用索氏抽提法测定(GB/T 6433—2006),抽提试剂为石油醚;粗灰分含量采用550 ℃高温炉灼烧法测定(GB/T 6438—2007)。原料及饲料中氨基酸含量采用盐酸水解后(GB/T 18246—2019),用Waters高效液相色谱仪(510型,美国)检测分析。血清葡萄糖(GLU)、尿素氮(UN)、总蛋白(TP)、胆固醇(CHO)、甘油三酯(TG)、高密度脂蛋白胆固醇(HDL-C)、低密度脂蛋白胆固醇(LDL-C)含量及谷丙转氨酶(GPT)、谷草转氨酶活性(GOT)活性委托广州金域医学检验中心,采用全自动生化分析仪(日立7600,日本)测定。血清总抗氧化能力(T-AOC)、总超氧化物歧化酶(T-SOD)及谷胱甘肽过氧化物酶(GSH-Px)、过氧化氢酶(CAT)、碱性磷酸酶(AKP)、酸性磷酸酶(ACP)、溶菌酶(LZM)活性及丙二醛(MDA)含量均采用南京建成生物工程研究所试剂盒测定,具体操作步骤按照试剂盒说明书进行。将用Bouin’s液固定的肝脏样本,经脱水浸蜡、透明、包埋、切片、展片及烤片后,用苏木精-伊红(HE)染色,并用中性树胶封固,用光学显微镜(Nikon 80i)观察、拍照。
1.5 计算公式
增重率(%)=100×[终末体均重(g)+死亡体重(g)-初始体均重(g)]/初始体均重(g); 特定生长率(%/d)=100×[Ln终末体均重(g)-Ln初始体均重(g)]/养殖天数(d);摄食率(%/d)=100×摄食总干物质量(g)/{[初始鱼数量(尾)+终末鱼数量(尾)]/2}/养殖天数(d);肥满度(condition factor,CF,g/cm3)=100×全鱼体重(g)/鱼体长(cm)3;脏体比(viscerosomatic index,VSI,%)=100×内脏重(g)/鱼体重(g);肝体比(hepatosomatic index,HSI,%)=100×肝脏重(g)/鱼体重(g);腹脂率(intraperitoneal fat,IPF,%)=100×腹腔脂肪重(g)/全鱼体重(g);蛋白质效率(%)=100×[终末体均重(g)-初始体均重(g)]/蛋白质摄入总量(g);脂肪效率(%)=100×[终末体均重(g)-初始体均重(g)]/脂肪摄入总量(g);饲料系数(FCR)=摄食饲料量/[终末体均重(g)+死亡鱼重(g)-初始体均重(g)];成活率(%)=100×终末鱼数量(尾)/初始鱼数量(尾)。
1.6 数据统计分析
试验数据采用Excel 2010进行整理,通过SPSS 23.0软件对数据进行单因素方差分析(one-way ANOVA)。若等方差,则采用Duncan氏法进行多重比较,若不等方差,则采用邓尼特T3检验法进行多重比较。试验所得数据用平均值±标准误表示,P<0.05表示差异显著。
2 结 果
2.1 FSBE替代鱼粉对大口黑鲈生长性能的影响
由表4可知,随着FSBE替代鱼粉比例的增加,大口黑鲈的终末体均重、特定生长率呈下降趋势,替代比例高于20%时均显著低于对照组(P<0.05);且FSBE30、FSBE40组均显著低于FSBE20组(P<0.05),但FSBE30、FSBE40组之间无显著差异(P>0.05)。FSBE替代鱼粉比例高于20%时会导致摄食率显著降低(P<0.05),而饲料系数则显著升高(P<0.05);而替代比例高于10%时蛋白质效率和脂肪效率显著降低(P<0.05)。其中,FSBE30组摄食率与饲料系数最高,而蛋白质效率和脂肪效率最低,且与对照组相比差异显著(P<0.05)。大口黑鲈成活率各组间无显著差异(P>0.05)。与SB30组相比,FSBE30组大口黑鲈的摄食率、饲料系数、蛋白质效率和脂肪效率存在显著差异(P<0.05),而终末体均重、特定生长率和成活率则无显著差异(P>0.05)。
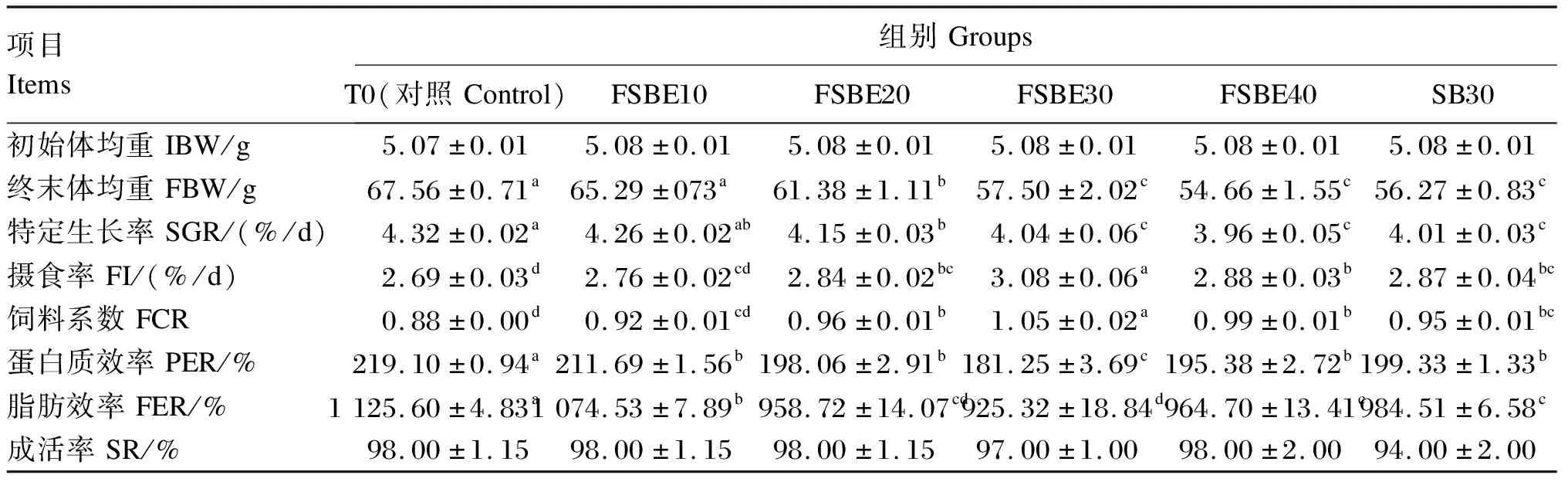
表4 FSBE替代鱼粉对大口黑鲈生长性能的影响
2.2 FSBE替代鱼粉对大口黑鲈形体指标的影响
由表5可知,FSBE替代鱼粉对大口黑鲈的HSI和VSI影响显著(P<0.05),而对CF和IPF影响不显著(P>0.05)。随着替代比例的增加,CF和IPF呈先增后降趋势,而HSI和VSI呈下降趋势。FSBE30组HSI最低,FSBE10组VSI最低,且均显著低于对照组(P<0.05),但各替代组间的HSI、VSI均无显著差异(P>0.05)。FSBE30组VSI和IPF均显著高于SB30组(P<0.05);FSBE30组CF和HSI均高于SB30组,但差异不显著(P>0.05)。

表5 FSBE替代鱼粉对大口黑鲈形体指标的影响(湿重基础)
2.3 FSBE替代鱼粉对大口黑鲈体成分的影响
由表6可知,FSBE替代鱼粉对全鱼水分、粗蛋白质、粗灰分和总磷含量无显著影响(P>0.05),而对粗脂肪、钙含量影响显著(P<0.05)。随着FSBE替代鱼粉比例的增加,全鱼水分含量呈上升趋势,粗蛋白质和粗脂肪含量呈下降趋势,而粗灰分、钙和总磷含量则呈先升后降趋势。其中,FSBE30组全鱼粗灰分、钙及总磷含量最低,且钙含量显著低于对照组(P<0.05)。除总磷外,FSBE30组全鱼水分、粗蛋白质、粗脂肪、粗灰分及钙含量均高于SB30组,但差异不显著(P>0.05)。
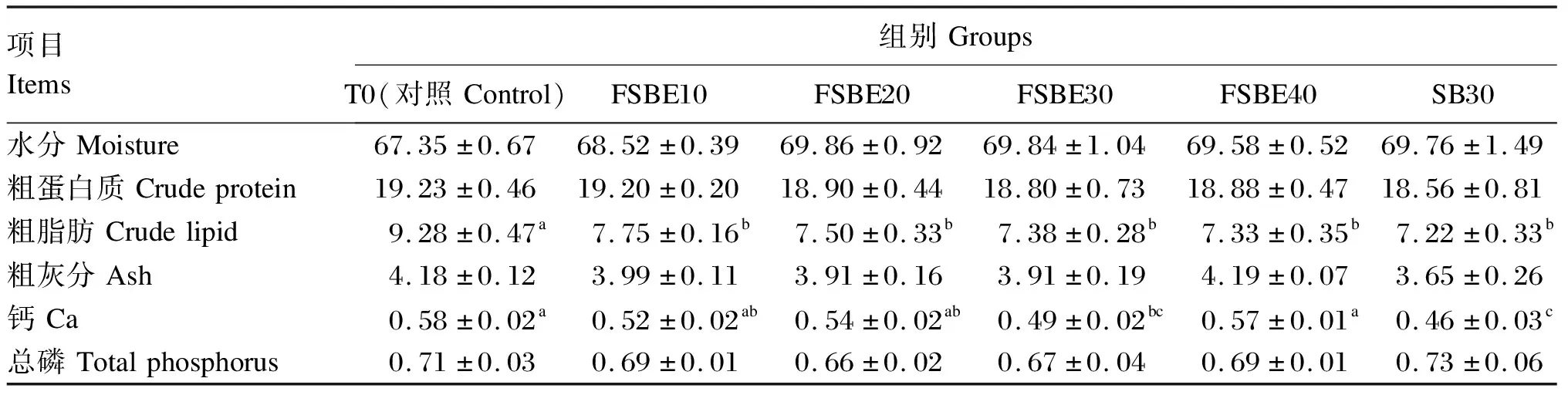
表6 FSBE替代鱼粉对大口黑鲈体成分的影响(湿重基础)
2.4 FSBE替代鱼粉对大口黑鲈血清生化指标的影响
由表7可知,FSBE替代鱼粉仅对血清中的TG、HDL-C、GLU含量及GPT和GOT活性产生显著影响(P<0.05),而对其他血清生化指标的影响不显著(P>0.05)。随着FSBE替代鱼粉比例增加,血清中TG含量呈下降趋势,而GPT活性、HDL-C含量则与此相反。各试验组中,FSBE30组血清中的TP、UN、TG、GLU含量及GPT活性最低,且TG含量和GOT活性与对照组存在显著差异(P<0.05)。除FSBE10组外,对照组血清中TG含量均显著高于其他各试验组(P<0.05),而GOT活性则显著降低(P<0.05)。FSBE30组血清HDL-C含量最高,且显著高于对照组和FSBE10组(P<0.05),但对照组和FSBE10组之间差异不显著(P>0.05)。SB30组血清中TP、UN含量高于FSBE30组(P>0.05),SB30组血清中的GPT活性则显著高于FSBE30组(P<0.05)。而SB30组血清中的CHOL、TG、LDL-C、GLU含量及GOT活性均低于FSBE30组,但差异不显著(P>0.05)。
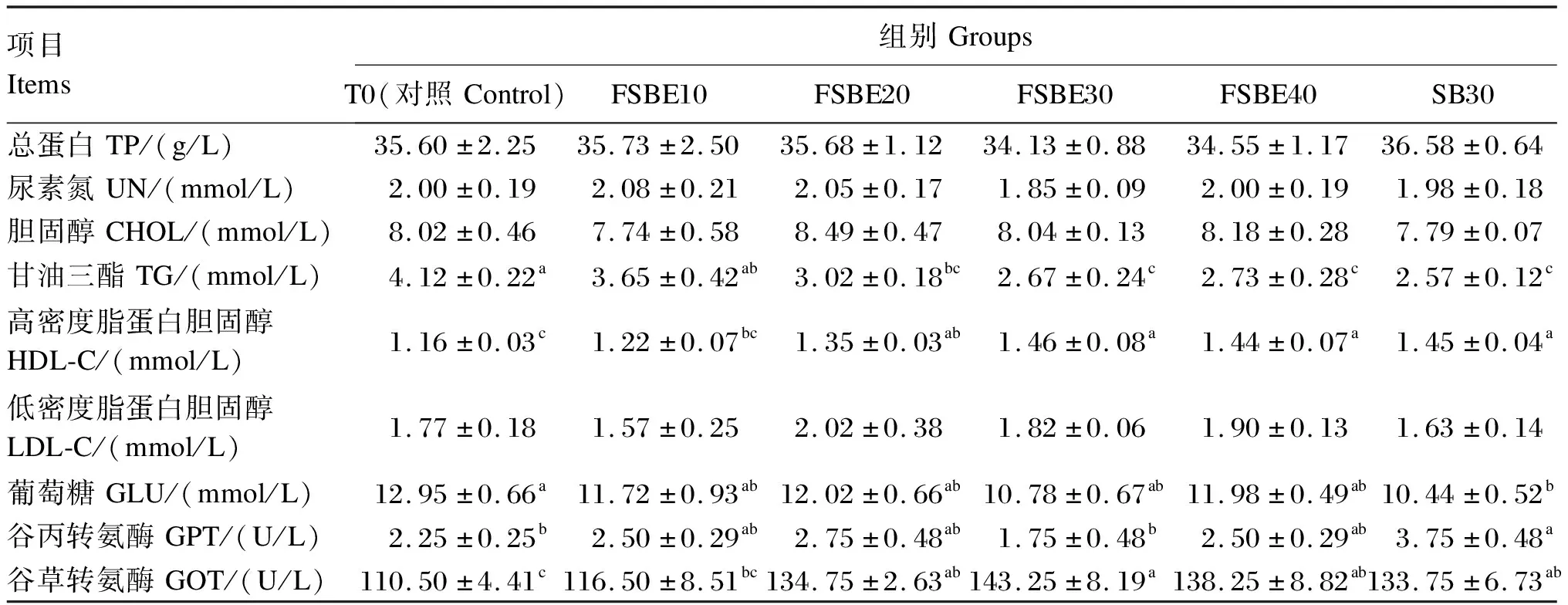
表7 FSBE替代鱼粉对大口黑鲈血清生化指标的影响
2.5 FSBE替代鱼粉对大口黑鲈血清免疫和抗氧化指标的影响
由表8可知,除CAT外,FSBE替代鱼粉对血清中T-AOC、T-SOD、GSH-Px、AKP、ACP、LZM活性及MDA含量产生显著影响(P<0.05)。随着FSBE替代鱼粉比例增加,血清中T-SOD及ACP活性升高;T-AOC、GSH-Px和LZM活性呈先升后降趋势;而MDA含量降低。其中,与对照组相比,FSBE20组血清T-SOD、AKP、ACP及LZM活性及MDA含量存在显著差异(P<0.05)。除T-AOC和LZM外,FSBE30组其他血清免疫和抗氧化指标均高于SB30组,且MDA含量及AKP、ACP及LZM活性在二者间差异显著(P<0.05)。
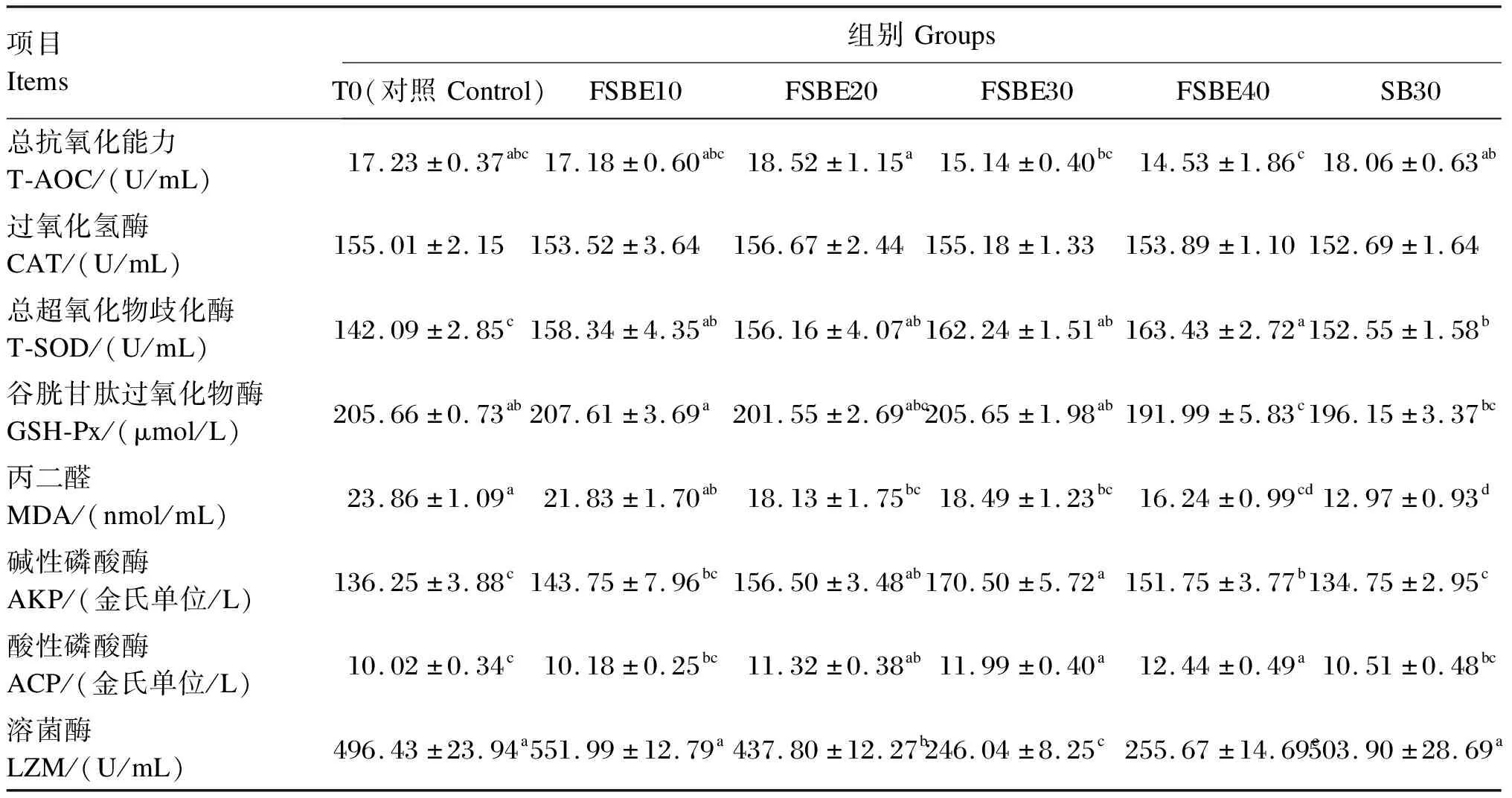
表8 FSBE替代鱼粉对大口黑鲈血清免疫和抗氧化指标影响
2.6 FSBE替代鱼粉对大口黑鲈肝脏组织形态的影响
如图1所示,各试验组织中均广泛可见大量肝细胞呈空泡状(黑色箭头)。对照、FSBE10组肝细胞排列较整齐,细胞核和膜界限清晰,肝脏组织未见明显炎症,但随着FSBE替代鱼粉比例增加,肝细胞轮廓模糊,细胞核朝质膜移位,肝脏血窦扩张;且肝脏局部血管周围的可见炎性细胞灶性浸润(黄色箭头)程度加剧。较FBSE30和FBSE40组而言,SB30组肝脏血窦扩张及其局部血管周围的可见炎性细胞灶性浸润(黄色箭头)程度最为严重。
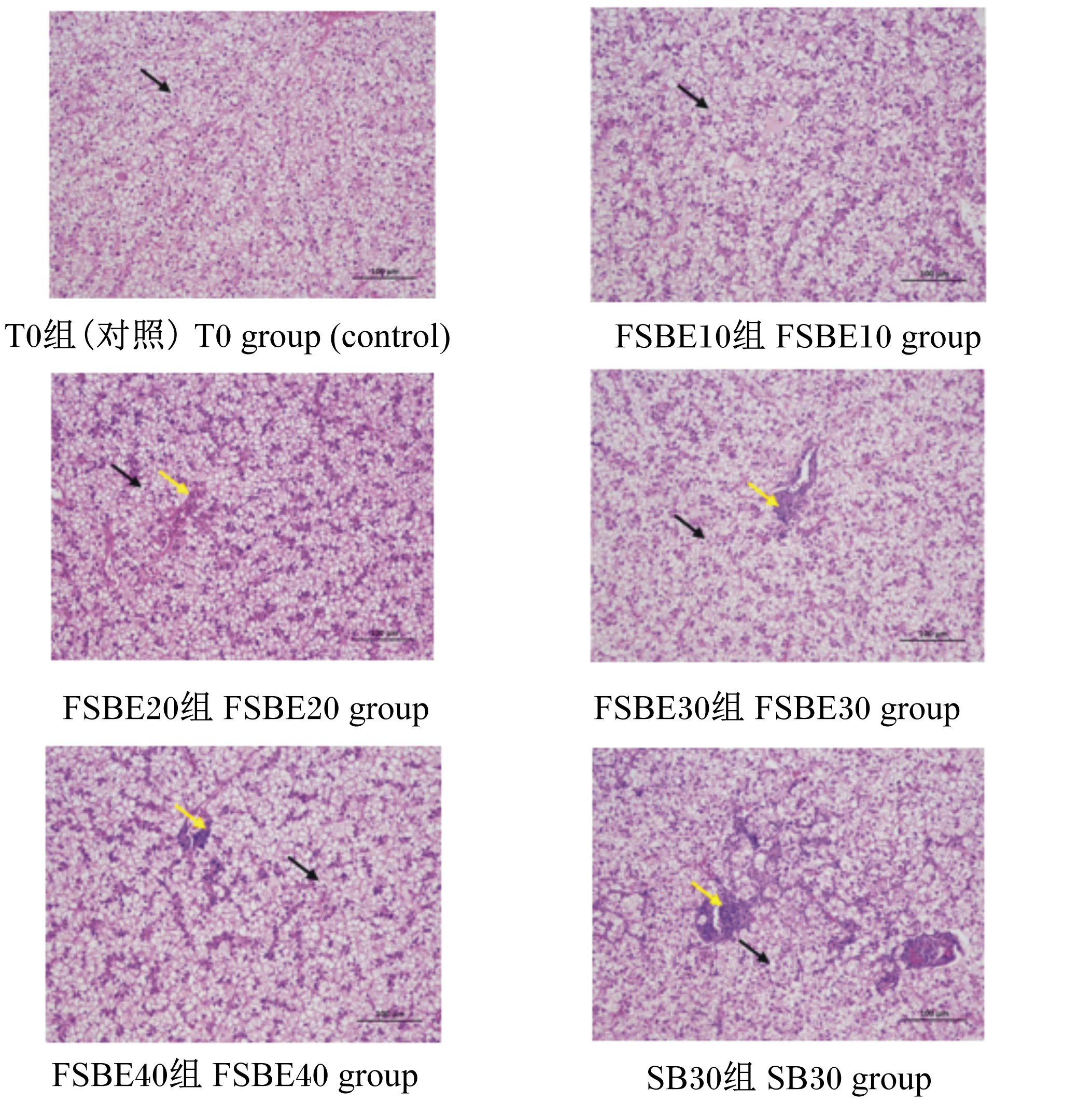
图1 FSBE替代鱼粉对大口黑鲈肝脏组织形态的影响(HE染色)
3 讨 论
3.1 FSBE替代鱼粉对大口黑鲈生长性能的影响
相比其他蛋白质原料,鱼粉具营养品质、适口性等优势,在水产动物、特别是肉食性种类配合饲料广泛应用。但鱼粉价格居高、资源有限,饲料中高鱼粉用量会直接导致养殖成本增加。因此,随着水产养殖规模不断扩大,其配合饲料中鱼粉替代物研究成为人们关注焦点。豆粕是水产动物饲料中鱼粉重要的替代物质,但过高水平饲喂后因适口性差、必需氨基酸失衡、磷利用率低、抗营养因子(ANFs)含量高以及易引起脂质代谢紊乱等因素,会导致生长性能下降[1-2]。豆粕经发酵后可削减,甚至消除抗营养因子,改善营养品质,提高消化率[16,21]。罗智等[22]研究发现,同等替代比例发酵豆粕组石斑鱼(Epinepheluscoioides)增重率、特定生长率、饲料系数和蛋白质效率均高于豆粕组。普通豆粕可替代牙鲆(ScophthalmusmaximusL.)饲料中30%鱼粉,而发酵豆粕则可替代45%鱼粉[23]。何明等[18]研究发现,发酵豆粕替代大口黑鲈饲料中30%鱼粉对饲料系数和摄食量无显著影响,但同等普通豆粕比例替代后摄食量和饲料系数则显著升高,表明发酵豆粕营养价值优于豆粕。彭松等[24]在凡纳滨对虾、罗智等[22]在石斑鱼以及Yammoto等[25]在虹鳟(Oncorhynchusmykiss)的研究结果相似。本试验中同等比例替代FSBE组特定生长率和成活率等均优于普通豆粕组,进一步证明发酵豆粕价值优于普通豆粕。饲料系数升高表明饲料利用效率下降,摄食率升高表明抢食能力增强。但本试验结果发现,同等比例替代鱼粉的FSBE的饲料系数和摄食率均高于普通豆粕,而蛋白质效率和脂肪效率则低于普通豆粕,与以往研究结果不一。原因可能与FSBE改善了饲料适口性,导致抢食能力强、摄食率升高[16]。过高比例FSBE替代鱼粉则会因部分营养素不平衡、营养吸收利用不同步,从而导致饲料系数升高[26],但具体原因有待进一步研究探讨。
一般而言,发酵豆粕替代水产动物饲料中鱼粉比例为16%~40%,过高发酵豆粕会导致生长性能下降。随着饲料中发酵豆粕替代鱼粉比例增加,斑点叉尾(Ietaluruspunetaus)[27]、凡纳滨对虾[3,5]、石斑鱼(Epinepheluscoioides)[22]、日本鳗鲡(Anguilljaponica)[7]生长性能呈先升后降趋势,其适宜替代比例分别为2.5%、8%、7%和20%。本研究中,饲料中FSBE替代鱼粉比例≥20%时,大口黑鲈特定生长率和摄食率显著下降,饲料系数显著升高,表明大口黑鲈饲料中FSBE适宜替代比例≤20%,超过20%则会对生长产生不利影响。何明等[18]研究发现,发酵豆粕替代饲料中≥30%鱼粉会导致大口黑鲈增重率显著下降,而饲料系数显著升高,本研究结果与此相似,说明大口黑鲈对鱼粉依赖性较强。
鱼体的形体指标受许多因素影响,如鱼体种类、生长阶段、营养素缺乏、代谢紊乱、抗营养因子、有毒有害物质等。形体指标可用来反映生长发育和健康状况。以往有关发酵豆粕对形体指标影响研究结果不一致。Barnes等[28]发现50% PepSoyGen(PSG,一种经曲霉和杆菌发酵后的豆粕)会导致虹鳟HSI显著下降,但对CF无显著影响。何明等[18]研究发现,发酵豆粕和普通豆粕对大口黑鲈CF和VSI均无显著影响。钟国防[19]报道,发酵豆粕替代鱼粉后大口黑鲈CF、HSI差异不显著,但对VSI影响显著(30%)。随着发酵豆粕替代比例增加,杂交罗非鱼[15]、圆斑星鲽(Veraspervariegatus)[29]的CF、HSI和VSI降低,其原因可能是发酵豆粕替代鱼粉对消化系统产生影响。此外,鱼体对抗营养因子敏感性也会影响内脏正常发育。肝脏是储存脂肪的重要器官,其脂肪含量降低会导致HSI下降。王新霞[12]研究发现,饲料中添加<10%的发酵豆粕,VSI和HSI显著下降,体脂肪沉积也减少,认为这与发酵豆粕降低了脂肪在体内的沉积有关。也有研究者认为HSI变小可能与饲料消化能降低及磷缺乏有关[30]。本试验中,随着FSBE替代比例升高,形体指标中VSI显著下降,HSI有下降趋势但不显著,这与钟国防[19]研究结果相似。本试验中,30% FSBE替代组各形体指标均优于30%普通豆粕替代组,且在VSI和IPF达到显著水平。张改改等[20]在研究发酵豆粕和普通豆粕同等比例替代鱼粉结果发现,相比对照组,发酵豆粕替代鱼粉可导致CF、VSI和HSI均显著降低,发酵豆粕组VSI和HSI显著高于豆粕组,本研究结果与此结果基本相似。这进一步说明豆粕和发酵豆粕在营养品质上存在差异,也说明饲料中添加植物蛋白质源可影响形体指标状况。
鱼体组成受饲料营养成分、食物组成、养殖水环境以及季节等外界条件影响。除遗传因素外,饲料营养是影响体组成的重要因素。林一帆等[14]研究发现,随着饲料中鱼粉被含50%发酵豆粕的复合蛋白质源替代比例的增加,青鱼(Mylopharyngodonpiceus)幼鱼全鱼和肌肉粗脂肪含量呈现上升的趋势,替代比例≥40%粗脂肪含量显著高于对照组。随着发酵豆粕使用量增加,日本鳗鲡黑仔鳗肌肉粗蛋白质含量呈现先减少后增加趋势,粗脂肪含量则相反[7]。饲料中使用PepSoyGen达50%时,虹鳟肌肉蛋白质含量显著降低,但水分、粗脂肪和粗灰分含量变化不显著[28]。随着发酵豆粕水平升高,圆斑星鲽[29]、斑点叉尾[27]和杂交罗非鱼[15]水分和粗灰分含量显著升高。但也有研究结果显示鱼体组成不受发酵豆粕水平的影响。发酵豆粕替代饲料中鱼粉对大口黑鲈肌肉中水分、粗蛋白质、粗脂肪和粗灰分含量均无显著影响[17-18],对花鲈(Lateolabraxjaponicus)[4]、石斑鱼[22]肌肉粗蛋白质和粗脂肪含量影响亦不显著;对杂交罗非鱼[15]粗灰分和粗脂肪含量影响不显著;对黑鲷[28]全鱼体成分和氨基酸组成影响不显著。冷向军等[5]发现,发酵豆粕水平对凡纳滨对虾肌肉水分和粗蛋白质含量影响不显著。上述结果说明,发酵豆粕对水产动物体组成影响结果因水产动物种类、饲料配方及养殖条件而迥异。本试验研究发现,FSBE替代鱼粉显著降低了大口黑鲈全鱼粗脂肪、钙含量,与日本鳗鲡黑仔鳗[7]]结果相似。随着FSBE替代鱼粉比例升高,水分含量呈升高趋势,而粗蛋白质含量下降。一般认为,饲料中植物蛋白质原料升高会导致机体水分含量升高,粗蛋白质和粗脂肪含量下降[31]。鱼体脂肪,特别是肌肉脂肪对于机体健康、风味品质具有重要作用。本试验结果表明,FSBE和豆粕会对机体脂肪组成及血清脂肪代谢物质含量产生显著影响,但FSBE和豆粕同等比例替代鱼粉对这些指标的影响无显著差异。豆粕中含有的抗营养因子会降低机体对脂肪的消化吸收,从而可能引起机体脂肪沉积下降,但发酵豆粕调控脂肪代谢机理尚需进一步研究。
3.2 FSBE替代鱼粉对大口黑鲈血清生化、免疫和抗氧化指标的影响
鱼类血清生化指标与机体新陈代谢、营养吸收以及健康状况关系密切,是评价生理学、病理学的重要指标,被广泛应用于衡量机体代谢与健康状况。血清TP是反映机体蛋白质代谢和机体健康与否的重要指标。由本试验结果可知,随着FSBE替代鱼粉比例增加,血清TP含量保持相对稳定;同等比例替代鱼粉的FSBE的血清TP含量低于普通豆粕,与日本鳗鲡血清TP含量研究[7]结果相似,表明替代后大口黑鲈肝脏蛋白质合成功能正常,能满足蛋白质需要,未造成自身蛋白质的丢失[32]。但钟国防[19]发现发酵豆粕替代比例≥10%后,大口黑鲈血液中TP含量变化显著。血清TG是鱼体脂类转运的主要形式之一,主要来自肝脏,其含量变化可反映机体脂类吸收和代谢状况。本试验结果表明,FSBE替代鱼粉比例≥20%后,血清TG含量显著下降,与王经远等[33]研究结果相似,表明发酵豆粕提高了机体对脂类的吸收利用。HDL-C和LDL-C含量反映脂类在体内的分解转运以及代谢状况。本研究结果表明,高比例FSBE替代鱼粉导致血清HDL-C含量显著升高,而对LDL-C含量无显著影响,表明FSBE可能会干扰了脂类在大口黑鲈体内消化、吸收和沉积。血糖是机体主要功能物质,其含量受营养和采食量等影响。本试验结果表明,FSBE替代鱼粉对血清葡萄糖含量无显著影响,表明其对血糖代谢无不良影响[32]。
GPT和GOT是机体分布最广、活性最强的转氨酶,主要存在于肝脏,在蛋白质和脂肪代谢中具有重要作用。肝功能障碍或肝功能受损时,GPT和GOT会释放到血液中,引起血清转氨酶活性升高。随着FSBE替代鱼粉比例增加,血清中2种转氨酶活性升高,当替代鱼粉比例≥20%时,血清GOT活性升高显著,这可能与发酵豆粕中存在部分抗营养因子削减不全有关。当替代鱼粉比例过高时,抗营养因子及氨基酸失衡等均会造成对肝脏的损伤,本试验结果与FSBE在洛氏鱥[32]研究中的结果相似。
机体的防御系统与免疫、抗氧化能力及健康密切相关。T-SOD和CAT在清除氧自由基稳态系统中发挥重要作用,与水产动物免疫能力密切相关。MDA是脂质过氧化后的产物,其含量高低可以用来衡量机体损害严重程度。T-AOC是衡量鱼体抗氧化能力的综合指标。LZM具抗感染、抗炎、增强抗生素效力和提高吞噬活力等功能,是一种重要的非特异免疫因子,为鱼类免疫防御系统重要组成部分。AKP是鱼类生理活动及疾病诊断的重要指标之一,可反映生物有机体抗应激能力。本试验结果发现,除CAT外,FSBE适量替代鱼粉可提高血清中T-AOC、T-SOD、ACP、GSH-Px和LZM活性;而使MDA含量下降。这与刘韬等[3]在凡纳滨对虾、李宁宇等[7]在日本鳗鲡仔鳗的研究结果一致,表明发酵豆粕可增强大口黑鲈抗氧化能力。闫磊等[34]研究发现,当FSBE替代鱼粉比例≥49.5%时,洛氏鱥血清中SOD和LZM活性显著降低,而AKP活性各试验组间差异不显著。本试验中,FSBE替代鱼粉比例≥20%时,导致血清LZM活性显著下降,T-SOD活性显著升高,AKP活性降低,与闫磊等[34]的研究结果相似。Rahimnejad等[9]、Li等[17]及Wang等[23]分别对鲈鱼(Lateolabraxjaponicus)、大口黑鲈和大菱鲆(ScophthalmusmaximusL.)研究发现,发酵豆粕可显著提高血清中LZM活性。而Choi等[8]研究发现,发酵豆粕替代鱼粉可显著提高虹鳟血清AKP活性。上述迥异结果可能与豆粕发酵工艺、受试动物种类、饲料营养及养殖条件等影响其免疫功能作用的调控有关。植物蛋白质原料中的抗营养因子可引起氧化应激,替代鱼粉后会造成非特异性免疫受损[35]。本试验结果中,血清T-SOD活性显著升高表明FSBE替代鱼粉可能会使大口黑鲈处于应激状态中,而FSBE替代鱼粉比例≥20%时,血清LZM活性显著下降表明抗病能力有所减弱。
3.3 FSBE替代鱼粉对大口黑鲈肝脏组织形态的影响
肝脏组织形态是了解肝脏病理情况的一种重要途径。高水平植物蛋白质原料饲喂容易导致鱼发生代谢紊乱、肝脏损伤,认为主要由于植物蛋白质源中含有大量抗营养因子所致。林一帆等[14]报道,含50%发酵豆粕的复合动植物蛋白质源替代饲料20%以上鱼粉时,肝细胞逐渐变大,类似脂滴的空泡现象加剧,细胞轮廓变的模糊,细胞核朝质膜移位。本试验中肝脏细胞空泡化程度随着FSBE替代鱼粉比例增加而加剧,且血管增生并出现炎灶,这可能与豆粕虽经菌酶协同发酵后抗营养因子得到了部分降解,但仍降解不完全有关。因此,FSBE在生产中的有效应用之前,还需从对肝脏生理及病理方面的影响开展进一步研究。
4 结 论
综上所述,本试验用FSBE替代饲料中不超过20%鱼粉,对大口黑鲈生长性能、形体指标、体成分、血清生化指标、免疫和抗氧化指标和肝脏组织形态结构不会产生明显不良的影响,但随着替代比例的进一步升高,特定生长率、免疫和抗氧化能力下降。同等比例替代鱼粉,FSBE效果优于普通豆粕。因此,本试验条件下,FSBE替代饲料(含50%鱼粉)鱼粉比例不超过20%较为适宜。
猜你喜欢
当代水产(2022年8期)2022-09-20
当代水产(2022年6期)2022-06-29
基层中医药(2021年6期)2021-11-02
当代水产(2019年11期)2019-12-23
当代水产(2019年7期)2019-09-03
当代水产(2019年6期)2019-07-25
当代水产(2019年6期)2019-07-25
当代水产(2019年1期)2019-05-16
创新作文(小学版)(2018年10期)2018-08-20
河南畜牧兽医(2017年8期)2017-11-24
