
番茄砧木耐热性评价及热胁迫生理响应研究
2021-06-15 14:29田茂燕向婷颖阳燕娟于文进
中国蔬菜 2021年5期
田茂燕 向婷颖 钟 川 王 鹏 阳燕娟 于文进
(广西大学农学院,广西南宁 530004)
番茄(Solanum lycopersicum)是世界上重要的经济作物,在我国蔬菜产业中占有重要地位,是我国主要的大宗蔬菜之一(毛胜利 等,2001;王冬梅,2003)。番茄原产地位于赤道附近的南美西部高原地带,比较适应高地干燥冷凉的环境条件,是对温度较为敏感的喜温植物,适宜的生长温度为15~32 ℃,过高或过低的温度均会影响其正常的生长发育(安凤霞 等,2005;张富存 等,2011;Wang et al.,2020)。近年来,随着全球温室效应的加剧,气温升高,我国番茄生产面临热胁迫的严峻挑战,特别是南方地区露地番茄主栽季节,春延后栽培和秋提前栽培的番茄易受异常高温的影响,成为限制番茄生产的主要原因之一。研究表明,高温不仅降低番茄种子活力,抑制种子萌发(钱春梅等,2002),而且导致植株体内抗氧化系统失衡,破坏叶绿体膜结构,通过非气孔因素影响光合作用(魏爽 等,2019)。不耐热的番茄幼苗受到35 ℃以上高温热胁迫后,呼吸作用会发生紊乱,且随着胁迫时间的延长,植株茎叶与花器的发育受到严重影响,导致茎叶萎焉与畸形果实的产生,甚至植株茎叶停止伸长直至枯萎,严重影响番茄的营养生长,降低产量和品质;耐热的番茄品种则受热胁迫影响较小,且幼苗的耐热性与采收期的耐热性呈显著正相关(刘祖祺,1994;Li et al.,2011;彭勇政 等,2019)。采用耐热砧木进行嫁接栽培是提高番茄高温耐受性的重要方法。嫁接栽培可以增强番茄的抗逆性和生长势,提高坐果率,提早收获,提高产量与改善品质(范双喜和王绍辉,2005)。利用耐热性较强的砧木嫁接番茄,嫁接苗即使处于高温环境下,植株叶片叶绿素含量、净光合速率都显著高于自根苗,说明嫁接栽培可以显著提高番茄的耐热性(董灵迪 等,2010)。因此,鉴定和筛选耐热番茄砧木种质具有重要的现实意义。本试验通过比较热胁迫对不同番茄砧木种子萌发和幼苗生长的影响,评价番茄砧木种质的耐热性,研究砧木幼苗对热胁迫的生理响应,以期为番茄耐热砧木种质的鉴定和应用提供参考。
1 材料与方法
1.1 试验材料
试验于2018 年11 月至2019 年7 月在广西大学农学院蔬菜基地进行。供试材料为广西大学农学院蔬菜育种课题组保存的20 份番茄砧木种质,其中9 份种质来源于中国,11 份种质来源于日本,均为多代纯合自交系(表1)。

表1 供试番茄砧木种质来源和类型
1.2 番茄砧木种子耐热性鉴定
设置28、31、34、37、40 ℃共5 个温度处理,其中28 ℃为常温对照(CK),进行种子发芽试验,评价砧木种质的耐热性。在直径11 cm 的干净培养皿中垫2 张9 cm 的滤纸,用无菌水润湿后,将20份供试砧木种子(2018 年12 月采种,种子干燥后贮藏于4 ℃冷藏箱内)分别播到滤纸上,每只培养皿均匀播种100 粒,加无菌水保持润湿,种子吸水2 h 后,将加盖的培养皿放入以上5 个不同温度的恒温培养箱内,处理14 d(金春燕 等,2011),期间保持湿润。每份种质、每个温度处理设3 次重复。种子开始发芽后,每天同一时间统计发芽数。
1.3 番茄砧木幼苗耐热性鉴定
将供试砧木种子分别播种到50孔穴盘内育苗,育苗基质按泥炭∶椰糠∶珍珠岩=2V∶1V∶1V的比例配制。当幼苗长至4 片真叶时,转入人工气候箱中,在28 ℃/20 ℃(16 h/8 h)、相对湿度70%~75%、每天光照16 h、光照强度300 μmol · m-2·s-1条件下预培养2 d。第3 天进行热胁迫处理,设置常温对照(28 ℃/20 ℃)、亚高温(34 ℃/28 ℃)、高温(37 ℃/28 ℃)3 个温度处理,每份砧木、每个温度处理幼苗30 株,3 次重复。相对湿度和光照条件同预培养,其他管理条件相同。处理7 d 后调查幼苗热害级数,计算热害指数。采用便携式叶绿素测定仪(SPAD 502)测定植株最上部完全展开叶片的叶绿素SPAD 值。
参考宋敏丽等(2012)的方法,根据幼苗叶片失水萎蔫程度和颜色变化表型,将热害级数分为5 级(图1):植株没有明显高温伤害症状为0 级;植株轻度失水,下部老叶边缘发黄为1 级;植株中度失水,中下部叶片卷曲为2 级;植株严重失水,上部叶片卷曲为3 级;植株多数叶片枯萎死亡为4级。热害指数=∑(各级株数×相应级数)/(最高级数×总株数)。参考马宝鹏(2013)的方法,根据热害指数将苗期耐热性(CI)分为5 级:强耐高温(HR),0 ≤CI ≤0.2;中耐高温(MR),0.2 <CI ≤0.4;耐高温(R),0.4 <CI ≤0.6;高温敏感(S),0.6 <CI ≤0.8;高温高敏感(HS),0.8 <CI ≤1.0。
1.4 热胁迫对番茄砧木幼苗生长及生理响应研究
根据幼苗耐热性鉴定结果,选取强耐高温种质G3、Q2、N3 和高温高敏感种质G2、Y3 进行热胁迫生理响应研究。育苗和热胁迫处理方法同1.3。处理7 d 后,进行植株生长指标和叶片丙二醛(MDA)、脯氨酸含量的测定。用直尺测量植株株高,用游标卡尺测量植株第1 节茎粗(横径),用天平测量地上部、地下部鲜质量和干质量。将植株用自来水冲洗干净,再用去离子水冲洗3 遍,吸水纸吸干表面水分后测定鲜质量;然后置于烘箱105℃杀青15 min,75 ℃恒温烘至恒重后测定干质量。
取植株最上部完全展开叶片,参考李合生等(2000)的方法,采用硫代巴比妥酸法测定叶片MDA含量,采用水合茚三酮法测定叶片脯氨酸含量。
1.5 番茄砧木种质耐热性综合评价
参考但忠等(2013)的方法,运用SPSS 19.0软件对20 份番茄砧木种质进行隶属函数值法分析,综合评价其耐热性。隶属函数值计算公式为R(Xi)=(Xi-Xmin)/(Xmax-Xmin),Xi为某一指标测定值,Xmin、Xmax分别为所有参试材料某一指标的最小值和最大值;如果为负相关,则用反隶属函数进行转换,计算公式为:R(Xi)=1-(Xi-Xmin)/(Xmax-Xmin)。
1.6 统计分析
各指标数据应用Microsoft Excel 2010 软件进行处理和作图,应用SPSS 19.0 统计软件进行方差分析。
2 结果与分析
2.1 热胁迫对番茄砧木种子萌发的影响
从表2 可以看出,常温(28 ℃,CK)条件下,所有参试种质的种子发芽率均在92%以上。31 ℃条件下,有15 份种质发芽率降至80%~88%,5 份种质发芽率降至80%以下,其中Q3 降至69.00%。34 ℃条件下,除N3 的发芽率为64.33%之外,其余种质的发芽率均在50%以下,其中Q3、Q4 的发芽率降至27%左右。37 ℃条件下,除N3、Y2、G3 的发芽率为20%~23%外,其余种质的发芽率均降至20%以下,9 份种质的发芽率为0。40 ℃条件下,所有参试种质的种子发芽率均为0,丧失萌发能力。以上说明,高温对番茄砧木种子萌发具有抑制作用,随着温度升高,种子发芽率明显降低,不同砧木种子萌发受热胁迫的影响不同。
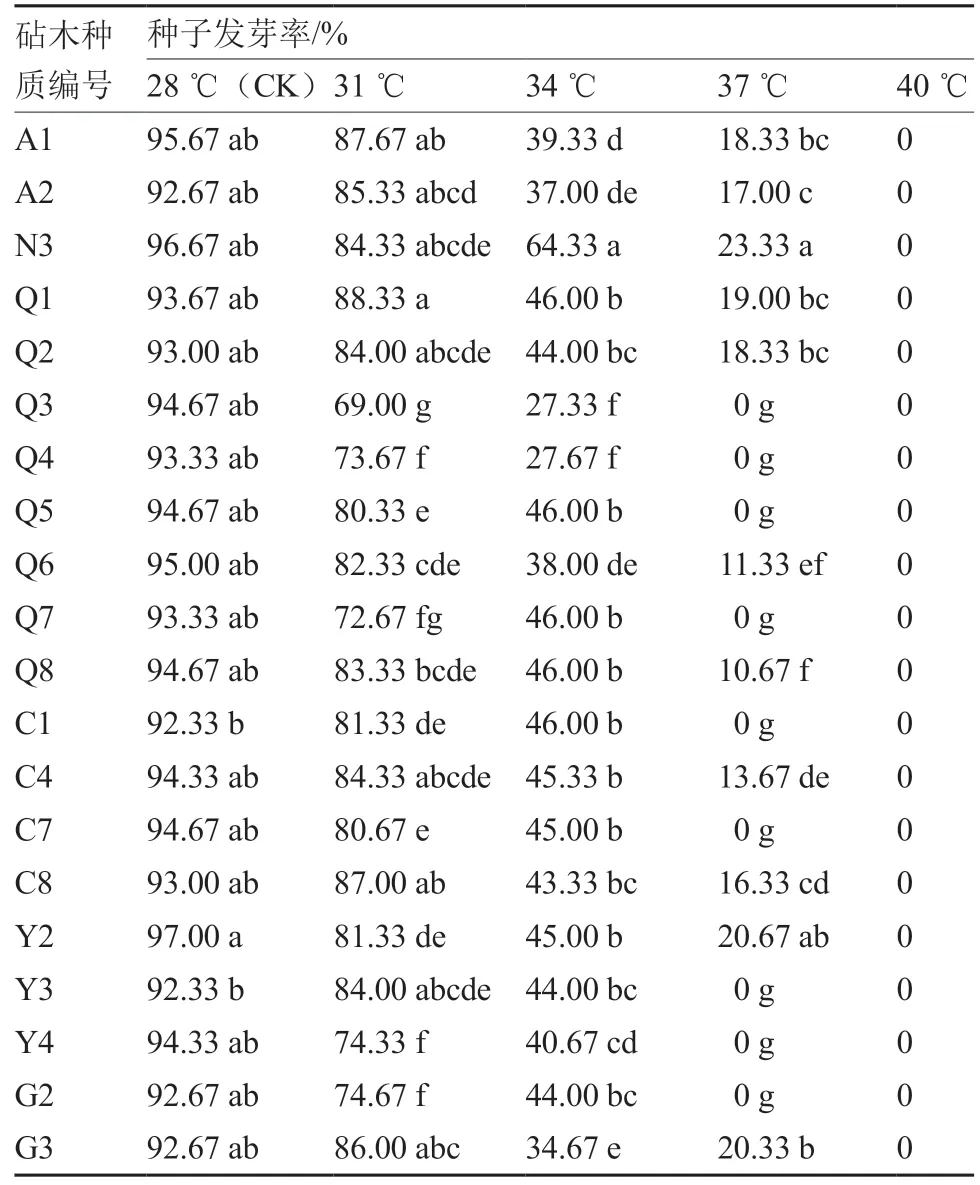
表2 不同温度处理对番茄砧木种子发芽率的影响
2.2 热胁迫对番茄砧木幼苗耐热性的影响
从表3可以看出,亚高温(34 ℃/28 ℃)条件下,有9 份种质的幼苗热害发生率为80%~89%,其余11 份种质的热害发生率为90%~99%;G3、Q2、N3 的热害指数为0.12~0.16,表现强耐高温(HR),Q3、Q5 的热害指数分别为0.39、0.40,表现中耐高 温(MR),A1、Q4、Q7、C1、C7、Y4 等6 份种质的热害指数为0.51~0.60,表现耐高温(R),其余9 份种质的热害指数为0.64~0.80,表现高温敏感(S)。高温(37 ℃/28 ℃)条件下,所有参试种质的幼苗热害发生率均为100.00%;G3、Q2、N3 的热害指数为0.16~0.19,仍然表现强耐高温(HR),A1、Q3、Q4、Q5、Y4 等5 份种质的热害指数为0.55~0.60,表现耐高温(R),其余12 份种质的热害指数为0.71~0.95,表现高温敏感(S)或高温高敏感(HS)。以上说明,番茄砧木幼苗对热胁迫的表型差异明显,种质G3、Q2、N3 较耐高温。
2.3 热胁迫对番茄砧木幼苗叶片叶绿素含量的影响
从表4可以看出,常温(28 ℃/20 ℃,CK)条件下,参试种质的幼苗叶片SPAD 值为42.05~46.57。亚高温(34 ℃/28 ℃)胁迫下,N3、Q2、Q3、Q5、G3 等5 份种质的幼苗叶片SPAD 值比对照降低20%~30%,其余15 份种质的幼苗叶片SPAD 值比对照降低31.13%~36.51%。高温(37 ℃/28 ℃)胁迫下,Q2、Q3、Q5、G3 等4份种质的幼苗叶片SPAD 值比对照降低53.20%~58.94%,A1、N3、C1、Y4 等4 份种质的幼苗叶片SPAD 值比对照降低66.14%~69.66%,其余12 份种质的幼苗叶片SPAD 值比对照降低71.09%~76.40%。以上说明,热胁迫影响番茄砧木幼苗叶片叶绿素合成而使其含量降低,高温比亚高温处理下降幅度大。

表4 热胁迫对番茄砧木幼苗叶片叶绿素含量(SPAD 值)的影响
2.4 热胁迫对番茄砧木幼苗生长的影响
2.4.1 热胁迫对番茄砧木幼苗株高和茎粗的影响 由表5 可知,亚高温(34 ℃/28 ℃)胁迫下,强耐高温种质G3、Q2、N3的株高比对照(28 ℃/20 ℃)增加12.64%~15.49%,高温高敏感种质G2、Y3的株高分别比对照增加23.43%、24.20%;高温(37℃/28 ℃)胁迫下,G3、Q2、N3 的株高比对照增加4.32%~8.09%,G2、Y3 的株高分别比对照增加17.42%、19.44%。亚高温胁迫下,G3、Q2、N3 的茎粗分别比对照减小8.05%~9.58%,G2、Y3 的茎粗分别比对照减小27.51%、23.17%;高温胁迫下,G3、Q2、N3 的茎粗比对照减小26.04%~30.13%,G2、Y3 的茎粗分别比对照减小50.74%、53.13%。以上说明,热胁迫导致番茄砧木幼苗株高增加而茎粗降低,幼苗表现徒长,强耐高温种质G3、Q2、N3 的株高增幅和茎粗降幅比高温高敏感种质G2、Y3 小,幼苗徒长程度相对较小。
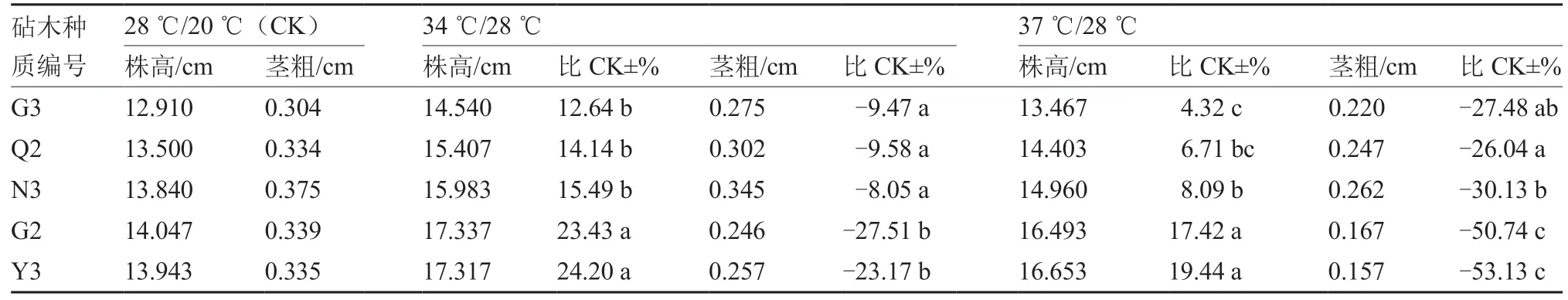
表5 不同温度处理对番茄砧木幼苗株高与茎粗的影响
2.4.2 热胁迫对番茄砧木幼苗鲜质量和干质量的影响 从表6、7 可以看出,亚高温(34 ℃/28 ℃)胁迫下,G3、Q2、N3 的地上部鲜质量比对照(28℃/20 ℃)降低16.10%~18.40%,地下部鲜质量比对照降低23.79%~30.00%;G2、Y3 的地上部鲜质量分别比对照降低38.38%、41.80%,地下部鲜质量分别比对照降低52.08%、56.51%。高温(37℃/28 ℃)胁迫下,G3、Q2、N3 的地上部鲜质量比对照降低30.57%~38.56%,地下部鲜质量比对照降低34.43%~60.74%;G2、Y3 的地上部鲜质量分别比对照降低58.41%、58.86%,地下部鲜质量分别比对照降低75.63%、80.70%。干质量的降幅与鲜质量类似。以上表明,热胁迫导致番茄砧木幼苗的鲜质量和干质量降低,强耐高温种质G3、Q2、N3 的降幅比高温高敏感种质G2、Y3 小,生物量增长受抑制程度相对较小。

表6 不同温度处理对番茄砧木幼苗地上部鲜质量与干质量的影响

表7 不同温度处理对番茄砧木幼苗地下部鲜质量与干质量的影响
2.5 热胁迫对番茄砧木幼苗叶片MDA和游离脯氨酸含量的影响
从图2、3 可以看出,热胁迫导致番茄砧木幼苗叶片的MDA 含量和游离脯氨酸含量增加。亚高温(34 ℃/28 ℃)胁迫下,G3、Q2、N3的叶片MDA含量比对照(28 ℃/20 ℃)升高29.60%~50.10%,G2、Y3分别比对照升高89.99%、129.84%。高温(37 ℃/28 ℃)胁迫下,G3、Q2、N3的叶片MDA含量比对照升高55.41%~89.14%,G2、Y3分别比对照升高120.76%、154.75%。亚高温胁迫下,G3、Q2、N3 的叶片游离脯氨酸含量比对照升高84.34%~125.58%,G2、Y3分别比对照升高8.75%、24.73%。高温胁迫下,G3、Q2、N3 的叶片游离脯氨酸含量比对照升高226.40%~248.70%,G2、Y3比对照分别升高94.83%、130.74%。综合以上结果,强耐高温种质G3、Q2、N3 叶片MDA 含量的增幅比高温高敏感种质G2、Y3 小,说明其因细胞膜脂过氧化引起伤害相对较轻;叶片游离脯氨酸含量增幅大,说明其细胞调节渗透能力相对较强。
2.6 番茄砧木幼苗热害指数与生长生理指标的相关性
分别对亚高温(34 ℃/28 ℃)、高温(37 ℃/28℃)胁迫下番茄砧木幼苗的热害指数及生长生理指标的测定值进行相关性分析。结果显示(表8、9),热害指数分别与株高、叶片MDA 含量呈极显著正相关,与叶片SPAD 值、茎粗、全株鲜质量、全株干质量呈显著负相关,与叶片游离脯氨酸含量呈极显著负相关。以上说明,可用以上7 项生长生理指标对番茄砧木幼苗的耐热性进行综合评价。

表8 亚高温(34 ℃/28 ℃)胁迫下番茄砧木幼苗的热害指数与生长生理指标的相关性
2.7 番茄砧木种质耐热性综合评价
用与热害指数显著相关的7 项生长生理指标进行隶属函数分析,根据隶属函数值的平均值大小对供试种质的耐热性进行排序。结果显示,供试种质在亚高温(34 ℃/28 ℃)和高温(37 ℃/28 ℃)胁迫下的耐热性排序相同(表10、11),强耐高温(HR)种质G3、N3、Q2 排序第1~3 位,与亚高温胁迫的热害指数排序结果一致。以上说明,可以通过幼苗生长生理指标隶属函数分析,对番茄砧木种质的耐热性进行评价。

表9 高温(37 ℃/28 ℃)胁迫下番茄砧木幼苗的热害指数与生长生理指标的相关性

表10 亚高温(34 ℃/28 ℃)胁迫下番茄砧木种质耐热性隶属函数分析

表11 高温(37 ℃/28 ℃)胁迫下番茄砧木种质耐热性隶属函数分析
3 讨论与结论
番茄种子萌发的适宜温度为25~30 ℃,超过30 ℃会抑制番茄种子的萌发,降低种子活力(钱春梅 等,2002)。付丽军等(2017)认为耐热性强的黄瓜种子在高温下仍能保持较高的种子活力和发芽率,出苗更整齐,幼苗健壮。本试验结果表明,番茄砧木种质的种子发芽率随着温度升高而明显降低,34 ℃条件下,大部分种质发芽率降至50%以下,37 ℃条件下的发芽率降至24%以下,40 ℃条件下的发芽率降至0,部分砧木种质对34~37 ℃高温有一定程度的耐受力。
作物在热胁迫下,耐热品种能保持较高的叶绿素含量,不耐热品种的叶绿素合成受阻或降解,叶绿素含量降低,光合作用下降,影响植株正常生长(马晓娣 等,2004;张冉,2017)。植株热害指数是衡量作物受热害程度的常用表观指标,热害指数越大,作物越不耐热(宋敏丽 等,2012;但忠 等,2016)。本试验根据幼苗热害指数,筛选出强耐高温的番茄砧木种质3 份,中耐高温或耐高温的种质5 份。试验结果表明,热胁迫使番茄砧木幼苗叶片叶绿素含量降低,高温比亚高温处理的下降幅度大,强耐高温种质的株高增幅、茎粗降幅和鲜干质量降幅比高温敏感种质小,幼苗徒长和生物量增长受抑制程度相对较小。
植物细胞膜系统对逆境响应敏感,温度过高使细胞膜透性增大,造成超氧自由基等活性氧大量积累(张佳平 等,2016;胡能兵 等,2018;Ozturk et al.,2020),氧化胁迫会引起膜脂发生过氧化和脱脂化作用,膜结构被破坏,使膜脂过氧化产物MDA 含量增加,从而导致植株代谢紊乱(聂庆娟等,2007;崔萌 等,2016;Balyan et al.,2020)。植物在逆境条件下,游离脯氨酸积累,提高了植物对逆境的抗性及适应性,抗逆性强的游离脯氨酸积累量大(欧阳敦君和张鸽香,2016;周桂英 等,2016)。叶片MDA 和游离脯氨酸含量可作为番茄、豇豆的耐热性判定指标(尹贤贵 等,2001)。本试验结果显示,热胁迫导致番茄砧木幼苗叶片的MDA 含量和游离脯氨酸含量增加,强耐高温种质的MDA 含量增幅比高温高敏感种质小,游离脯氨酸含量增幅则比高温高敏感种质大,说明强耐高温种质因细胞膜脂过氧化引起伤害相对较轻,细胞调节渗透能力相对较强。
胡能兵等(2018)在辣椒耐热性上的研究显示,热害指数与植株生长、MDA 含量等指标具有相关性。本试验结果表明,番茄砧木幼苗的热害指数与株高、茎粗、全株鲜质量、全株干质量和叶片SPAD 值、MDA 含量、游离脯氨酸含量等生长生理指标具有显著相关性,可通过隶属函数分析对种质的耐热性强弱进行评价。
综合本试验结果,热胁迫抑制番茄砧木种子萌发,随着温度升高种子发芽率明显降低。34 ℃条件下,除N3 外其余种质的种子发芽率均降至50%以下;37 ℃条件下的种子发芽率均降至24%以下,40 ℃条件下的种子发芽率均为0。亚高温(34℃/28 ℃)和高温(37 ℃/28 ℃)胁迫下,种质G3、N3、Q2 均表现强耐高温(HR),种质A1、Q3、Q4、Q5、Y4 表现中耐高温(MR)或耐高温(R)。幼苗热害指数与株高、叶片MDA 含量呈极显著正相关,与叶片SPAD 值、茎粗、全株鲜质量、全株干质量、叶片游离脯氨酸含量呈显著或极显著负相关。利用以上7 项生长生理指标,通过隶属函数分析对供试种质的耐热性进行综合评价,种质G3、N3、Q2 的耐热性排序居前3 位,与亚高温胁迫下幼苗热害指数排序结果一致。种质G3、N3、Q2 强耐高温,可作为选育耐热番茄砧木的骨干材料。
猜你喜欢
中国果树(2022年5期)2022-06-09
陶瓷学报(2021年4期)2021-10-14
陶瓷学报(2021年4期)2021-10-14
防爆电机(2021年4期)2021-07-28
落叶果树(2021年6期)2021-02-12
新农业(2017年3期)2017-05-17
安徽农学通报(2015年4期)2015-07-30
食品工业科技(2014年23期)2014-03-11
烟台果树(2012年1期)2012-11-03
