
水旱两种栽培模式下海南山栏稻对白叶枯病抗性鉴定与评价
2021-06-29 01:21刘志超黄孟雨翟楠鑫李来一迟会陈妹姑袁潜华
植物保护 2021年3期
关键词:旱作
刘志超 黄孟雨 翟楠鑫 李来一 迟会 陈妹姑 袁潜华
摘要 :为研究山栏稻对白叶枯病抗性,本试验以17个山栏稻品种为材料,在大田与温室大棚中同时种植,于大田中观察山栏稻在水、旱两种栽培模式下对海口当地白叶枯病菌的自然抗性并测产;在温室中检测水、旱两种栽培模式下山栏稻对菲律宾白叶枯病菌小种PXO99(P6)、PX145(P7)的抗性;并以基因功能性标记检测其抗性基因,分析各指标与抗病性的关系。结果表明,大田环境下17个供试品种中有12个品种在水作栽培模式下感病指数高于旱作,有10个品种水作栽培模式下产量比旱作高。温室中用P6接种山栏稻,水作模式下仅有1个品种表现抗病,8个品种感病;旱作模式下2个品种表现抗病,3个品种感病。接种P7,水作时表现抗病和感病的品种数量分别为7个和2个;旱作时表现抗病的品种高达9个,仅1个品种感病。供试品种中有16個品种含有Xa1抗病基因,4个品种含有Xa27抗病基因,所有品种均不含xa13、Xa21抗病基因。因此认为,山栏稻对海口当地白叶枯病菌小种以及菲律宾白叶枯病菌小种都有一定抗性,且抗性受栽培条件的影响,山栏稻旱作时抗性强于水作。
关键词 :山栏稻; 白叶枯病; 抗性基因; 水作; 旱作
中图分类号:
S 474.1
文献标识码: A
DOI: 10.16688/j.zwbh.2020062
Resistance evaluation and molecular identification of Hainan Shanlan upland
rice to bacterial blight under flooding and drying cultivation conditions
LIU Zhichao, HUANG Mengyu, ZHAI Nanxin, LI Laiyi, CHI Hui, CHEN Meigu, YUAN Qianhua*
(College of Tropical Crops, Hainan University, Haikou 570228, China)
Abstract
In order to investigate the resistance of Shanlan upland rice to bacterial blight (BB), 17 varieties of Shanlan upland rice were used as materials and planted in the field and greenhouse at the same time in this study. In the field, the resistance of Shanlan upland rice to Haikou local races of BB was observed and the yield was measured under flooding and drying cultivation conditions. In the greenhouse, the resistance to two Philippine races of BB, PXO99 (P6) and PX145 (P7) was evaluated under flooding and drying cultivation conditions and the functional marker method was used to detect its resistance genes and relationship between various factors and resistance. The results showed that the disease indices of 12 varieties from 17 varieties under flooding cultivation were higher than that under drying cultivation in the field. In addition, there were 10 varieties with higher yield under flooding cultivation than drying cultivation. After inoculated with P6 in the greenhouse, only one variety showed resistance and eight varieties were susceptible under flooding cultivation, while two varieties were resistant and three varieties showed susceptibility under drying cultivation. After inoculated with P7, seven varieties showed resistance and two were susceptible under flooding cultivation, and nine varieties showed resistance and only one was susceptible under drying cultivation. According to the functional marker detection, 16 varieties contained Xa1 or its homologous gene and four varieties contained Xa27 gene. However, all varieties did not contain resistance genes xa13 and Xa21. Therefore, it is believed that Shanlan upland rice had certain resistance to the bacterial blight races of Filipino and Haikou. The resistance was affected by different cultivation conditions and the resistance was stronger under drying cultivation than flooding cultivation.
Key words
Shanlan upland rice; bacterial blight; resistance gene; flooding cultivation; drying cultivation
由水稻黄单胞菌Xanthomonas oryzae pv. oryzae (Xoo)引起的水稻白叶枯病(rice bacterial blight, BB)是世界范围内的细菌病害,在高温、高湿环境下,水稻白叶枯病容易发生,如遇大风暴雨夹带冰雹等天气,叶片受伤严重,病害往往泛滥成灾[1]。白叶枯病菌由受损叶片的伤口侵入,在维管束中定殖繁衍,病斑扩展后叶片开始萎蔫卷曲,影响光合作用,最终严重影響水稻产量[2]。目前认为白叶枯病菌小种和水稻品种的互作关系符合“基因对基因”假说[3],且不同抗性基因间可能会相互影响[4]。至今已鉴定出45个水稻抗白叶枯病基因,部分基因已完成克隆[56]。这些抗病基因只有少数具有广谱抗性,如Xa21和Xa23等,其他基因则由于抗谱较窄或抗性较弱,尚不能在生产中利用[78]。所以培育和种植含有广谱抗病基因或多抗病基因组合型的品种是防治白叶枯病的有效措施。
山栏稻是海南地区旱稻的主要类型[9],大多数属于热带粳稻或偏粳型种质资源[10],具有优良的农艺特性,米质黏糯。目前山栏稻种植一般采用旱作模式,产量较低。本实验室前期研究发现山栏稻在水作栽培时产量相对旱作会显著增加[11],灌溉方式下产量比传统旱作提高了250.96%[12]。笔者调查发现,海南各地旱作山栏稻至今仍有不同程度的白叶枯病存在,水作种植田块也常有白叶枯病发生。目前关于山栏稻在水、旱栽培条件下对白叶枯病抗性变化的相关研究未见报道,本试验根据白叶枯病菌易随水传播的特点以及山栏稻水作栽培可以提高产量的情况,从海南山栏稻中选取了17个高产品种,在大田、温室试验条件下设置水作、旱作两种栽培模式,在大田处理组中观察山栏稻在不同栽培条件下的自然抗性差异与产量差异,在温室处理组中检测山栏稻在不同栽培条件下接种菲律宾白叶枯病菌小种P6、P7的抗性差异,鉴定各品种抗性基因,从栽培方式、叶长宽比、抗病基因等方面分析影响山栏稻对白叶枯病抗性。本研究结果将为寻找山栏稻对白叶枯病有效的抗性基因型及种植时应选择的品种和栽培方式提供参考依据。
1 材料与方法
1.1 试验材料
山栏稻品种:‘LS134‘LS181‘Q18‘ZO7‘WMS‘LS11‘昌山糯‘昌山麻糯‘海大红‘海东1‘海东2‘琼中白15‘黏稻白沙‘白沙团‘白沙微红‘白沙旱1‘白沙红2,来自海南各县市种植品种及本实验室选育的新品系。
对照组水稻:感病品种‘IR24,含有单个抗病基因的品种‘IRBB1‘IRBB13‘IRBB21‘IRBB27。
白叶枯病菌:菲律宾白叶枯小种PXO99 (P6)、PXO145 (P7)。
对照组水稻及白叶枯病菌由海南大学夏志辉老师实验室提供。
1.2 试验方法
1.2.1 试验材料种植
所有供试品种在大田、温室大棚内都设置水、旱两种栽培条件,育苗后移栽。大田组每个品种水、旱栽培条件下各种植1 m2,行株距20 cm×20 cm,不同品种间相隔40 cm。不喷洒防治白叶枯病的药剂。水作处理组保持10 cm左右水深;旱作处理组三叶一心期后不再进行人工浇水,靠雨水灌溉,设置排水管道防止田间积水,其余管理方式与常规农作方式一致[13]。温室组种植于海南大学水稻资源圃大棚内,每个品种水、旱栽培条件下各种4盆,每盆3穴,水作处理组保持水深10 cm左右;旱作处理组三叶一心期后约每5 d浇水1次,并及时排出积水。
1.2.2 接种及鉴定标准
按照国际水稻研究所《水稻标准化评价体系》[14],以病斑面积占该叶片总面积的比值作为病叶的感病等级评定依据,比值<1%时等级为0,比值在1%~5%时为1级,6%~12%时为3级,13%~25%时为5级,26%~50%时为7级,51%~100%为9级,0级抗性最强,9级最弱。本试验中抗病、感病分类标准为:0级≤感病等级≤3级为抗病(R),3级<感病等级≤6级为中抗(MR),6级<感病等级≤9级为感病(S)[4]。
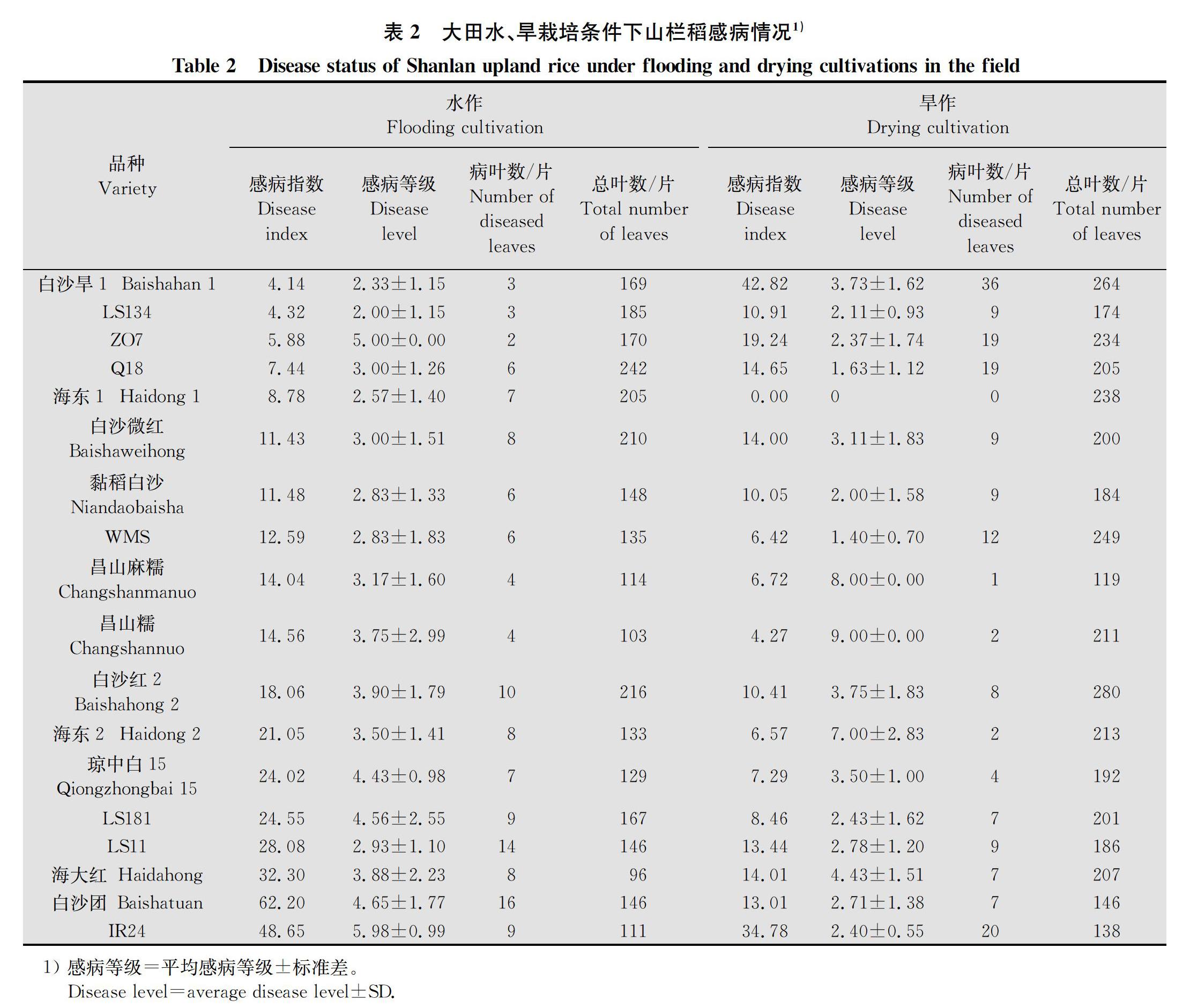
大田组不进行人工接种,采用五点取样法调查山栏稻自然感病指数。每品种调查5穴,在分蘖期统计5穴山栏稻的总叶数、总病叶数及病叶感病等级,计算病叶平均感病等级和感病指数。病叶平均感病等级=∑病叶感病等级/总病叶数,感病指数=(总病叶数×病叶平均感病等级)/总叶数×100。以感病指数表示该品种的自然抗性,感病指数越低,自然抗性越强。
温室组采用剪叶法在分蘖初期接种。白叶枯病菌菌株用PSA 培养基进行扩大培养,制成浓度为1×109 cfu/mL的菌液[1314],将每穴顶部3~5片叶用消毒后的剪刀蘸取菌液剪去叶尖端2~3 cm,每个品种水、旱栽培模式P6、P7菌株各接种3穴,并分别做好标记。接种20 d后,调查每个品种接种P6、P7后的病叶数及感病等级,以各自叶片的平均感病等级分别表示该品种对P6、P7的抗性,感病等级越低,抗性越强。
1.2.3 叶长宽比测量
温室组测量每品种接种叶片的长、宽比值,以平均值表示该品种的叶长宽比。用R语言分析叶长宽比与感病等级的相关性。
1.2.4 产量测定
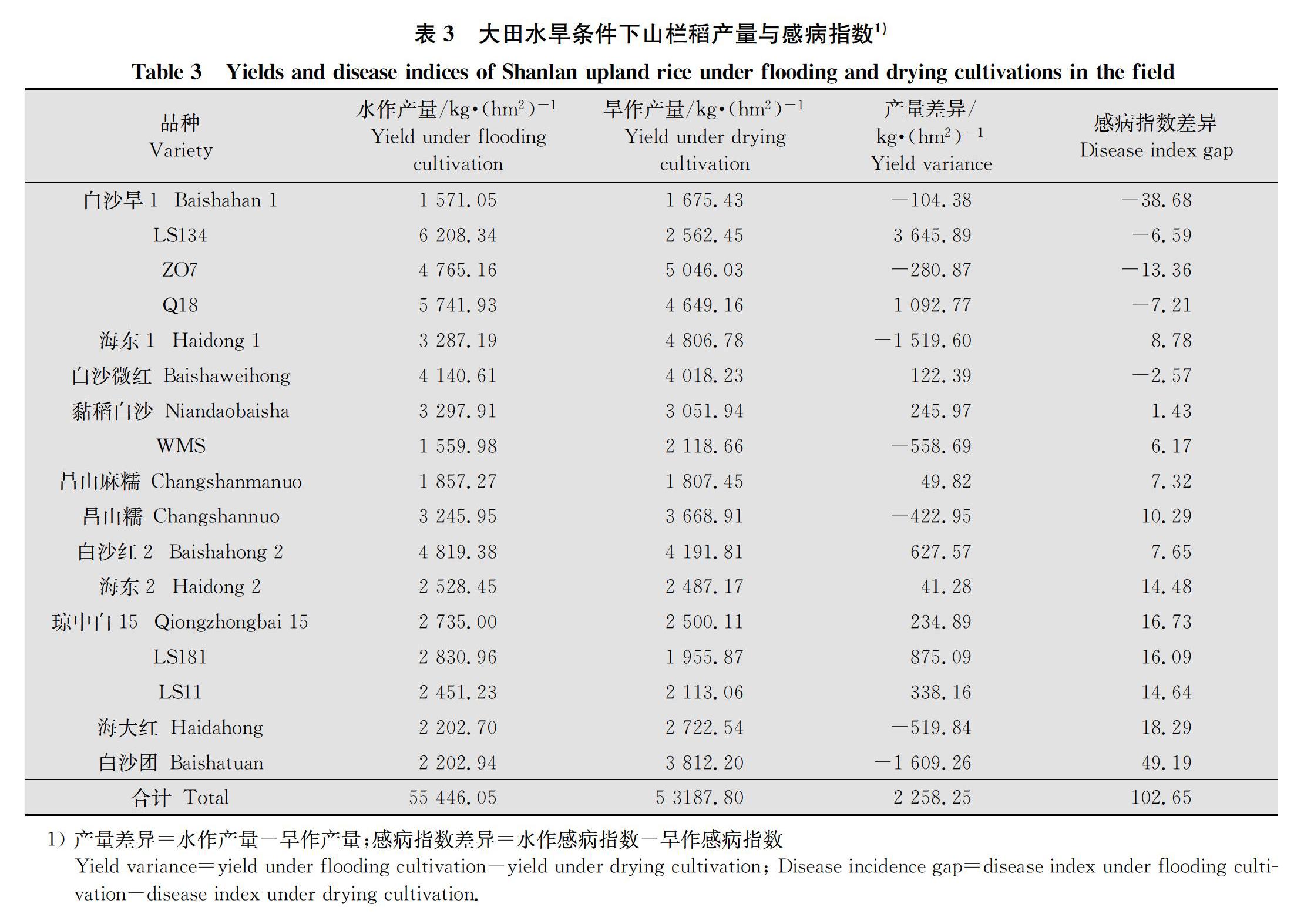
大田处理组水稻成熟后每品种单独收割测产,分析水、旱栽培模式下产量差异。
1.2.5 抗性基因鉴定
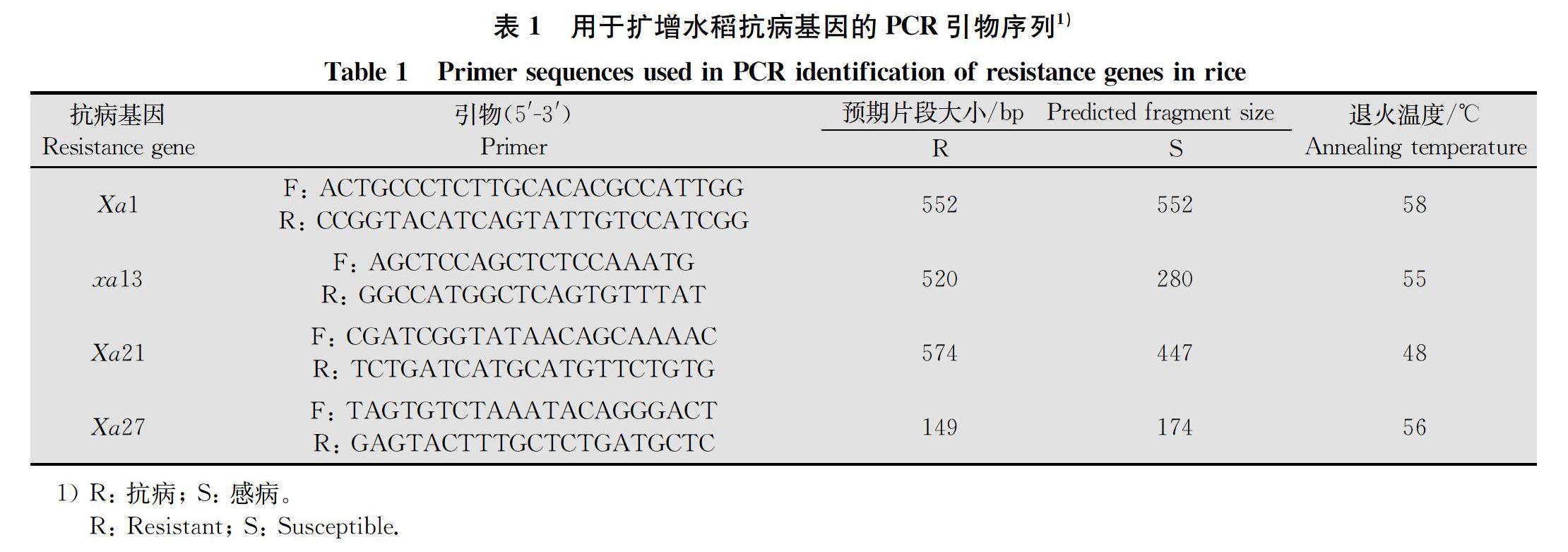
为避免单株间可能存在的遗传差异,每个品种取多个单株叶片混合,用联科生物MKF0104-050型试剂盒提取基因组DNA。在GenBank中查询Xa1、Xa21、Xa27和xa13的基因序列,根据基因序列中多态性位点来设计引物[1516],引物序列见表1。以基因组DNA为模板,进行PCR检测,PCR产物送赛维尔公司测序,测序结果由DNAMAN软件进行多序列比对。
1.3 数据统计与分析
采用Excel 2016统计数据,用R语言进行相关性分析,用DNAMAN进行多序列比对。
2 结果与分析
2.1 大田水、旱栽培对山栏稻白叶枯病抗性及产量的影响
2.1.1 水、旱栽培模式下山栏稻抗性差异分析
山栏稻在不同栽培条件下对当地白叶枯病菌抗性不同。由表2数据可知,17个供试品种中有12个品种水作时感病指数高于旱作,占品种总数的70.59%,代表品种有‘白沙团‘海大红等,其中‘白沙团在水作时感病指数为62.20,旱作时仅为13.01,前者是后者的4.78倍;‘海大红感病指数在旱作时为14.01,水作时达到32.30,是旱作感病指数的2.31倍。‘白沙旱1‘ZO7等5个品种在大田水作时抗性较强,而旱作时感病指数大幅上升,其中‘ZO7水作时感病指数为5.88,旱作时感病指数为19.24,‘白沙旱1在两种栽培模式下感病指数差异更为明显,由水作时的4.14变为旱作时的42.82,提高了9.34倍。综合抗性最好的品种是‘海东1,水作时感病指数为8.78,旱作时未发现病叶。整体而言,大部分山栏稻品种在水作时的自然抗性较差,易被试验区自然界中的白叶枯病菌侵染。
因感病指数与单叶感病等级、病叶数及总叶数都有关,对比相同模式下的不同品种可以发现,即使感病指数接近,各个指标所占的比重也可能不同,或是感病指数不同,但其中某个指标相同。如旱作时‘Q18和‘海大红感病指数接近,分别为14.65和14.01,但病叶数分别为19片和7片,前者是后者的2.71倍,病叶平均感病等级分别为1.63和4.43,后者是前者的2.72倍;水作时‘白沙微红和‘海大红则是病叶数相同,病叶平均感病等级分别为3.00和3.88,而总叶数前者是后者的2.19倍,最终造成后者感病指数比前者高1.83倍。相同品种在不同栽培条件也会由于各个指标不同,造成感病指数的差异,如‘白沙旱1,水作时有3片病叶,旱作时有36片病叶,相差12倍,且病叶感病等级也高于水作,是造成感病指数相差9.34倍的主要指标。由图1相关性分析结果可知,水作感病指数与叶片平均感病等级呈显著正相关,平均等级越高,感病指数越高,与旱作时感病指数差异越明显。没有与旱作感病指数直接相关的指标。
2.1.2 水、旱栽培模式下山栏稻产量与感病指数的关系
分析表3可知,水作产量高的品种,旱作产量一般也高,且呈显著正相关(图1);将同一品种不同栽培模式下的产量差异与自然感病指数差异结合分析,发现二者并无显著的相关关系。17个品种中有8个品种产量与感病指数相关而与栽培模式没有明显相关性,感病指数低时产量高,如品种‘LS134在水作时感病指数较低,旱作时感病指数是水作的2.53倍,相应水作产量是旱作产量的2.42倍。品种‘海东1在旱作时感病指数为0,水作时感病指数为8.78,相应旱作时产量是水作时的1.46倍。有9个品种在某种栽培模式下感病指数虽然相对另一种栽培模式较高,但产量并未下降,如‘海东2在水、旱模式下感病指数分别为21.05和6.57,产量分别为2 528.45 kg/hm2和2 487.17 kg/hm2,即前者感病指数是后者的3.2倍的前提下,产量仍比后者高1.66%;‘LS181水作时感病指数是旱作时的2.9倍。产量却是旱作时的1.45倍。供试材料中有8个品种水作时产量和感病指数同时大于旱作,说明水作时虽然感病严重,但水作时的其他某些增产因素弥补了白叶枯病害对山栏稻的影响,最终提高了产量。仅有‘白沙旱1和‘ZO7两个品种旱作时产量和感病指数均大于水作,即受到病情危害时,旱作模式很难对产量起到弥补作用。
2.2 山栏稻对菲律宾白叶枯病菌小种抗性
温室接种20 d后,各品种抗性反应见表4。统计数据可以发现,山栏稻对P6小种的抗性较弱,而对P7小种的抗性相对较强,整体旱作抗性强于水作。水作模式下接种P6,17个品种仅有‘WMS表现出抗性,另外8个品种表现感病,8个品种表现中抗,‘白沙微红的感病等级高达7.13;接种P7条件下,抗性品种数量明显多于接种P6,7个品种表现抗性,仅有2个品种‘白沙红2和‘白沙微红感病,且等级分别为7和6.67,低于接种P6的感病等级,另外8个品种为中抗。旱作模式下接种P6,仅有3个品种感病,是水作条件下表现感病的品种数量的37.5%,12个品种中抗,其余2个品种为抗性;接种P7时仅1个品种感病,另外7个品种为中抗,9个品种为抗性。
综合抗性较强的品种有‘WMS和‘LS134,‘WMS在旱作接种P6的处理模式下表现中抗,感病等级为3.29,水作接种P6以及水作、旱作接种P7后的3种模式下都表现出较强抗性。‘LS134在水作接种P6时表现中抗,感病等级为3.11,已接近抗性水平,其他3种处理模式均表现出抗性。抗性较弱的品种有‘白沙红2,仅在旱作接种P6时为中抗,其他处理均为感病。不同栽培条件下对相同菌种抗性差异较大的品种有‘海大红和‘海东2等,‘海大红水作条件下对P6表现为感病,感病等级(6.33±0.94),在旱作条件下表现中抗,感病等级(3.50±1.00);‘海东2在水旱条件下对P7的抗性反应分别为中抗(5.00±2.66)和抗性(1.00±0.50)。
分析山栏稻叶长宽比与各处理模式下的感病指数可知,叶长宽比仅与旱作接种P7后的抗性呈显著正相关,即叶片越细长的品种,在旱作栽培时对P7的接种抗性越强。而叶长宽比与水作条件下对P7的抗性以及水、旱两种模式下接种P6后的抗性都不相关。接种抗性与各指标的相关性如图2。
2.3 山栏稻抗病基因PCR檢测结果
同一品种不同单株之间抗性差异较大,说明该品种间存在较大的遗传差异,杂合程度较高,本试验采用功能基因标记方法鉴定了供试品种的抗性基因,图3和图4为鉴定结果。
對Xa1的鉴定结果显示,除了‘昌山麻糯外,其他品种均能扩增出与阳性对照IRBB1相同的条带,约为552 bp。然而‘昌山麻糯对P6和P7的抗性都较强,推测此品种含有除Xa1、xa13、Xa21和Xa27外的其他抗性基因。将Xa1 PCR产物进行测序,与GenBank中公布的Xa1序列进行比对,发现只有个别碱基不同,部分品种序列则完全相同,说明这些材料含有抗性基因Xa1或其同源基因。
对xa13、Xa21的检测结果表明,阳性对照IRBB13和IRBB21的条带大小分别为520 bp和574 bp,所有检测品种与阳性对照的条带位置都不同,且都扩增出了与IR24一致的条带,长度约为280 bp和447 bp,推断检测品种中均不含有抗性基因xa13、Xa21,而是含有与其对应的感病基因Xa13、xa21。
Xa27的鉴定结果表明‘白沙旱1‘LS134‘ZO7和‘WMS这4个品种含有该抗病基因,长度约149 bp,与IRBB27的条带一致,推测该4个品种含有抗性基因Xa27。
3 讨论
整体分析大田数据可知,水作时虽然比旱作感病严重,但最终产量不比旱作低。柯智等[12]认为山栏稻水作或滴灌栽培方式与传统旱作相比,能显著提高产量相关要素,如剑叶净光合速率、结实率和千粒重等,最终提高产量。而本试验水作、旱作产量相差不明显,可能是因为前人试验中做了防止白叶枯病危害的措施,排除了白叶枯病对水作产量的影响。吴鸿健等[17]在研究水稻抗稻瘟病时提出“感病品种在非病害流行期,往往有高产、优质等良好表现”,针对本研究中产量随抗病性变化的两个品种‘海东1‘白沙团分析,可推测在控制病害的情况下,两品种产量会有所提高。
部分品种在水、旱两种栽培条件下对白叶枯病菌的抗性变化有所不同,如‘LS11在温室中对P6小种的抗性为水作强于旱作,而在大田中对于试验区优势菌种的抗性则是旱作强于水作,推测是因为大田中环境复杂,同一植株可能被不同菌种侵染,且侵染植株的菌种与温室不同,因而其抗性变化有差异[18]。此结果提醒我们在进行品种推广前应对当地菌种类型及致病机理开展相应调查和研究。
从分子角度讲,并非抗性基因越多的品种抗性越强,含有多个抗性基因的品种,基因在表达时可能会相互影响,将xa5、xa13和Xa21整合到品种‘PR106后,会显著增加其对多个菲律宾小种的抗性[18];‘IRBB52品种中同时含有Xa4和Xa21,但是它对某印度白叶枯病菌小种的抗性不如只含有Xa21的品种‘IRBB21[19]。本试验中‘白沙旱1和‘LS134,已验证都含有抗性基因Xa1,但是对于P6小种的抗性差异很大,所以推测这两个品种至少有一个品种含有除xa13、Xa21和Xa27之外的其他抗性相关基因,并且此基因可能影响Xa1的表达,或对P6小种有抗性。Amantebordeos 等[20]认为Xa27可以高抗P6,本试验中4个含有该基因的山栏稻品种中,仅有‘WMS和‘LS1342个品种表现出较强的抗性,其他2个品种对P6抗性较差,推测其他2个品种中,Xa27基因可能受到某种因素抑制而无法正常表达。所有供试品种对白叶枯病菌小种都表现出一定程度的抗性,说明其中存在抗性基因,且不同单株间的抗性差异较大,说明供试品种的遗传多样性较高,可能含有新的抗性基因[21],这些基因及其表达方式都有较大的科研价值,这有待于我们进一步研究。
本研究结果表明,山栏稻对海口当地白叶枯病小种以及菲律宾小种都有一定抗性。水作模式下山栏稻对白叶枯病抗性相对低于旱作模式,但两种模式总产量相差不大。综合分析认为,山栏稻对白叶枯病的抗性受栽培条件与致病菌种的影响。在白叶枯病情受到控制的前提下,相对于旱作,大部分山栏稻品种可通过采用水作栽培模式提高其产量。
参考文献
[1] 白文周. 浅析水稻白叶枯病的发生及防治[J]. 云南农业, 2017(7): 4243.
[2] 种藏文, 肖锋, 王长方, 等. 福建省水稻白叶枯病和细条病比较研究[J]. 福建农业科技, 1998(6):14.
[3] 莫伟兰, 张子宇, 赵帅, 等. 水稻白叶枯病菌基因XOO2193的突变体构建及其毒力和胞外多糖分析[J]. 基因组学与应用生物学, 2011, 30(5): 564570.
[4] AMRITA B, SOMNATH R, KUMAR B M, et al. A survey of bacterial blight (Xanthomonas oryzae pv. oryzae) resistance in rice germplasm from eastern and northeastern India using molecular markers [J]. Crop Protection, 2018, 112: 168176.
[5] HUTIN M, SABOT F, GHESQUIRE A K, et al. A knowledge-based molecular screen uncovers a broad-spectrum, OsSWEET14, resistance allele to bacterial blight from wild rice [J]. The Plant Journal, 2015, 84(4):694703.
[6] NEELAM K, MAHAJAN R, GUPTA V, et al. High-resolution genetic mapping of a novel bacterial blight resistance gene xa-45(t) identified from Oryza glaberrima and transferred to Oryza sativa [J]. Theoretical and Applied Genetics, 2020, 133(3): 689705.
[7] SONG Wenyuan, WANG Guoliang, CHEN Lili, et al. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21 [J]. Science, 1995, 270(5243): 18041806.
[8] WANG Chunlian, ZHANG Xiaoping, FAN Yinglun, et al. Xa23 is an executor R protein and confers broad-spectrum disease resistance in rice [J]. Molecular Plant, 2015, 8(2): 290302.
[9] 王官远, 陈泰琼, 黄丹. 海南岛作物种质资源考察文集[M]. 北京: 农业出版社, 1992: 1319.
[10]郑成木, 黄东益, 陈辉, 等. 海南山栏稻种质类缘与杂交育种利用的研究[J]. 热带作物学报, 1998,19(2): 7481.
[11]吴丹, 吴川德, 何美丹, 等. 水作和旱作对山栏稻生长的影响[J]. 热带生物学报, 2017, 8(3): 318323.
[12]柯智, 黃孟雨, 刘志超, 等. 栽培方式对山栏稻光合作用和产量的影响[J]. 热带生物学报, 2019, 10(4): 331337.
[13]向妙莲, 蒋海燕, 曾晓春, 等. 茉莉酸甲酯诱导大田水稻抗白叶枯病的效应研究[J]. 植物保护, 2016, 42(2): 9598.
[14]International Rice Research Institute. Standard evaluation system for rice [DB/OL]. 5th Edition. IRRI PO Box 933, 1099 Manila, Philippines. http:∥inger.irri.org.
[15]夏志辉, 韩飞, 高利芬,等. 利用功能标记鉴定普通野生稻中的白叶枯病抗性基因[J]. 中国水稻科学, 2009, 23(6): 653656.
[16]赵天龙, 范玉龙, 王艺洁, 等. 白叶枯病抗病基因Xa21与Xa23的聚合育种研究[J]. 分子植物育种,2015, 13(3): 513517.
[17]吴鸿健, 潘湘濒, 朱荣生, 等. 水稻穗颈段组织结构对稻瘟病抗病性的研究[J]. 植物保护, 1989, 15(3): 24.
[18]夏立琼, 李明容, 谢仕猛, 等. 海南水稻白叶枯病菌优势生理小种的分离及致病力分析[J]. 分子植物育种, 2016(5):13361340.
[19]SINGH S, SIDHU J S, HUANG N, et al. Pyramiding three bacterial blight resistance genes (xa5, xa13 and Xa21) using marker-assisted selection into indica rice cultivar PR106 [J]. Theoretical and Applied Genetics, 2001, 102(6/7):10111015.
[20]AMANTEBORDEOS A, SITCH L A, NELSON R, et al. Transfer of bacterial blight and blast resistance from the tetraploid wild rice Oryza minuta to cultivated rice, Oryza sativa [J]. Theoretical and Applied Genetics, 1992, 84(3/4): 345354.
[21]李栋, 何美丹, 吴丹, 等. 海南普通野生稻对水稻白叶枯病的抗性鉴定[J]. 分子植物育种, 2018, 16(3): 832839.
(责任编辑:田 喆)
猜你喜欢
科学导报(2018年16期)2018-05-14
农家科技下旬刊(2017年8期)2017-11-13
吉林农业·下半月(2017年11期)2017-11-11
农业工程技术·综合版(2016年9期)2017-04-23
农民致富之友(2017年4期)2017-04-10
农业工程技术·综合版(2016年3期)2016-07-27
世纪之星·交流版(2016年3期)2016-05-19
天津农业科学(2015年8期)2015-08-06
农业科技与装备(2014年2期)2014-11-28
现代农业科技(2009年17期)2009-03-08
