
木贼镰孢基因功能注释及比较基因组学分析
2021-06-30 03:18李雪萍许世洋范雨轩汪学苗张怡忻李敏权
微生物学杂志 2021年1期
李雪萍, 许世洋, 范雨轩, 汪学苗, 张怡忻, 李敏权*
(1.甘肃省农业科学院 植物保护研究所,甘肃 兰州 730070;2.甘肃农业大学 草业学院,甘肃 兰州 730070)
木贼镰孢(Fusariumequiseti(Corda)Sacc.)是一种功能复杂的真菌,有性时期为错综赤霉(GibberellaintricansWollenw.),其寄主广泛,致病性强,是多种植物的病原[1-3]。近年来其危害逐渐加重,能侵染小麦、玉米、大麦、燕麦、黑小麦等多种作物,引起小麦冠腐病、根腐病和茎基腐病[4],玉米鞘腐病[5]、穗腐病[6],水稻立枯病[7]等。木贼镰孢也能侵染马铃薯、芦笋、豇豆、番茄、洋葱、豌豆、蚕豆、鹰嘴豆、大头菜、布什豆、油菜、甜瓜等蔬菜瓜果类作物[8],导致豇豆、大豆根腐病[9-10]、莴苣叶斑病[11]、甜瓜枯萎病[12]、向日葵枯萎病[13]、青枣腐烂病[14]及萝卜苗期叶部病害[15]等。此外,木贼镰孢还能引起诸如小茴香枯萎病[16]、五味子茎基腐病[17]、鱼尾葵褐斑病[18]、桑树桑疫病[19]、大花蕙兰及苜蓿根腐病[20-21]等一些花草树木及中药材的病害。木贼镰孢之所以能使多种不同作物感病,多是因为其可以产生一些毒素[22]。而这些毒素的产生、调控机制及如何危害植物,都需要找出具体的原因。ukasz等[23]通过分析木贼镰孢tef-1α序列产生毒素的基因,发现来自ZEA合成途径的2个聚酮体合成酶的基因PKS13和PKS4,以及来自单端孢霉烯合成途径的TRI5基因。但仅进行小片段的序列分析难以找出更多相关的调控基因。目前,全基因组测序技术已经发展成熟,如通过全基因组测序,发现了大肠埃希菌(E.coli)O104∶H4编码志贺毒素的stx2基因[24]、猪链球菌基因组中存在一个89 kb的毒力岛[25]、Ⅱ型糖尿病人易感基因[26]、3个与大肠埃希菌细胞分裂直接相关的基因[27]、苹果滇楸叶斑病的抗性基因[28]等。由此可见,通过全基因组测序能深入理解病原的作用机制、耐药基因和致病基因的传播规律、植物与病原的互作机制等,从而解决病害给农业生产带来的各种问题。前期研究中,基于Illumina Hiseq 4000和PacBio平台对木贼镰孢D25-1的全基因组进行从头测序、组装分析、提交至NCBI,Illumina平台的测序结果在DDBJ/ENA/GenBank数据库的注册号为QJGT00000000,PacBio平台的组装结果在DDBJ/ENA/GenBank数据库的注册号为QOHM00000000。在此基础上,本研究对其基因功能进行全面的注释,找出产毒基因,并进行比较基因组学分析,为木贼镰孢甚至镰孢菌属功能基因的挖掘以及致病机理的研究、抗生素药物开发和相应病害控制策略的建立提供切实有效的参考。
1 材料与方法
1.1 材料
1.1.1 基因组基本信息 木贼镰孢D25-1基因组大小为40 776 005 bp,组装得到16个Contig,最长Contig为8 344 890 bp,最短Contig为2 316 bp,GC含量为48.01%,见表1。在NCBI中选取能引起根腐病的外属病原麦根腐平脐蠕孢(Bipolarissorokiniana),同属病原燕麦镰孢(Fusariumavenaceum)、尖孢镰孢(Fusariumoxysporum)、假禾谷镰孢(Fusariumpseudograminearum)、茄病镰孢(Nectriahaematococca)5 个菌株作为参考,与木贼镰孢D25-1的全基因组序列进行比较分析,基本信息如表2所示。

表1 木贼镰孢D25-1的组装信息
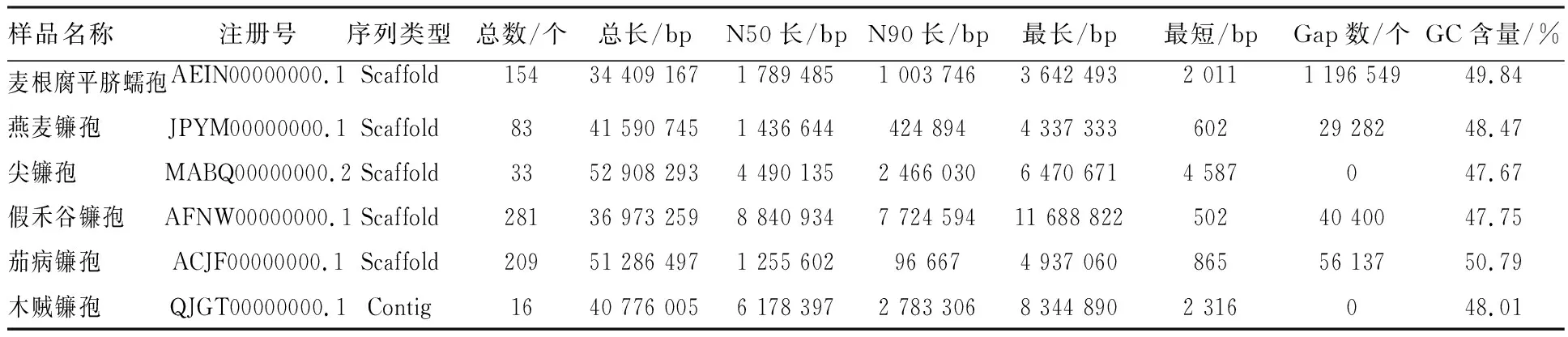
表2 基因组基本信息
1.1.2 培养基 马铃薯葡萄糖培养液(g/L):马铃薯200,葡萄糖20,蒸馏水1 000 mL。
1.1.3 试剂与仪器 50 g/mL玉米赤霉烯酮标准品(Sigma-Aldrich,CAS:17924-92-4);恒温振荡培养箱(ZHLY-50,上海知楚仪器有限公司);超声波细胞破碎仪(Biosafer900-92,Biosafer);液相色谱仪(Agilent 1200,Agilent)。
1.2 方法
1.2.1 基因功能注释 基于氨基酸序列进行基因功能注释。使用BLAST软件将木贼镰孢D25-1待测基因对应的氨基酸序列与Gene Ontology (GO)[29]、Kyoto Encyclopedia of Genes and Genomes(KEGG)[30]、Cluster of Orthologous Groups of proteins(COG)[31]、EggNOG[32]、NR、Antibiotic Resistance Genes Database(ARDB)、Carbohydrate-Active enZYmes Database (CAZy)等数据库进行比对,得到相应的功能注释信息。
1.2.2 产毒基因进化分析 镰孢属可产生多种毒素,主要包括玉米赤霉烯酮、单端孢霉毒素、串珠镰刀菌素和伏马菌素等[33]。基于1.2.1基因注释结果,从中寻找产毒相关基因,将其序列与GenBank中收录的序列进行比对,运用Mega1.7.9构建基因的系统进化树。
1.2.3 玉米赤霉烯酮分泌规律测定 将木贼镰孢D25-1接种于马铃薯葡萄糖培养液中,25 ℃,120 r/min摇瓶培养,根据前期预实验,从第5天起每天定时抽取培养液,HPLC法检测其玉米赤霉烯酮含量,检测方法依据国标GB5009.209-2016[34]。色谱条件:①色谱柱:C18柱,柱长150 mm,内径4.6 mm,粒度4 μm;②流动相:V(乙睛)∶V(水)∶V(甲醇)=46∶46∶8;③流速:1.0 mL/min;④波长:检测波长274 nm,发射波长440 nm;⑤进样量:100 μL;⑥柱温:室温。
1.2.4 比较基因组学分析 首先取参考菌株的基因集为Reference基因集,选取目标菌株Query基因集与Reference基因集进行BLAST比对;根据比对的长度与Identity值过滤比对结果,然后分别计算Reference与Query中每个基因的覆盖比率BCR值(The Blast Coverage Ratio)。Reference与Query基因的BCR值计算公式[35]:BCR(Ref)=(Match/Length(R))×100%;BCR(Que)=(Match/Length(Q))×100%;其中Match为二者比对有效长度;Length (R)为Reference基因长度;Length (Q)为Query基因长度。最后根据设定的阈值确定泛基因(Pan基因)和共有基因(Core基因)。使用Treebest 1.9.2的PHYML(最大似然法)算法构建系统进化树,bootstraps参数设置为1 000。
2 结果与分析
2.1 基因功能注释
由表3可知,通过与各数据库BLAST比对分析,被注释的基因达13 134个,占总基因数的94.97%。其中NR数据库、NOG数据库、IPR数据库的注释条数较多为70%以上;病原与宿主互作数据库(PHI)注释到1 422个基因,占10.28%。

表3 总体注释情况
如图1所示,通过KEGG Pathway数据库对木贼镰孢D25-1基因组的注释结果表明,其基因功能总体可分为6类,包括细胞过程、环境信息处理、基因信息处理、人类疾病、新陈代谢和有机系统。其中与细胞过程有关的功能基因有4种,共590条;与环境信息处理有关的功能基因有3种,共310条;与基因信息处理有关的功能基因有4种,共861条;与人类疾病有关的功能基因有12种,共791条;与新陈代谢有关的功能基因有12种,共3 781条;与有机系统有关的功能基因有10种,共569种。结果表明,与新陈代谢有关的功能基因的种类和丰度最高。

图1 KEGG功能注释分类Fig.1 KEGG annotations classification
2.2 产毒基因分析
基于2.1的注释结果对产毒基因进行挖掘。发现2个与玉米赤霉烯酮相关的基因,分别为D25-1_GLEAN_10000531和D25-1_GLEAN_10000533。D25-1_GLEAN_10000531与玉米赤霉烯酮合成过程中非还原迭代I型聚酮合成酶相关;D25-1_GLEAN_10000533与玉米赤霉烯酮合成过程中高度还原迭代I型聚酮合成酶相关(表4)。

表4 与玉米赤霉烯酮有关的基因
2.3 玉米赤霉烯酮合成酶基因进化分析
将木贼镰孢D25-1基因组中与分泌玉米赤霉烯酮有关的功能基因D25-1_GLEAN_10000531和D25-1_GLEAN_10000533分别命名为ALM1和ALM2。将其序列提交至NCBI获得的登录号分别为ALM1(MT038378)和ALM2(MT038379)。如图2所示,构建系统发育树进化分析发现,ALM1与玉米赤霉烯酮合成过程中非还原迭代I型聚酮合成酶的遗传距离为0,自展支持率为100;ALM2基因与玉米赤霉烯酮合成过程中高度还原迭代I型聚酮合成酶的遗传距离为0,自展支持率为100;均确定为玉米赤霉烯酮合成酶基因。

图2 木贼镰孢D25-1 ALM1和ALM2基因的系统进化树Fig.2 Phylogenetic tree of Fusarium equiseti D25-1 ALM1 and ALM2 genes
2.4 玉米赤霉烯酮分泌规律
如图3 HPLC检测结果所示,在木贼镰孢D25-1的代谢产物中检测到玉米赤霉烯酮,培养第5~8天时玉米赤霉烯酮分泌量较大,呈指数增长。从第9天开始,玉米赤霉烯酮的含量达到40 μg/L以上,并趋于平稳。
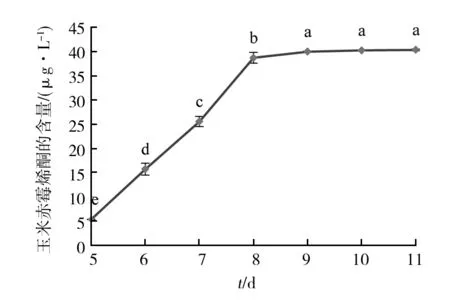
图3 木贼镰孢D25-1分泌玉米赤霉烯酮规律Fig.3 Regulation of Zearalenone secretion by Fusarium equiseti D25-1a、b、c、d、e表示在5%水平上差异显著性,字母不同表示差异显著a,b,c,d,e represents the significance of the difference at the 5% level,different letters indicate significant differences
2.5 比较基因组学分析
6株菌的Core基因共有7 378个,总长为3 821 847个碱基。Pan基因24 332个,总长为9 094 975个碱基,是Core基因和非必需基因的总和(表5)。随着菌株数的增加,Core基因数的变化幅度降低,共有基因数趋于稳定(图4)。随着菌株数增多,Pan基因数变多,变化幅度降低,6株菌的 Pan基因数目在24 000以上(图5)。

表5 Core Pan基因统计表

图4 Core 基因盒图Fig.4 Core gene box figurei

图5 Pan 基因盒图Fig.5 Pan gene box figure
基于CorePan分析结果进行系统进化分析发现,木贼镰孢D25-1与假禾谷镰孢遗传距离最近,其次是燕麦镰孢,与麦根腐平脐蠕孢的遗传距离最大(图6)。
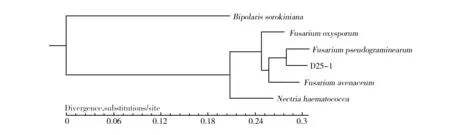
图6 木贼镰孢D25-1的系统进化树Fig.6 The phylogenetic tree of Fusarium equiseti D25-1
3 讨 论
本研究通过14个数据库注释到木贼镰孢D25-1的功能基因13 134个,占全基因组94.97%,注释结果全面良好,有多个基因与木贼镰孢细胞过程、新陈代谢、分子功能、致病性相关,并发现玉米赤霉烯酮合成酶相关的产毒基因2个。玉米赤霉烯酮(Zearalenone,ZEN)是一种重要的真菌毒素,又称 F-2毒素,多由镰孢菌产生,其污染范围广,污染率高[36],在霉变的玉米、小麦、高粱、燕麦、小米等谷类作物、花生等油料作物及其农副产品(如饲料和加工食品等)中检出率高[37-39]。其性质稳定,不易降解,有类雌激素的作用,通过粮食加工产品或畜产品食入动物或人体内,会造成人和家畜生殖激素分泌紊乱,破坏生殖系统,造成畸形、流产、死胎等生殖疾病[40]。此外,玉米赤霉烯酮还具有免疫毒性[41]、细胞毒性[42]、遗传毒性[43]、肝毒性等毒性作用[44],可以降低人畜免疫力,引起肝肾功能衰退紊乱、基因突变等,甚至诱发癌变[45]。由此可见,玉米赤霉烯酮不但污染农作物,造成生产上的损失,而且还会随着食物进入食物链,对人畜健康造成巨大的威胁。因此,应加强对玉米赤霉烯酮相关基因及调控机理的研究,从根本上阻断玉米赤霉烯酮的污染。
本研究通过对木贼镰孢全基因组注释、系统进化分析及产玉米赤霉烯酮规律的测定,确认两个与玉米赤霉烯酮合成酶相关的基因,但其基因功能验证及调控机理还有待进一步研究。在植物病原真菌方面,禾谷镰孢(Fusariumgraminearum)、轮枝镰孢(Fusariumverticillium)、尖镰孢(Fusariumoxysporum)、燕麦镰孢(Fusariumavenaceum)、Eremotheciumgossypii、Magnaporthegrisea、Nectriahaematococca、Ustilagomaydis全基因组均已测序完成,为相应的病害控制策略研究以及抗生素药物开发等提供了大量基础数据。本研究通过比较基因组学分析发现木贼镰孢与假禾谷镰孢遗传距离最近,其次是燕麦镰孢。目前数据库中关于镰孢菌的产毒基因及全基因组数据相对较少,因此,某个基因具体的功能和作用机制还有赖于后基因组学的研究及验证,分类地位也可能会随数据库中全基因组数据信息量的不断扩增而发生变化。
猜你喜欢
中国饲料(2022年5期)2022-04-26
食品安全导刊·中旬刊(2022年3期)2022-04-15
河北果树(2022年1期)2022-02-16
植物保护(2019年2期)2019-07-23
江苏农业科学(2016年1期)2017-05-17
湖北农业科学(2016年24期)2017-03-18
吉林农业(2016年12期)2017-01-06
中国粮油学报(2016年1期)2016-02-06
中国兽药杂志(2013年6期)2013-10-09
