
代谢组学在水产动物毒理学研究中的应用
2021-07-09 06:16陈天赐王小翠
饲料工业 2021年12期
■陈天赐 李 玲 王小翠 郭 慧
(广东海洋大学水产学院,湛江市海洋生态与养殖环境重点实验室,广东湛江 524088)
近年来,代谢组学技术在水产动物病理学、生理学、毒理学及水产品质量检测等方面都有应用[1-3]。在毒理学的研究中,传统的毒性测试实验具有通量低、周期长、敏感度低等不利因素[4],随着研究的深入人们发现毒性胁迫时生物体内的内源性小分子代谢物也会发生显著的变化[1],代谢组学这种快速、高效的检测技术逐渐在毒理学研究中得到广泛应用。代谢组学应用于毒理学技术研究的基本原理是当毒性物质胁迫生物体时,造成生物体内代谢途径中内源性代谢物的变化,代谢组学的检测技术能对生物体受到胁迫之后的细胞、组织或者整个生物体的各种代谢通路中涉及的化合物进行定性定量分析,可以很好地证明生物体在受到毒性胁迫后细胞、组织或生物体受到的损害[4]。与基因组学不同对的是,代谢组学可以证明毒性物质对生物体造成的最终影响,而且在不同的生物体中,基因的结构和功能可能各不相同,但是代谢物在不同物种间结构与功能是高度保守的[5],例如丙酮酸、乳酸这些代谢物,无论是在人类、老鼠、植物中其结构与功能都是一样的,这也意味着代谢组学中相同的分析与计算工具可以适用于任何物种[6]。此外,代谢组学技术得到的信息不一定与我们试验假设的结果有因果关系,可能是其他因素影响的结果,这也有利于我们从中发掘到一些新的信息[5]。
1 代谢组学分析技术
代谢组学常用的分析技术包括核磁共振技术(nuclear magnetic resonance,NMR)、液相色谱与质谱联用技术(liquid chromatography-mass spectrometry,LC-MS)和气相色谱与质谱联用技术(gas chromatog⁃raphy-mass spectrometry,GC-MS)三种[7]。Bloch等[8]和Purcell等[9]发现,当具有奇数核子(包括质子和中子)的原子核置于磁场中,在外加特定频率的射频场时,会产生原子核吸收射频场能量的现象,这就是核磁共振现象[10],核磁共振吸收的大小不仅与原子核本身有关,而且与所受的磁场强度有关[11]。其中氢谱(1H-NMR)是目前广泛应用的核磁共振检测技术[2];1985年北京分析测试学术报告会暨展览会国际会议上首次展出GC-MS联用仪,此后,各种联用技术在对复杂化学物质分析过程中不断发展[12],其中色谱与质谱联用技术(主要是LC-MS与GC-MS技术)结合了色谱高分离和质谱高鉴别性能的优点,在生命科学和医药研究的各领域广泛应用[13]。
2 代谢组学在水产动物毒理学研究中的应用
代谢组学在水产动物研究中的应用主要是通过分析水产动物受毒性物质胁迫后组织、细胞或生物体中的代谢物随时间的变化,确定毒性作用的靶器官和组织、毒性作用的过程和生物标志物,以进行毒性物质作用机制研究或毒性评价[14]。
2.1 代谢组学在水产动物重金属毒性研究中的应用
甲壳类动物是渔业资源重要组成部分,是浅近海捕捞和养殖的重要对象。Ji等[15]对重金属污染海域下的脊腹褐虾(Crangon affifinis)进行研究。1H-NMR分析表明,黄河入海口海域脊腹褐虾丙氨酸、精氨酸、谷氨酰胺含量上升,可能与脊腹褐虾的渗透调节相关,丙二酸、磷酸胆碱、AMP含量的变化可能与脊腹褐虾能量代谢相关。莱州湾海域脊腹褐虾氨基酸和其他渗透压相关有机物的变化则证明重金属污染对脊腹褐虾渗压调节的影响,ATP、AMP含量升高,葡萄糖含量的显著降低,表明金属污染引起能量消耗的增强,而乳酸含量的减少证明无氧呼吸的减弱,此外,也发现丙氨酸和精氨酸可以作为重金属镉(Cd)污染的生物标记物,缬氨酸、亮氨酸、异亮氨酸和酪氨酸可以作为重金属砷(As)污染的生物标记物。Yu等[16]基于1HNMR的代谢组学对渤海金属污染海域的三疣梭子蟹(Portunus trituberculatus)进行研究,其中三个位点三疣梭子蟹Cd含量均超过了海洋生物质量标准一级,S3562位点Cu含量超过了海洋生物质量标准二级。代谢组学分析结果显示三个位点中三疣梭子蟹支链氨基酸含量均处于较低水平,支链氨基酸在免疫细胞蛋白质合成中起重要作用,因此,高浓度的Cd可能诱导了三疣梭子蟹的免疫响应。S6151位点磷酸胆碱、丙二酸含量升高,乙酰乙酸含量下降,证明金属污染影响三疣梭子蟹三羧酸循环和脂质代谢。S6351位点ATP含量显著上升,2-羟基戊二酸含量下降,可能是2-羟基戊二酸脱氢酶活性下降造成对ATP合成酶的抑制减弱,导致ATP含量的升高和糖原减少,此外,肌苷含量减少可能造成肌肉运动的减缓。S3562位点牛磺酸、甘氨酸、龙虾胆碱含量显著升高,但是酪氨酸、组氨酸、苯丙氨酸等含量显著的下降,可能是金属胁迫下三疣梭子蟹通过降低氨基酸含量调节渗透平衡。Ding等[17]用GC-MS代谢组学研究慢性Pb暴露对日本沼虾(Macrobrachium nipponense)幼虾的影响,发现氧代脯氨酸和赖氨酸含量较高,牛磺酸、鸟氨酸、甘氨酸和脯氨酸的含量较低,有相关研究认为牛磺酸与抗氧化和线粒体代谢调控相关,而且Pb的毒性机制之一是氧化胁迫,因此,牛磺酸降低表明Pb胁迫引起了氧化应激反应。甘氨酸是发育中神经系统的主要神经递质,鸟氨酸是大脑中最丰富的氨基酸,因此,Pb胁迫也可能会造成神经系统的损伤。
鱼类是最古老的脊椎动物,存在于地球上大部分的水生环境中,因此水体污染极可能对水体中鱼类动物的健康造成影响。Cappello等[18]用以1H-NMR代谢组学方法研究Hg对金鮻(Liza aurata)鳃的毒性作用,代谢组学分析显示肌酸、乳酸、丙氨酸浓度上升,甘油磷酸胆碱、还原性谷胱甘肽含量下降。鳃中肌酸含量的上升可能是生成更多Na+/K+-ATP以维持渗透压的平衡,乳酸含量的上升证明在Hg胁迫下机体通过无氧呼吸产生更多的能量以维持机体正常代谢,丙氨酸可以将游离氨携带到肝脏中,丙氨酸含量上升可能是为了减少毒性胁迫下蛋白质分解产生的大量游离氨基酸,甘油磷酸胆碱的显著下降可能与细胞膜修复相关,还原性谷胱甘肽减少可能是作为抗氧化剂应对Hg胁迫。Khan等[19]基于1H-NMR技术研究了Pb和Cd毒性作用下银鲫(Carassius auratus gibelio)的血液代谢谱,Pb暴露的银鲫血液丙氨酸、赖氨酸和酪氨酸显著下降,Cd暴露后,赖氨酸、异亮氨酸、亮氨酸、丙氨酸含量显著下降,3-羟基丁酸、丙酮、乳酸、胆碱、肌苷、鸟苷和苏氨酸含量增加,Pb和Cd共暴露时丙酮酸、鸟苷和肌苷含量的增加比单一毒性物质暴露时更加显著,结果证明Pb暴露影响银鲫的神经传导与氨基酸代谢,此外,Pb或Cd暴露均能干扰能量代谢和渗透调节,Pb和Cd共暴露时对能量代谢的影响更加显著。Steven等[20]运用非靶向NMR技术研究了雨季与旱季两个不同区域(正常水体和金属Sb污染水体)食蚊鱼(Gambusia holbrooki)的代谢物质差异,结果发现在不同时间地点采集的样品代谢物种类相似,但是在旱季采集的样品,金属污染组代谢物的数量具有显著的差异,金属污染组大多数代谢物的含量上升,少数几种与金属结合、排泄、解毒等功能相关代谢物含量显著下降,其中天冬氨酸、组氨酸、肌醇、牛磺酸和胆碱含量可以作为金属毒性的生物标记物。
软体动物因分布广泛、易收集和对带毒物质耐受性强等特点常常用做环境检测生物。Wu等[21]研究菲律宾蛤(Venerupis philippinarum)对金属污染的生物监测作用,测试了两个家系的蛤对海洋金属Cd和Zn毒理学效应,结果表明,在Cd或Zn胁迫下两种蛤的代谢物之间具有显著的差异,例如在Cd胁迫的白蛤中发现了支链氨基酸、脯氨酸和糖原增加,谷氨酸减少,而在斑马蛤中却发现了谷氨酰胺增加,精氨酸、乙酰乙酸和天冬氨酸减少。此外,在Zn胁迫下两种蛤渗透调节相关代谢物(例如:牛磺酸、亚牛磺酸、龙虾胆碱)的变化也具有显著的差异。这表明两种蛤对金属胁迫具有不同的响应机制。Zhang等[22]利用1H-NMR代谢组学技术研究Cu对菲律宾帘蛤(Ruditapes philippi⁃narum)的毒性影响,结果显示与渗透压调节、柠檬酸循环相关的代谢物质发生显著的变化,例如龙虾胆碱、支链氨基酸的增加,琥珀酸、丙氨酸和二甲胺含量减少,因此,Cu胁迫可能干扰菲律宾帘蛤渗透调节和能量代谢。Wu等[23]运用蛋白质组学和代谢组学方法对不同盐度下As对菲律宾蛤(Ruditapes philippinarum)鳃的毒性作用进行研究,发现正常盐度(31.1 psu)下As胁迫造成琥珀酸盐和延胡索酸盐水平上升,可能与鳃的能量代谢相关,中低盐度(23.3 psu和15.6 psu)下甜菜碱浓度明显的上升,可能是As胁迫造成对菲律宾蛤鳃的渗透压的影响,低盐度(15.6 psu)下As胁迫造成ATP和甜菜碱含量显著上升,缬氨酸、亮氨酸、异亮氨酸、苏氨酸和酪氨酸含量下降,可能与细胞能量代谢相关。蛋白质组学技术分析显示ATP合成酶上调及琥珀酰-CoA合成酶和核苷二磷酸激酶的下调,表明As影响菲律宾蛤的能量代谢。代谢组学和蛋白质组学联合分析进一步证实As胁迫影响菲律宾蛤鳃的能量代谢。Yu等[24]运用NMR代谢组学与蛋白质组学研究了亚砷酸盐As(Ⅲ)和砷酸盐As(Ⅴ)对贻贝(Mytilus galloprovincialis)的毒理学效应,代谢组学分析表明As(Ⅲ)和As(Ⅴ)胁迫引起多类代谢物发生显著改变,包括氨基酸、有机渗透物质、能量储存物质,结果表明As(Ⅲ)和As(Ⅴ)可干扰贻贝渗透调节以及免疫应激,As(Ⅴ)造成能量代谢的紊乱。蛋白质组学与代谢组学联合分析进一步证明了As(Ⅴ)胁迫下可以造成贻贝能量代谢的紊乱。
2.2 代谢组学在水产动物其他毒性物质研究中的应用
除重金属外,代谢组学技术也应用到水体其他有毒有害物质研究中(如表1)。例如有机农药、氨氮、抗生素等,代谢组学的分析结果显示,这些水体污染物均在不同程度的影响水产动物内源性代谢物质的差异变化,透过差异代谢物的变化揭示了水体中毒性物质的毒性机制,例如引起水产动物体内免疫、抗氧化、渗透调节、能量代谢、细胞损伤等变化,进而影响其健康生长甚至导致水产动物的死亡。

表1 近年关于代谢组学应用于水产动物毒理学研究的报道
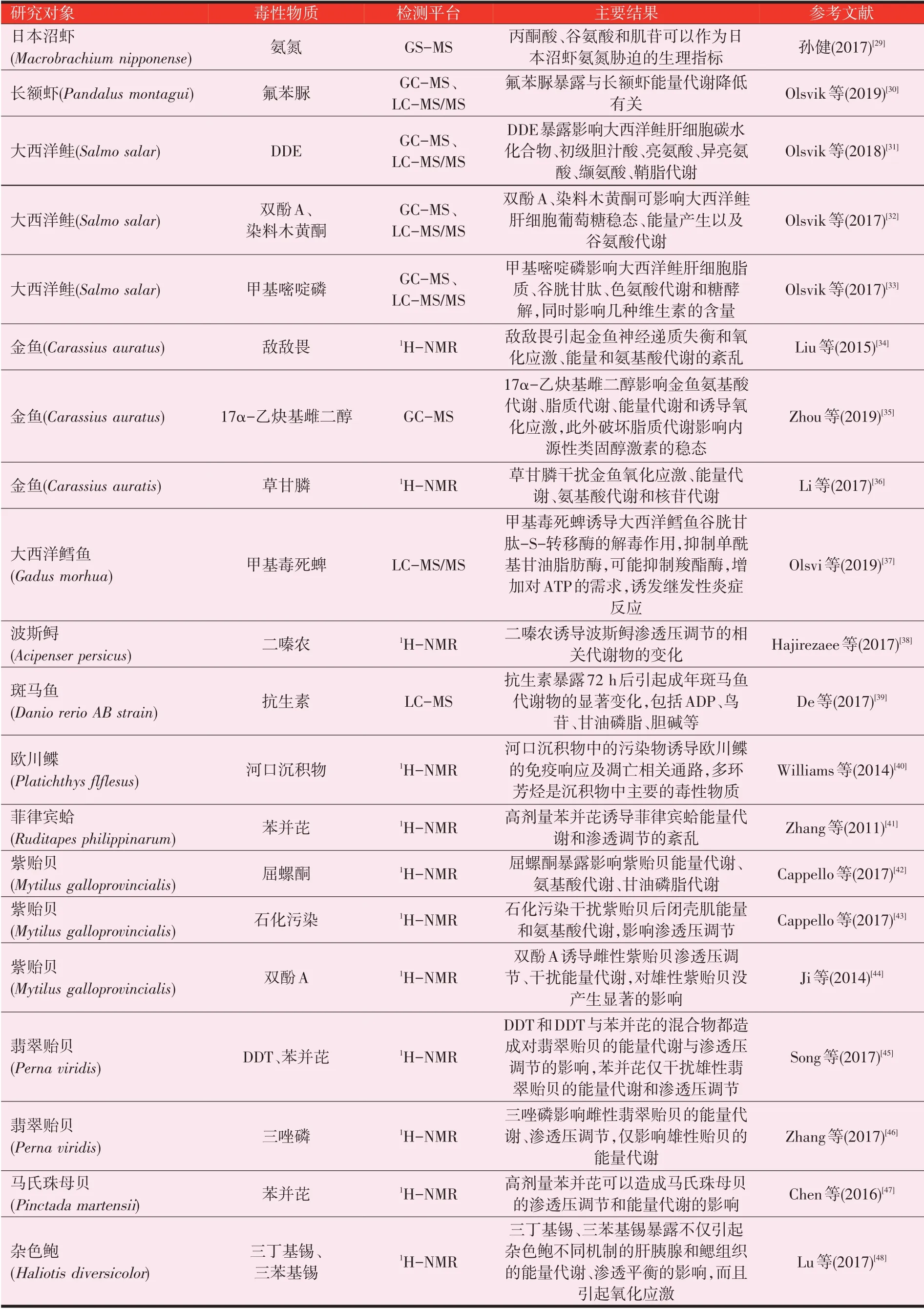
表1(续)近年关于代谢组学应用于水产动物毒理学研究的报道
3 不足与展望
目前代谢组学技术在水产动物毒理学研究中应用广泛,但还存在着一些不足:①代谢组学要求对代谢组中所有代谢物进行分析,但是,目前没有任何一种技术能对生物体内所有的代谢物进行检测[49];②代谢组学可以得到大量的信息,但是,缺乏相应的数据库,信息处理较为繁杂,因此,对这些信息处理的技术还是需要不断地提高才能更好地研究代谢产物的变化是表征的生理或病理的变化[7];③不同年龄阶段、遗传变异、动物品系的代谢产物具有明显的差异,这导致如何判断生物体是发生了病理的变化还是生理的变化成为一个难题[50];④当机体的生理和药理效应超敏时受试物即使没有相关毒性,也可能引起明显的代谢变化,导致假阳性结果[51];⑤灵敏度和动态检测范围方面具有一定的局限性,检测的灵敏度受到检测样品量的影响[7]。但是,代谢组学技术快速、高效的特点是传统毒理学检测手段所不能比拟的,随着代谢组学技术的不断发展,在水产动物毒理学研究中的应用不断成熟,代谢组学技术必定会为促进水产动物毒理学研究做出重大贡献。
猜你喜欢
现代农药(2022年4期)2022-11-19
现代临床医学(2022年4期)2022-09-29
中山大学学报(自然科学版)(中英文)(2022年4期)2022-08-05
昆明医科大学学报(2021年6期)2021-07-31
科学(2020年1期)2020-08-24
当代水产(2019年7期)2019-09-03
农药科学与管理(2019年5期)2019-08-13
大陆桥视野·下(2017年12期)2017-11-29
分忧(2015年3期)2015-06-08
中学生理科应试(2014年12期)2015-01-15
