
海拔和郁闭度对祁连山青海云杉林叶凋落物分解的影响
2021-07-09 09:23赵传燕常亚鹏杨建红
生态学报 2021年11期
李 娜,赵传燕,*,郝 虎,臧 飞,常亚鹏,汪 红,杨建红
1 兰州大学草地农业生态系统国家重点实验室,兰州大学草地农业科技学院, 兰州 730020 2 甘肃祁连山国家级自然保护区管理局,张掖 734000
森林生态系统是最大的营养物质循环库,拥有最大的生物量,枯落物是森林生态系统生物量的重要组成物质[1]。枯落物分解是养分循环和能量流动的重要途径[2]。枯落物分解涉及到复杂的物理化学过程,影响枯落物分解因素众多,包括枯落物本身的性质、气候和土壤(动物和微生物)等,对于环境因素,气候是主导因素[3]。对于生物因素,土壤动物能够物理破碎枯落物,加快分解[4],微生物在枯落物分解后期对难分解物质的降解起到重要作用[5]。近年来,用于枯落物分解的研究植被多集中在杉木、人工林、混交林和马尾松等方面[6],而对青海云杉植被枯落物分解及养分释放方面的研究较少。
祁连山作为我国西北地区高大山系,对河西地区水资源的供给具有重要的贡献。青海云杉 (Piceacrassifolia) 是祁连山山地森林主要乔木优势物种[7],青海云杉叶枯落物分解是祁连山森林生态系统物质循环的重要过程,由于青海云杉林叶分解慢且受多种复杂因素控制,对该重要过程研究较少。地形导致水热组合不同,使得枯落物分解过程随海拔而不同。海拔梯度影响着枯落物分解影响因素,海拔升高,气温降低,湿度降低,辐射增强,这些环境因素使不同海拔梯度下的枯落物分解有差异[8-9]。海拔梯度的不同还影响着枯落物分解的生物因素,如海拔影响土壤pH,pH又是影响微生物群落的关键因子,土壤动物群落及其多样性随着海拔高度的增加而减小[10-11],从而影响枯落物分解。森林郁闭度能改变林下微环境,郁闭度反映森林环境和结构,也反映林冠的郁闭程度[12]。郁闭度影响着林下水热条件,郁闭度的高低影响林下光照、空气温湿度和土壤状况(土壤动物、微生物和水分等),如郁闭度越大,透光率变小,气温降低,湿度增加,土壤含水量增大,也影响着枯落物的分解过程[13- 15]。目前,在祁连山海拔和郁闭度对枯落物分解影响的研究鲜见。
青海云杉林在祁连山分布的海拔跨度较大(2500—3600 m),由于海拔跨度大,青海云杉林在低海拔受水分胁迫,在高海拔受温度胁迫,生境差异导致郁闭度不同[16],本研究采用野外枯落物自然分解试验,模拟不同海拔和郁闭度下青海云杉林叶凋落物的分解过程,探究叶凋落物分解速率以及N、P元素含量的动态变化,青海云杉叶凋落物分解速率随海拔的变化如何,不同的郁闭度下叶凋落物分解如何变化,是理解祁连山区森林生态系统地球化学循环空间异质性要回答的科学问题。揭示气候因子对叶凋落物分解的影响,丰富研究区枯落物分解的理论实践,为物质循环的研究提供数据支撑和理论依据。
1 材料与方法
1.1 研究区概况
研究区位于祁连山黑河上游的天涝池流域(38°20′—38°30′ N, 99°44′—99°59′ E),海拔 2600—4450 m。年均气温-0.6—2.0 ℃,年降水量400—500 mm,属于典型的大陆性高寒半湿润山地气候。青海云杉林主要为藓类青海云杉林,林下发育苔藓层,苔藓层与枯落物组成厚度较大的地被物层,是祁连山青海云杉林生态系统的典型特征[17-18]。
1.2 样地选择和样品制备与采集
在海拔高度为2850 m、3050 m、3250 m和3450 m的青海云杉林中设置固定样地,在海拔2850 m的固定样地中,以样地中心为起点,向东南西北四个方向上分别选择郁闭度为高、中和低(80%、73%和67%)的样点。2015年8月底,在每个固定样地的枯落物收集框中收集青海云杉林凋落物,去除枝条和球果,将剩下的叶子装入自封袋带回实验室处理。将采集的凋落叶用去离子水洗净,65℃烘干至恒重,少许凋落物用于初始化学成分(N、P)的测定,大部装入分解网袋。分解网袋大小为10 cm×8 cm,网袋孔径为1 mm,每个分解网袋中装入5 g凋落叶,将装入凋落叶的分解网袋放在四个海拔梯度的固定样地中及2850 m不同郁闭度样点上。放置前清除地表杂物,凋落物袋与土壤充分接触,放置时间为2015年9月30日,于2015年12月25日、2016年4月30日、6月18日、8月16日和11月18日取样,每次每样点取3袋,每次共取样48袋(即四个海拔处共取样12袋,东南西北四个方向上高中低郁闭度下共取样36袋),共取5次样,取样总计240袋。在每个固定样地布设自动气象站(HOBO U30 Station,USA),用于观测大气温度、降雨等要素,每30 min记录数据一次。
1.3 实验室分析
每次取样后,分解袋中的枯落物用去离子水洗净,65 ℃烘干至恒重,天平(感量为0.001 g)称重,记录。然后将其粉碎过100目筛,用于N、P元素测定分析。首先消煮,称粉碎枯落物样品0.100 g于消化管,加入加速剂(硫酸钾∶硫酸铜=10∶1)1.65 g,再加5 mL浓硫酸,将消化管连同支架放在消化炉(温度为420 ℃)上消化,待溶液为兰绿色,继续煮30 min,消化完成后冷却至室温,然后稀释至50 mL,最后用流动注射分析仪(FlAstar5000 Analyzer, FOSS, Denmark)测定N、P。
1.4 数据处理与分析
根据连续取样的重量,计算枯落物质量残留率(D,%)[19],公式如下:
(1)
式中:Wt为时间t取样的重量(g),W0为初始时间样品的重量(5 g)。根据以下公式是计算分解速率(K,g/d)[20]:
(2)
式中:Wt,W0含义同公式(1),T为取样与放样的时间间隔。凋落物的分解是一个动态过程,利用Olson指数衰减模型[21]拟合凋落物分解的时间动态,公式表示如下:
(3)
式中:Wt,W0含义同公式(1),t是凋落物降解时间,K同公式(2)。凋落物在分解过程中,凋落物的N、P含量将发生变化,或者累积或释放,用累积系数(NAI)[22]表示:
(4)
式中:Wt,W0含义同公式(1),Xt为t时间取样的枯落物N或者P元素的浓度(g/kg),X0为初始放样时间的枯落物N或者P元素的浓度(g/kg)。用来判断养分N或者P在分解过程中的积累与释放。当NAI>100%,养分净累积;当NAI<100%,养分净释放;NAI=100%,既不累积,也不释放。运用Excel对数据进行作图,运用SPSS 23软件对不同海拔和郁闭度下质量损失率、分解速率、N和P含量及养分累积系数进行单因素ANOVA方差分析,并且再利用多重比较(Duncan法)进行显著分析(显著水平α=0.05),大写字母表示相同分解时间不同海拔(郁闭度)间的显著性,小写字母表示同一海拔(郁闭度)不同分解时间之间的显著性。
将30 min的气象要素(温度和降雨)平均或加和获得小时尺度上的数据,同样获得日尺度上的数据,再获得月尺度上数据,利用Excel对数据进行分析并作图。
2 结果与分析
2.1 枯落物分解期间气温和降雨量的变化
气温和降雨的变化如图1所示,从2015年11月到2016年2月,气温均为负值,气温随时间先下降后上升,在1月降到最低,不同海拔下气温由高到低依次是海拔2850 m、3450 m、3050 m和3450 m。2016年4月—10月,气温均为正值,气温随时间先上升后下降,在7月达到最高值,不同海拔下气温由高到低依次是海拔2850 m、3050 m、3450 m和3250 m。
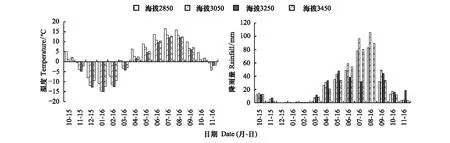
图1 枯落物分解时间内不同海拔下温度和降雨量的月变化图Fig.1 Monthly changes of temperature and rainfall at different altitudes during the decomposition period of litter
冬季(2015年12月—2月),降雨量最小,从2016年3月—11月,降雨量先增大后减小,在7月和8月降雨量最多,春季(3月—5月),海拔3050 m、3250 m下降雨量最大,夏季(6月—8月),不同海拔下降雨量由高到低依次是海拔3050 m、3450 m、2850 m和3250 m,秋季(9月—11月),海拔3050 m、3250 m下降雨量最大。
2.2 青海云杉枯落物质量损失率的变化
2.2.1不同海拔下青海云杉枯落物质量损失率的变化
随着分解时间的延长,不同海拔下枯落物的质量损失率在增大(图2)。在第415天,海拔2850 m、3050 m、3250 m和3450 m下的枯落物质量分别损失了14.89%、16.18%、13.42%和19.67%。分解后期321—415 d的枯落物损失率显著高于分解前期86—262 d(P<0.05)。
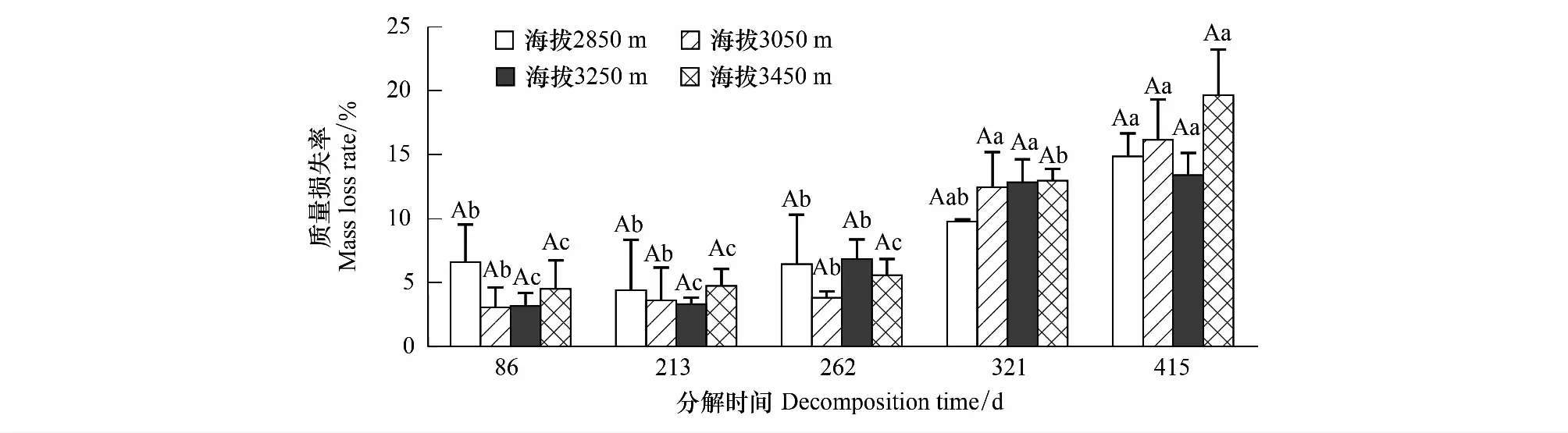
图2 不同海拔下枯落物质量损失率的时间变化Fig.2 Variation of mass loss rate of leaf litter with time at different altitude大写字母表示不同海拔之间的显著性,小写字母表示同一海拔不同分解时间之间的显著性(P<0.05)
2.2.2不同郁闭度下青海云杉枯落物质量损失率的变化
在三种郁闭度下,质量损失率随时间在波动增大(图3)。分解后期321—415 d的枯落物损失率显著高于分解前期86—262 d(P<0.05)。分解415 d时,在高、中和低郁闭度下质量损失率分别是10.73%、13.68%和14.88%。

图3 不同郁闭度下质量损失率的时间变化 Fig.3 Variation of mass loss rate of leaf litter with time under different canopy closures大写字母表示不同郁闭度之间的显著性,小写字母表示同一郁闭度不同分解时间之间的显著性(P<0.05)
2.3 青海云杉枯落物的分解速率
2.3.1不同海拔下青海云杉枯落物的分解速率
在观测期间(放置时间为2015年9月30日,于2015年12月25日、2016年4月30日、6月18日、8月16日和11月18日取样),不同海拔下枯落物分解速率整体上呈先减少后增大的趋势,海拔梯度上差异不显著(图4)。在86—213 d时段分解速率达到最低。
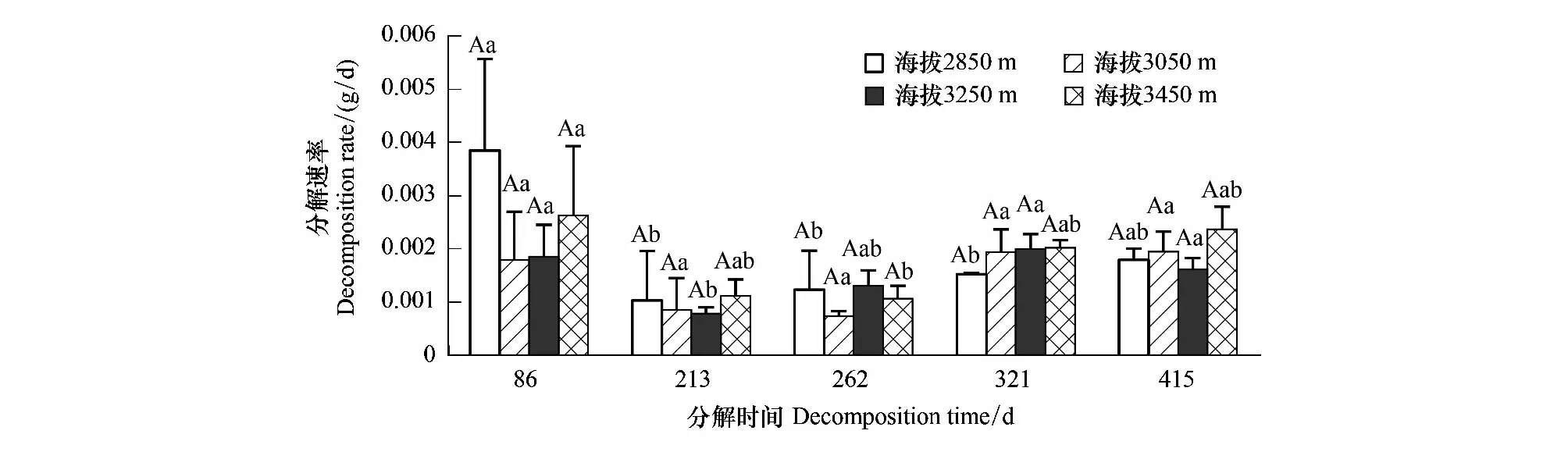
图4 不同海拔下枯落物分解速率的时间变化Fig.4 Variation of litter decomposition rate with time at different altitude大写字母表示不同海拔之间的显著性,小写字母表示同一海拔不同分解时间之间的显著性(P<0.05)
根据叶枯落物残留率y(1-D)随时间t的指数回归方程,估算枯落物分解一半(50%)所需要的时间t0.5和分解95%时所需要的时间t0.95(表1)。从表1可以看出,指数回归方程拟合效果较好,R2介于0.7872—0.8916之间。在海拔2850 m、3050 m、3250 m和3450 m的K值分别是0.127、0.146、0.132和0.171,分解速率从大到小为K3450>K3050>K3250>K2850,分解一半所需时间(t0.5)从海拔由低到高依次为5.5、4.8、5.3、4.1 a,分解95%所需时间(t0.95)从海拔由低到高依次为23.6、20.5、22.7、17.5 a。

表1 不同海拔青海云杉叶凋落物分解模型
2.3.2不同郁闭度下青海云杉枯落物分解速率
在观测期间(放置时间为2015年9月30日,于2015年12月25日、2016年4月30日、6月18日、8月16日和11月18日取样),不同郁闭度下分解速率整体上随时间先减小后增大(图5)。在86—213 d期间,分解速率达到最小,在高郁闭度下差异显著(P<0.05)。同一分解时间下不同郁闭度间的分解速率差异不显著。

图5 不同郁闭度下枯落物分解速率的时间变化 Fig.5 Variation of litter decomposition rate under different canopy closure大写字母表示不同郁闭度之间的显著性,小写字母表示同一郁闭度不同分解时间之间的显著性(P<0.05)
根据叶枯落物残留率y(1-D)随时间的指数回归方程,估算不同郁闭度下枯落物分解一半(50%)所需要的时间t0.5和分解95%时所需要的时间t0.95(表2)。从表2可以看出,指数回归方程拟合效果较好,R2介于0.8358—0.9341之间。高、中和低郁闭度的K值分别是0.101、0.134和0.141,分解速率从大到小为K低>K中>K高,分解一半所需时间(t0.5)由低郁闭度到高郁闭度依次为6.9、5.2和4.8 a,分解95%所需时间(t0.95)低郁闭度到高郁闭度依次为29.7、22.4、和21.2 a。
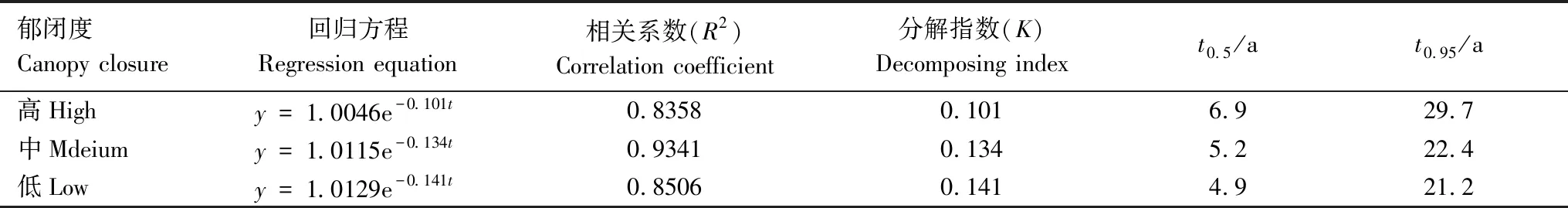
表2 不同郁闭度下青海云杉叶凋落物分解模型
2.4 枯落物分解过程中N和P的含量变化
2.4.1不同海拔下枯落物分解过程中N、P含量变化
不同海拔下枯落物N、P含量变化不同(图6)。分解0—213—262 d,N含量在海拔3050先升高后降低,在213 d,海拔2850 m和3050 m的N含量高于海拔3250 m和3450 m,在海拔3050 m 处显著(P<0.05);P含量在海拔3050 m下先降低后升高,在其余三个海拔下一直在降低,分解213 d,P含量在3050 m含量低于其余三个海拔,在海拔3250 m下显著(P<0.05)。分解262—321—415 d,N含量先降低后升高,且N含量在海拔2850 m处最大,依次是海拔3250 m、3050 m 和3450 m,在321 d最为显著(P<0.05);P含量在海拔3050 m和3450 m下先升高后降低,在海拔2850 m和3250 m下一直在升高,在321 d,海拔2850 m和3250 m的P含量显著低于海拔3450 m和3050 m(P<0.05),在415 d,海拔2850 m和3250 m的P含量显著高于海拔3450 m和3050 m的含量(P<0.05)。
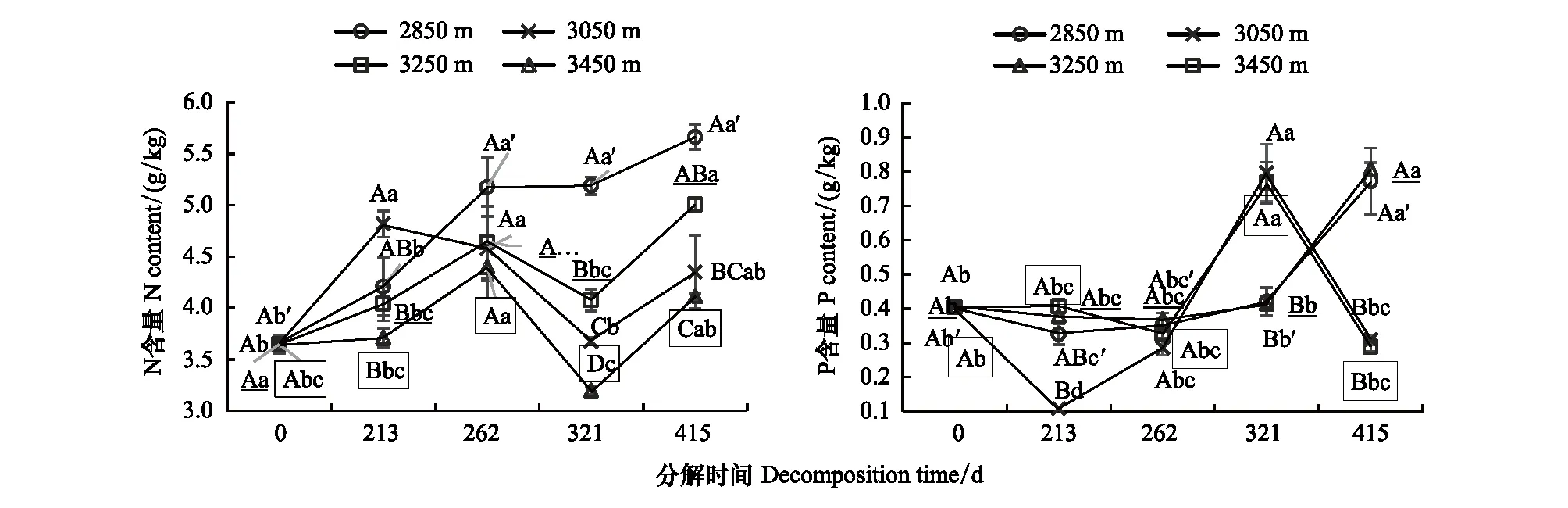
图6 不同海拔下枯落物分解过程中N、P含量的时间变化Fig.6 Changes of N and P contents during litter decomposition at different altitude用x代表图中显著性字母,x′、x、x符号分别表示海拔2850 m、3050 m、3250 m和3450 m的显著性字母标记;大写字母表示不同海拔之间的显著性,小写字母表示同一海拔不同分解时间之间的显著性(P <0.05)
从养分累积系数来看(表3和表4),在海拔2850 m处,N一直处于不同程度的积累状态,在213—321 d,P在释放,在415 d,P在积累。在海拔3050 m,分解321 d,N在释放,P在累积,其余分解时间下N在累积,P在释放。在海拔3250 m,N一直处于积累状态,在213—321 d,P在释放,在415 d,P在积累。在海拔3450 m,在262 d和415 d,N累积,在213 d和321 d,N在释放,在321 d,P在累积,其余分解时间下P在释放。

表3 不同海拔下N的养分累积系数(NAI)/(mean ± SE)%
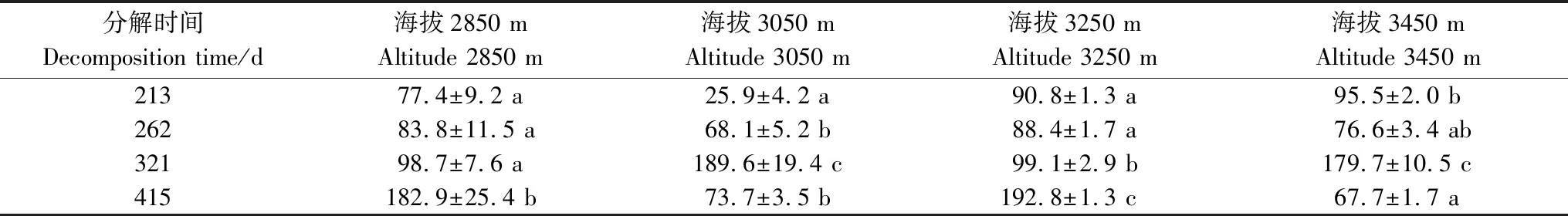
表4 不同海拔下P的养分累积系数(NAI)/(mean ± SE)%
2.4.2不同郁闭度下枯落物分解过程中N、P含量变化
不同郁闭度下N、P含量和养分累积系数变化不同(图7和表5)。在0—213—262 d,N含量在高和中郁闭度下升高,N在累积,在低郁闭度下先降低后升高,N先释放后累积;P含量在中郁闭度下降低,在高和低郁闭度下先降低后升高,不同郁闭度下P都在释放。在262—321—415 d,N含量先降低后升高,除了在321 d,N在中郁闭度下释放,其余情况下N都在累积;P含量在高郁闭度下升高,P在累积,在中郁闭度下先升高后降低,P先累积后释放。在低郁闭度下P含量变化不大,P在释放。相同分解时间不同郁闭度间N含量差异不显著,在分解213 d,中郁闭度下P含量显著高于高郁闭度(P<0.05),其余分解时间下差异不显著。
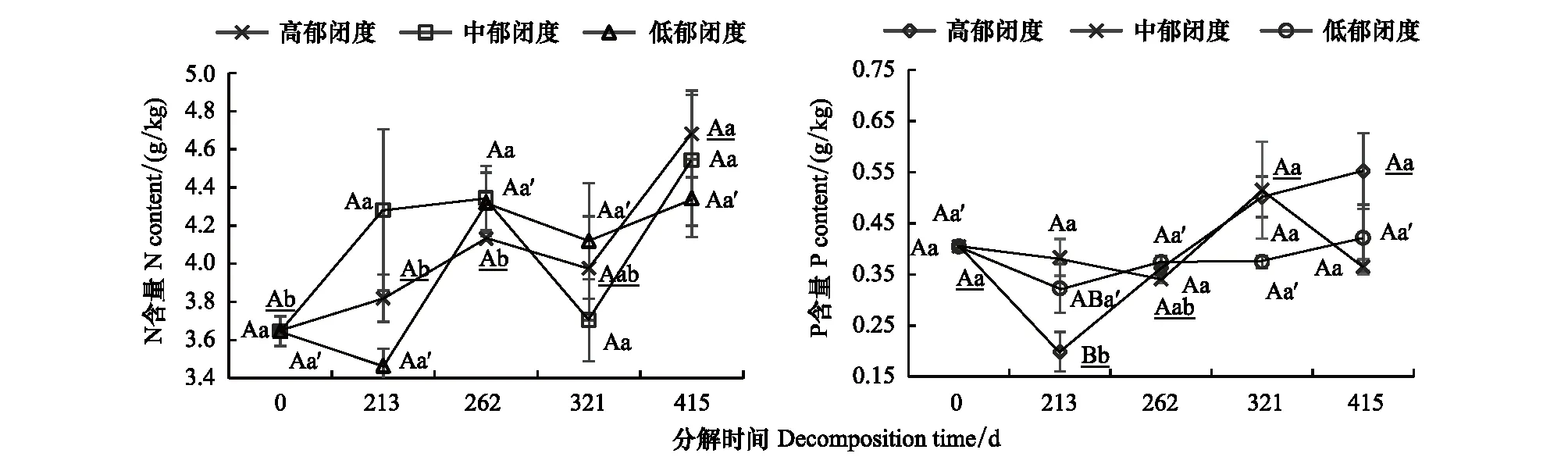
图7 不同郁闭度下N、P含量的时间变化Fig.7 Changes of N and P contents under different canopy closure degrees with time用x代表图中显著性字母, x、x、x′符号分别表示高郁闭度、中郁闭度和低郁闭度的显著性字母标记;大写字母表示不同郁闭度之间的显著性,小写字母表示同一郁闭度不同分解时间之间的显著性(P<0.05)
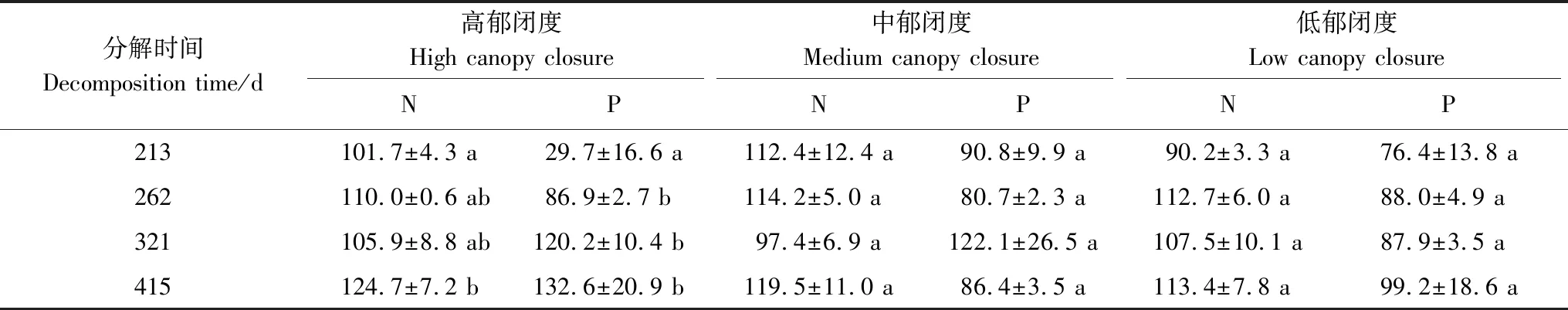
表5 不同郁闭度下N、P养分累积系数(NAI)/(mean ± SE)%
3 讨论
3.1 不同海拔和郁闭度下青海云杉枯落物的分解速率
本研究中,质量损失率是一个累积值,分解时间越长,枯落物损失量越多,损失率自然增大。不同郁闭度和海拔下的分解速率都随时间呈先减小后增大的趋势。与吴艳芹等[23]对一年内草地枯落物的分解速率的研究结果一致。一是由于分解初期,枯落物中大量可溶性成分大量淋失,导致一个较高的分解速率,随后,难分解的物质相对增加,分解速率受微生物控制[24]。二是受季节因素影响,在86—213 d,即取样时间为12月和4月,分解速率减小,是因为经历了冬季,温度较低,微生物活性低。在213—321 d(4月—8月),经历了春节和夏季,温度升高和降雨量的增加,微生物活性增加,增大枯落物的分解速率。在415 d(11月),从夏季过渡到秋季,气温降低,微生物活性受到影响,分解速率增加缓慢。
不同海拔下枯落物分解速率K在海拔3450 m最大,依次是K3050、K3250和K2850。与郭忠玲等[25]对长白山各树种凋落物分解和代力民等[26]对红松针叶凋落物分解的研究结果不尽相同,他们的结果是随着海拔的升高分解速率在减慢。而本研究中没有出现这样的变化规律,主要由于分解网袋所放置的环境(如温湿度、郁闭度、光辐射、坡度、动物和微生物等综合因素)对分解都有一定的影响,研究发现,升温显著增加了凋落物分解[27],湿度适中会加速分解[28],光降解作用能改变枯落物性质,加速分解[29],坡度越大,会延缓分解速率[30],随海拔升高,林分冠幅直径增加,林分密度呈"单峰"变化,对分解有影响[31]。海拔不同,环境因素也不同,环境会影响动物和微生物的分布[32],不同海拔下枯落物分解同时受到非生物因素和生物因素等多种综合因素的影响,同时分解网袋法有局限性,只能反映局部范围内枯落物的分解速率[33]。
不同郁闭度下枯落物分解速率在低郁闭度下最大,依次为中郁闭度和高郁闭度。郁闭度高低由冠层决定,冠层会改变林内微环境,进而影响枯落物的分解[34]。在冠层覆盖对半干旱非洲大草原凋落物分解的影响研究中发现冠层覆盖率越大,枯落物损失率越低,冠层覆盖率小的区域,枯落物暴露于太阳辐射的机率越多,导致地表温度高于高冠层覆盖区,低覆盖形成的相对高辐射和高温将加速枯落物的分解[35]。
3.2 不同海拔和郁闭度间N、P含量变化
淋溶和微生物降解是影响枯落物分解过程中养分变化的主要过程,淋溶是造成养分流失的关键过程[36],微生物降解过程中,会固定氮,氮大多以蛋白质形式存在,从而引起氮累积[37]。温度和水分影响微生物群落,高温促进微生物生物量增加,水分降低,微生物活性也会降低[38-39]。微生物活性的增加和降低影响养分浓度变化[40]。本研究中,在0—213 d(4月),N含量增大,海拔2850 m、3050 m处的N含量一直高于海拔3250 m、3450 m,经历了冬季,温度是主要限制因子,低海拔相对高海拔温度高,微生物量和活性增多,氮绝对量增加[37]。在262—321—415 d(6月—8月—11月),N含量先减小后增大,是因为经历了多雨的夏季,温度和降雨的双重作用使N大量释放,且在海拔2850 m处最大,在3450 m最小,海拔3250 m、3050 m 居中,与温度密切相关。从N的累积系数来看,N大多数情况下处于净累积状态,个别情况,如分解213 d,海拔3450 m处N处于净释放,受低温影响,微生物量少,固氮量少。分解321 d,海拔3050 m、3450 m处N在净释放,正值夏季,降雨的影响较大,分解0—321d,海拔3050 m、3450 m处的累积降雨量最大,分别为295.8 mm和231.6 mm (表6)。
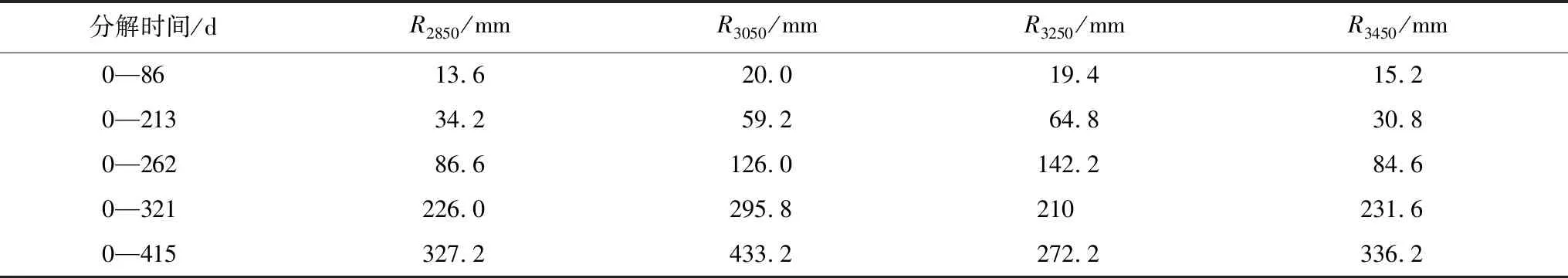
表6 不同海拔下枯落物不同分解时间的降雨量
不同郁闭度下N含量变化主要受土壤温度、湿度和光照的影响。郁闭度低,光照充足,土温高,微生物活性强[41]。在213 d和415 d,即一年中的4月和11月,中高郁闭度下N含量高于低郁闭度,由于祁连山春季和秋季温度较低,低郁闭度下土温降低更快,微生物数量减少,N的绝对含量减少。在262 d和321 d,正是一年中的夏季,即6月和8月,降水较多,低郁闭度下温度较高,水分多,微生物量增多,所以低郁闭度下N含量高于中高郁闭度。不同郁闭度下N大多都在净积累,但在213 d,N在低郁闭度下净释放,在321 d,N在中郁闭度下净释放,这与温度和降水有关。
枯落物分解初期,P以可溶性形态存在,最易大量淋失[42]。所以在分解初期213—262 d,不同海拔和郁闭度下P都在净释放。在213 d,海拔2850 m、3050 m处的P含量高于海拔3250 m、3450 m,中低郁闭度下P含量高于高郁闭度,与微生物活性有关,微生物在降解时,磷的供给限制微生物营养需要时,磷就会释放[22],高海拔和高郁闭下温度相对较低,微生物量少,磷可供微生物营养需要,多余的磷就会释放。在321 d,P含量在海拔3250 m、2850 m低于在海拔3050 m、3450 m,且前者在净释放,后者在净累积,P含量在中高郁闭度下高于低郁闭度,且在中、高郁闭度下净累积,在低郁闭度下净释放。与降水有关,分解321 d(8月),多雨季节,海拔3050 m、3450 m的累积降水量较多,中高郁闭度下的湿度相对较大,微生物活性大,当微生物对P的营养需要不足时,会出现累积。在415 d,即一年中的11月,P在不同海拔和郁闭度下的规律正好与分解321 d的相反,经历了秋季,温度是主要的限制因子。
4 结论
通过研究,得出以下结论:
(1)质量损失率随分解时间的延长呈增大趋势,分解时间越长,质量分解率随时间的差异就越显著。
(2)不同海拔和郁闭度下枯落物分解速率整体上呈先减小后增大的趋势。青海云杉林叶凋落物的半衰期为5 a,分解95%所需时间为22.5 a。
(3)不同海拔和郁闭度下枯落物N、P含量在分解过程中的变化不同,或积累或释放,与季节(环境条件和微生物活性)的影响密切相关。
猜你喜欢
有色冶金节能(2022年1期)2022-03-11
散文诗(2021年22期)2022-01-12
今日农业(2021年1期)2021-11-26
中国种业(2021年7期)2021-08-02
福建林业(2020年5期)2020-03-18
特种经济动植物(2019年9期)2019-10-29
环球时报(2019-05-23)2019-05-23
热带林业(2019年4期)2019-03-05
海外星云(2016年19期)2016-10-24
农民致富之友(2014年7期)2014-10-21
