
不同米曲霉对酱油低盐固态发酵的影响
2021-08-09 03:49石磊刘蕊藤薇侯丽华
中国调味品 2021年8期
石磊,刘蕊,藤薇,侯丽华*
(1.天津利民调料有限公司,天津 300308;2.天津科技大学食品营养与安全国家重点实验室,天津 300457)
酱油在我国传承发展千年,由豆酱逐渐演变成人们日常饮食中重要的液态调味品。通过微生物的作用对蛋白质原料与淀粉类原料进行分解,再经复杂的生化反应从而形成营养丰富、鲜香味美的酱油。现今酱油的种类众多,发酵工艺也不断改良创新。但低盐固态发酵法作为我国的传统发酵工艺,具有生产周期短、简单高效、成本低的特点[1],因此低盐固态发酵酱油仍占据不可取代的市场地位,并且这种价格低廉的中低档酱油也符合低盐健康饮食的理念。
制曲是酱油酿造的首要环节。生产优质成曲一方面可以从优化工艺参数方面入手[2-3],另一方面则涉及到发酵菌种的选择运用。米曲霉在制曲过程中发挥着重要的作用,它分泌产生的各种酶,为酱油发酵的原料分解、物质的转化合成奠定了基础[4]。因此,选择适宜于发酵工艺的优良米曲霉制曲是保证酱油质量的关键。米曲霉沪酿3.042具有生长快、分泌的酶系足够丰富、不产毒素等优点,在我国酱油行业中一直被普遍使用[5]。近年来,研究者们以米曲霉沪酿3.042作为出发菌株进行了菌种改造,得到米曲霉A100-8、米曲霉3.042-3、米曲霉3.042-3-c和米曲霉3.042-3-cd等。本文首次将以上米曲霉分别制曲,通过测定种曲孢子数和大曲酶活力,从中选取优势米曲霉进行低盐固态发酵的对比研究,探究其对酱油品质的影响,为改造构建的优良米曲霉在酱油低盐固态发酵中的实际应用提供了一定的理论参考。
1 材料与方法
1.1 原料与菌种
原料:麸皮、豆粕;菌种:米曲霉为本实验室保藏,见表1。

表1 本研究使用的5种米曲霉Table 1 The five kinds of A. oryzae used in this study
1.2 主要仪器设备
立式压力蒸汽灭菌器 日本雅马拓科技有限公司;Multiskan GO酶标仪 美国Thermo公司;数显电热恒温水浴锅 上海百典仪器设备有限公司;QP-2010气相色谱-质谱联用仪 日本岛津公司
1.3 试验方法
1.3.1 制曲与发酵工艺
制曲与发酵步骤见图1。

图1 制曲与发酵的具体步骤Fig.1 The specific steps of koji making and fermentation
1.3.2 种曲孢子数的测定
按照SB/T 10315-1999规定的方法进行测定。
1.3.3 大曲酶活力的测定
酸性、中性、碱性3种蛋白酶活力按照SB/T 10317-1999的方法测定。1个蛋白酶活力单位定义为在40 ℃条件下每分钟水解酪蛋白产生1 μg酪氨酸所需的酶量。糖化酶活力[8]、纤维素酶活力[9]、果胶酶活力[10]的测定均采用DNS显色法。1个糖化酶活力单位定义为在40 ℃、pH 4.6下每小时水解可溶性淀粉产生1 mg葡萄糖所需的酶量。1个纤维素酶活力单位定义为在40 ℃、pH 4.8下每小时水解CMC产生1 μmol葡萄糖所需的酶量。1个果胶酶活力单位定义为在45 ℃、pH 4.4下每分钟水解果胶质产生1 μg D-半乳糖醛酸所需的酶量。
1.3.4 发酵过程中霉菌数的测定
将酱醪取出,用无菌水进行10倍梯度稀释,选2~3个适宜稀释度的样品稀释液涂布于孟加拉红培养基平板进行计数。
1.3.5 酱油各项理化指标的测定
发酵结束后,将酱醪称重,加入1倍80 ℃左右的蒸馏水充分混合浸泡后,使其自然沉降,再用离心机在8000 r/min离心10 min,过滤去除残渣后得到的酱油用于测定各项理化指标。还原糖含量采取DNS显色法测定[11]。全氮和可溶性无盐固形物的含量按照国标GB 18186-2000规定的方法测定。氨基酸态氮与总酸含量按照国标GB 5009.235-2016规定的方法测定。
1.3.6 酱油颜色指数的测定
将10倍稀释的酱油液在不同波长下测定OD值,以蒸馏水作空白对照,计算红色指数、黄色指数和色深物质[12]。
1.3.7 酱油风味物质的检测
采用固相微萃取-气质联用(SPME-GC-MS)的方法检测[13]。
风味物质的鉴定:手动积峰,对照质谱仪记录的质谱图与检索数据库提供的标准质谱图,筛选相似度在80%以上的物质,定性分析,并以面积归一化法计算相对含量。
2 结果与分析
2.1 种曲孢子数的跟踪测定
由图2可知,5种米曲霉的种曲孢子数在第24~48 h期间增长缓慢,第48 h后,出现快速增长的趋势。在第72 h种曲孢子数均达到最大值,并且满足工业生产对种曲孢子数的要求(5×109个/g)。整体而言,多核米曲霉的种曲孢子数>单核米曲霉,即米曲霉沪酿3.042>米曲霉3.042-3;米曲霉3.042-3-cd>米曲霉3.042-3-c。在第72 h种曲收曲时,5种米曲霉的种曲孢子数从大到小依次为:米曲霉3.042-3-cd(7.50×109个/g)>米曲霉沪酿3.042(7.40×109个/g)>米曲霉3.042-3-c(7.29×109个/g)>米曲霉A100-8(7.15×109个/g)>米曲霉3.042-3(6.88×109个/g)。

图2 米曲霉种曲孢子数增长情况的比较Fig.2 The comparison of the number of spores of Aspergillus oryzae
2.2 大曲酶活力的测定
在大曲阶段的第36 h、第40 h和第44 h测定5种米曲霉的6种酶活力,综合比较5种米曲霉的产酶情况,为酱油发酵奠定了良好的基础。
2.2.1 蛋白酶活力
米曲霉具有分泌蛋白酶的强大能力。蛋白酶分解高温蒸煮变性的蛋白质原料,对酱油氨基酸、多肽、风味前体物质的产生具有重要作用。本试验对酸性、中性、碱性3种蛋白酶活力的测定结果见图3~图5。

图3 大曲阶段酸性蛋白酶活力的变化Fig.3 The changes of acid protease activity during Daqu stage

图4 大曲阶段中性蛋白酶活力的变化Fig.4 The changes of neutral protease activity during Daqu stage

图5 大曲阶段碱性蛋白酶活力的变化Fig.5 The changes of alkaline protease activity during Daqu stage
由图3可知,除米曲霉3.042-3-cd的酸性蛋白酶活力在第44 h出现小幅下降以外,其他4株米曲霉的酸性蛋白酶活力在第36~44 h均逐渐缓慢升高,第44 h酸性蛋白酶活力达到最高。其中米曲霉A100-8的酸性蛋白酶活力在测定的3个时间点均高于其他菌株。米曲霉A100-8是经诱变选育出的一株高产酸性蛋白酶菌株,说明其高产酸性蛋白酶的特性可以稳定遗传。但总体来看,大曲阶段各时间点的酸性蛋白酶活力均较低,米曲霉A100-8酸性蛋白酶活力在第44 h时最大值仅达到496.96 U/g,一方面可能与大曲中的pH环境有关,另一方面可能与所用原料有关,豆粕和麸皮与大豆和小麦混合原料制曲相比空隙小,空气穿透性较差,不利于曲霉生长产酶分解原料[14]。
由图4可知,在大曲阶段,中性蛋白酶活力远高于酸性蛋白酶,这也与研究乳酸菌与米曲霉共制曲时测定的酶活结果一致[15]。5株米曲霉的中性蛋白酶活力在制曲第36~44 h期间呈缓慢升高的趋势,在第44 h时均达到酶活力的最大值。米曲霉3.042-3-cd的中性蛋白酶活力在第44 h增加最多,并超越了其他米曲霉,达到1676.87 U/g。其他米曲霉的中性蛋白酶活力在第44 h差异不大,均在1450 U/g左右。
由图5可知,5株米曲霉的碱性蛋白酶活力变化不同。米曲霉沪酿3.042、米曲霉3.042-3、米曲霉3.042-3-c和米曲霉3.042-3-cd的碱性蛋白酶活力在制曲第36~44 h呈现先下降后上升的趋势。而米曲霉A100-8的碱性蛋白酶活力几乎维持在稳定的水平。但总体来看,每种米曲霉的碱性蛋白酶活力自身变化幅度不大,并且在第36 h、第40 h与第44 h均达到1000 U/g以上,其中米曲霉3.042-3碱性蛋白酶的活力最高,米曲霉A100-8酶活力次之。
2.2.2 糖化酶活力
糖化酶同样是酱油发酵中起到关键作用的酶系之一。淀粉质原料通过淀粉酶的作用,形成小分子糖类物质等成分,有利于提高酱油的口感,也为糖酵解途径提供了基础,从而增加了成品酱油中芳香物质的含量。
由图6可知,5株米曲霉的糖化酶活力在第36~40 h呈逐渐上升的趋势,第44 h时米曲霉3.042-3-cd与米曲霉A100-8的糖化酶活力出现小幅度下降。但米曲霉A100-8始终具有最高的糖化酶活力,第40 h达到最大值3324.10 U/g,第44 h虽然酶活力下降,但仍达到3056.85 U/g,因此米曲霉A100-8在分解淀粉质原料上具有优势。与其余4株米曲霉相比,米曲霉3.042-3-cd具有较高的糖化酶活力。

图6 大曲阶段糖化酶活力的变化Fig.6 The changes of glucoamylase activity during Daqu stage
2.2.3 纤维素酶活力与果胶酶活力
纤维素酶和果胶酶的存在使原料充分分解,提高了原料的利用率。但是米曲霉分泌过多的纤维素酶和果胶酶会影响淋油工艺,造成酱油无盐固形物含量降低等不利影响。
由图7可知,在制曲第36~40 h,5株米曲霉的纤维素酶活力呈下降趋势。但在第44 h,纤维素酶活力出现回升。整个制曲期间,米曲霉沪酿3.042保持最高的酶活力,而米曲霉3.042-3-c的酶活力远低于米曲霉沪酿3.042,第40 h为106.10 U/g,第44 h为205.74 U/g。其次酶活较低的为米曲霉3.042-3-cd,这说明整合到米曲霉的CreA基因及cis-acting顺式作用元件仍稳定地发挥作用。
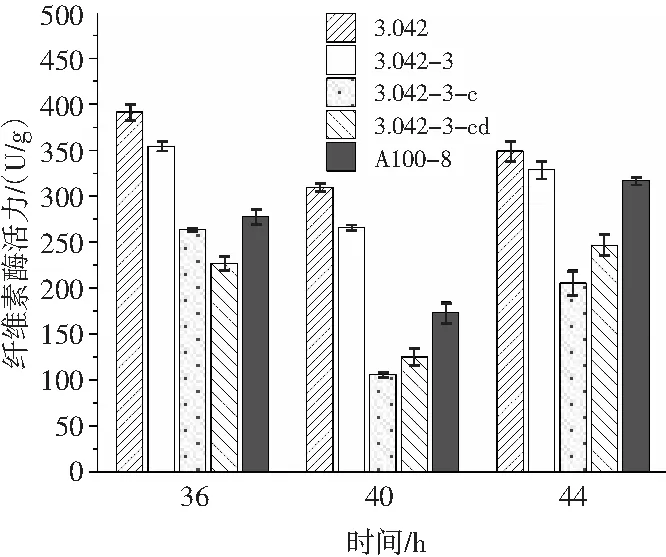
图7 大曲阶段纤维素酶活力的变化Fig.7 The changes of cellulase activity during Daqu stage
由图8可知,5株米曲霉的果胶酶活力均呈先下降后上升的趋势。米曲霉沪酿3.042的果胶酶活力最高。米曲霉3.042-3-cd的果胶酶活力虽在第36 h出现最大值372.44 U/g,但后期出现大幅度降低。第44 h时,米曲霉3.042-3-c的果胶酶活力最低,为220.91 U/g,其次是米曲霉3.042-3-cd,为274.77 U/g。

图8 大曲阶段果胶酶活力的变化Fig.8 The changes of pectinase activity during Daqu stage
综上所述,通过酶活力的测定,确定第44 h收曲,此时各种酶活力旺盛。在5种米曲霉中,米曲霉A100-8在分泌酸性蛋白酶活力和糖化酶活力上具有明显优势,米曲霉3.042-3-cd种曲孢子数多,其表达出的纤维素酶活力与果胶酶活力降低的性状能够稳定遗传并且在第44 h收曲时中性蛋白酶最高。所以,选择米曲霉A100-8与米曲霉3.042-3-cd分别制曲,进行低盐固态发酵。
2.3 发酵过程中霉菌数的变化
发酵过程中,每隔8 d取样,记录酱醪中霉菌数的变化情况。
由图9可知,霉菌数随着发酵的进行呈现降低的趋势。第8 天时,霉菌数已降到约为4.5 log CFU/g。第32 天发酵结束,霉菌数约为1.5 log CFU/g。这说明在酱油盐水发酵阶段,高温环境严重限制了米曲霉的生长。对比两种米曲霉制曲发酵,米曲霉3.042-3-cd比米曲霉A100-8霉菌数略高。

图9 发酵期间霉菌数的变化Fig.9 The changes of mold count during fermentation
2.4 酱油理化指标的测定
发酵结束后,对最终酱油进行了理化指标的对比分析,结果见表2。

表2 酱油各项理化指标的比较Table 2 Comparison of physicochemical indexes of soy sauce
氨基酸态氮、全氮与可溶性无盐固形物是评价酱油等级的重要指标。还原糖促进了酱油色泽与风味的形成[16]。发酵过程中pH降低,微生物发酵产酸等导致总酸的积累。由于米曲霉A100-8具有较高的酸性蛋白酶活力与糖化酶活力,使得米曲霉A100-8制曲的发酵酱油在氨基酸态氮、全氮、还原糖及无盐固形物含量上高于米曲霉3.042-3-cd,尤其是还原糖明显增加,而米曲霉3.042-3-cd制曲的发酵酱油总酸含量高。
2.5 发酵结束后颜色指数的测定
红色指数、黄色指数及色深物质可以一定程度反映酱油的呈色情况,测定结果见表3。
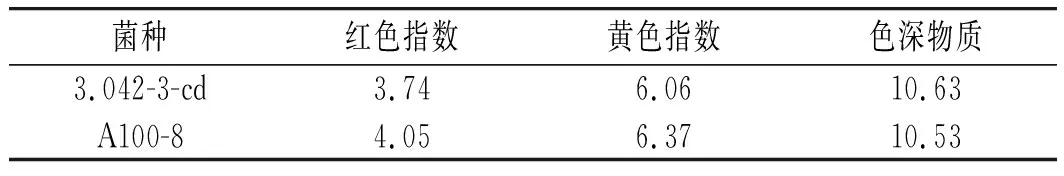
表3 酱油颜色指数的对比Table 3 Comparison of color indexes of soy sauce
一般红色指数超过3.0表明酱油红色比较显著。黄色指数与红色指数成正相关,黄色指数较高,炒菜上色不发乌[17]。由表3可知,红色指数和黄色指数较高的为米曲霉A100-8发酵酱油。色深物质则相反,指数较高的为米曲霉3.042-3-cd发酵酱油。
2.6 酱油风味物质的检测
通过GC-MS对两组酱油进行风味物质的检测,各类风味物质的相对含量见图10。

图10 酱油各类风味物质的相对含量Fig.10 The relative content of flavor compounds in soy sauce
由图10可知,本试验低盐固态发酵中醛类、醇类、酸类、酚类占比大,总相对含量达到90%左右,剩余10%的风味物质为酮类、酯类与其他类。米曲霉3.042-3-cd发酵酱油在酸类、醇类、酚类和其他类物质的相对含量高于米曲霉A100-8,而米曲霉A100-8发酵酱油在醛类与酯类风味物质的相对含量上相比于米曲霉3.042-3-cd都有微弱的提升,具体风味物质的相对含量及检测的种类个数见表4。

表4 酱油风味物质的相对含量及个数Table 4 The number and relative content of flavor compounds in soy sauce

续 表
由表4可知,米曲霉A100-8发酵酱油中检测到风味物质的种类较多,总计达到52种。米曲霉3.042-3-cd为46种。两组酱油均是醛类检测到的种类最多,主要以苯甲醛、苯乙醛、异戊醛为主,具有杏仁、焦糖味的苯甲醛和苯乙醛是许多食品中的芳香活性化合物,而异戊醛具有果味和麦芽味。酸类中主要以乙酸为主,醇类中占比较大的依次是异戊醇、苯乙醇、1-辛烯-3-醇。在酚类物质的检测中,愈创木酚的相对含量最高。另外,3.042-3-cd制曲发酵检测到微量的4-乙基-2-甲氧基苯酚(4-EG)成分。4-EG这种呈香物质的产生被认为与木质素的代谢有关,可以增加酱油的烟熏味。酯类、酮类和其他类(呋喃吡嗪类)也是酱油风味的重要组成部分,尤其是酯类具有强挥发性,使酱油的香气更协调。但总体而言,两组酱油酯类的相对含量较少,另外,与市售的低盐固态酱油相比,醇类的相对含量较少[18],这可能与本试验的低盐固态工艺特点有很大关系,需要进一步探究。
3 结论
本文通过对沪酿3.042、3.042-3、3.042-3-c、3.042-3-cd和A100-8 5株米曲霉种曲孢子数与大曲酶活力的测定发现米曲霉A100-8在第44 h收曲时具有最高的酸性蛋白酶活力(496.96 U/g)与糖化酶活力(3056.85 U/g)。米曲霉3.042-3-cd种曲孢子数最大(7.50×109个/g),生长快并且纤维素酶与果胶酶活力降低的性状能得到稳定遗传。在低盐固态发酵工艺中,运用米曲霉A100-8与米曲霉3.042-3-cd制曲发酵的酱油在理化指标上均为同一等级水平。米曲霉A100-8发酵酱油的颜色指数较高,风味物质种类更丰富。而米曲霉3.042-3-cd在酸类和醇类物质的相对含量上高于米曲霉A100-8。在实际生产中,可根据需要选择适宜的米曲霉来提升低盐固态酱油的品质。
猜你喜欢
中国调味品(2021年3期)2021-03-23
酿酒科技(2020年7期)2020-12-19
中国调味品(2019年11期)2019-11-15
食品与生物技术学报(2019年12期)2019-02-15
食品工业科技(2018年21期)2018-11-27
中成药(2018年7期)2018-08-04
食品与生物技术学报(2018年3期)2018-03-25
湖南农业大学学报(自然科学版)(2015年3期)2015-07-13
中国果菜(2015年2期)2015-03-11
中国酿造(2014年9期)2014-03-11
