
五味子酯甲通过抑制CCAT1和PI3K-AKT信号通路抑制肺癌细胞的迁移和侵袭
2021-08-26 21:55谭晖王吉昌董丹凤李恩孝李毅
世界中医药 2021年13期
谭晖 王吉昌 董丹凤 李恩孝 李毅
摘要 目的:探究五味子酯甲(SA)对肺癌细胞生长和转移的潜在作用机制。方法:本研究以非小细胞肺癌细胞系A549和H1299作为实验对象,将SA溶解在二甲基亚砜中,用不同剂量的SA(0 μmol/L、5 μmol/L、10 μmol/L、20 μmol/L、50 μmol/L)孵育A549和H1299细胞24 h、48 h和72 h。采用细胞计数盒-8检测细胞增殖,采用FC500流式細胞仪分析细胞凋亡,采用伤口愈合实验评价细胞迁移,采用Transwell检测细胞侵袭。通过实时定量逆转录聚合酶链反应(qRT-PCR)检测细胞中CCAT1 mRNA水平;通过Western Blotting检测细胞中p-PI3K、PI3K、p-AKT、AKT、PTEN、E-cadherin、MMP2和MMP9的蛋白表达水平。结果:与未处理的细胞比较,SA以剂量和时间依赖性方式降低了A549和H1299的细胞活力(P<0.05)。与DMSO组比较,SA 10 μmol/L组的A549和H1299细胞凋亡率显著升高了5.71倍和3.48倍(P<0.001),伤口愈合面积显著降低了63.42%和59.15%(P<0.001),侵袭细胞数显著降低了53.98%和53.36%(P<0.001)。与DMSO组比较,SA 10 μmol/L组A549和H1299细胞中E-cadherin的蛋白表达水平升高,而MMP-2和MMP-9的蛋白表达水平降低(P<0.01),CCAT1的mRNA表达水平均降低(P<0.01),p-PI3K和p-AKT的蛋白表达水平显著降低(P<0.01),PTEN的蛋白表达水平显著升高(P<0.01),而PI3K和AKT蛋白表达水平无明显变化(P>0.05)。结论:SA通过抑制CCAT1和PI3K/AKT信号通路抑制肺癌细胞的生长和转移。
关键词 五味子酯甲;肺癌;迁移;侵袭;增殖;凋亡;结肠癌相关转录本-1;PI3K/AKT信号通路
Abstract Objective:To explore the potential effect and mechanism of Schisantherin A(SA) on the growth and metastasis of lung cancer cells.Methods:In this study,the non-small cell lung cancer cell lines A549 and H1299 were used as experimental objects.SA was dissolved in dimethyl sulfoxide(DMSO),and A549 and H1299 was incubated with different concentrations of SA(0 μmol,5 μmol,10 μmol,20 μmol,50 μmol) for 24 h,48 h and 72 h.Then,Cell Counting Kit-8(CCK-8) was used to detect cell proliferation,FC500 flow cytometer was used to analyze cell apoptosis,wound healing experiment was used to evaluate cell migration,and Transwell was used to detect cell invasion.Real-time quantitative reverse transcription polymerase chain reaction(qRT-PCR) was used to detect CCAT1mRNA levels in cells.Western blot was used to detect the protein expression levels of p-PI3K,PI3K,p-AKT,AKT,PTEN,E-cadherin,MMP2 and MMP9 in cells.Results:Compared with untreated cells,SA reduced the cell viability of A549 and H1299 in a concentration and time-dependent manner(P<0.05).Compared with the DMSO group,the apoptosis rate of A549 and H1299 cells in the SA 10 μmol group was significantly increased by 5.71 times and 3.48 times(P<0.001),the wound healing area of A549 and H1299 cells in the SA 10 μmol group was significantly reduced by 63.42% and 59.15%(P<0.001),and the number of invasion cells of A549 and H1299 cells in the SA 10 μmol group was significantly reduced by 53.98% and 53.36%(P<0.001).Compared with the DMSO group,the protein expression levels of E-cadherin in A549 and H1299 cells in the SA 10 μmol group increased,while the protein expression levels of MMP-2 and MMP-9 decreased(P<0.01); the mRNA and protein expression levels of CCAT1 in A549 and H1299 cells in the SA 10 μmol group were lower(P<0.01); the protein expression of p-PI3K and p-AKT in A549 and H1299 cells in SA 10 μmol group decreased significantly(P<0.01),PTEN protein expression level increased significantly(P<0.01),while PI3K and AKT protein expression levels did not change significantly(P>0.05).Conclusion:SA inhibits the growth and metastasis of lung cancer cells by inhibiting the CCAT1 and PI3K/AKT signaling pathways.
Keywords Schisantherin A; Lung cancer; Migration; Invasion; Proliferation; Apoptosis; CCAT1; PI3K/AKT signaling pathway
中图分类号:R734.2,R285.5文献标识码:Adoi:10.3969/j.issn.1673-7202.2021.13.009
肺癌是公认的发病率和死亡率都很高的常见恶性肿瘤,非小细胞肺癌(Non-small Cell Lung Carcinoma,NSCLC)是最常见的肺癌亚型,占所有病例的近85%[1-3]。传统的化疗药物在NSCLC患者中引起的不良反应较大,这些不良反应往往直接导致患者死亡。此外,分子靶向治疗药物的疗效有限。因此,治疗NSCLC的新药仍然是迫切需要的。目前,植物来源的天然药物是抗癌药物的热点领域。五味子酯甲(Schisantherin A,SA)是从中药五味子中分离得到的一种主要的生物活性木脂素。最近的研究显示其具有改善脑功能、减轻肝肾缺血再灌注损伤的保护作用[4-6]。然而,目前尚无文献报道SA对肺癌细胞是否具有抗癌作用及其可能的机制。
长度超过200个核苷酸的长链非编码RNA(LncRNAs)在不同水平的肿瘤过程中扮演了主要角色,包括转录和转录后加工[7]。LncRNA介导致癌或抗癌作用,但许多参与肿瘤发生发展的LncRNA的具体调控机制尚不清楚,需要进一步阐明。PI3K/AKT信号通路是人类肿瘤中最频繁激活的信号通路之一,参与调控肿瘤活性及抗癌药物的耐药性[8-11]。本研究检测了SA对肺癌细胞迁移和侵袭的干预作用,及其对肺癌中LncRNA CCAT1和PI3K/AKT信号通路的影响,旨在探讨SA对肺癌细胞迁移和侵袭的影响,从而为抗癌药物的研发提供基础数据。
1 材料与方法
1.1 材料
1.1.1 细胞系 NSCLC细胞系(ATCC,美国,类型:A549和H1299)。本研究获得西安交通大学第一附属医院医学伦理委员会批准(伦理审批号:XJTU2BE2019JCS-065)。
1.1.2 药物 五味子酯甲(SA)(TargetMol公司,美国,货号:T2913);青霉素-链霉素(Gibco公司,美国,货号:15140163)。
1.1.3 试剂与仪器 胎牛血清(Gibco公司,美国,货号:0014020DJ);DMEM培养基(Gibco公司,美国,货号:A4192101);二甲基亚砜(DMSO)(Sigma公司,美国,货号:D2650);蛋白酶抑制剂(Sigma公司,美国,货号:11206893001);磷酸酶抑制剂(Sigma公司,美国,货号:P2850);细胞计数盒-8(CCK-8)(碧云天生物技术研究所,货号:C0037);结晶紫(碧云天生物技术研究所,货号:C0121);RIPA裂解缓冲液(碧云天生物技术研究所,货号:P0013B);Annexin V-FITC细胞凋亡检测试剂盒(碧云天生物技术研究所,货号:C1062S);Transwell(Corning公司,美国,货号:3428);Matrigel购自(BD Pharmingen公司,美国,货号:BD356234);TRIzol试剂(Invitrogen公司,美国,货号:15596018);PrimeScriptTMRT-PCR试剂盒(Takara公司,日本,货号:RR037A);SYBR Premix Ex TaqTMⅡ(Takara公司,日本,货号:RR820A);BCA试剂盒(Thermo Fisher Science公司,美国,货号:23228);ECL化学发光检测试剂盒(BioRad公司,美国,货号:1705062);p-PI3K(Cell Signaling Technology公司,美国,货号:17366);PI3K(Cell Signaling Technology公司,美国,货号:4257);p-AKT(Cell Signaling Technology公司,美国,货号:4060);AKT(Cell Signaling Technology公司,美国,货号:4691);PTEN(Cell Signaling Technology公司,美国,货号:9188);E-cadherin(Cell Signaling Technology公司,美国,货号:3195);MMP2(Cell Signaling Technology公司,美国,货号:87809);MMP9(Cell Signaling Technology公司,美国,货号:13667);GAPDH(Cell Signaling Technology公司,美國,货号:5174);辣根过氧化物酶标记的抗兔IgG抗体(Cell Signaling Technology公司,美国,货号:7074)。分光光度计(Thermo Scientific公司,美国,Nanodrop 2000型);流式细胞仪(Beckman Coulter公司,美国,FC500型)。实时荧光定量PCR系统(Applied Biosystem公司,美国,ABI 7500型)。
1.2 方法
1.2.1 细胞培养与给药处理 A549和H1299细胞均在添加了10%胎牛血清、100 U/mL青霉素和100 μg/mL链霉素的DMEM培养基中,培养条件为37 ℃、5% CO2。将SA溶解在DMSO中。采用不同剂量的SA(0 μmol/L、5 μmol/L、10 μmol/L、20 μmol/L、50 μmol/L)孵育A549和H1299细胞24 h、48 h和72 h。
1.2.2 细胞增殖检测 采用细胞计数盒-8(CCK-8)检测A549和H1299细胞增殖情况。将细胞置于96孔板中,细胞密度为3×104个/孔。每孔加入10 μL CCK8,37 ℃孵育2 h,按照CCK-8说明书的方法每隔24 h检测各组细胞增殖情况,以DMSO作为阴性对照组。
1.2.3 细胞凋亡分析 根据说明书的操作步骤,收集每组细胞(3×105个细胞)并用PBS洗涤。然后,将细胞转移到试管中,并在400 mL的结合缓冲液中重悬。将Annexin V(5 mL)和碘化丙啶(PI,5 mL)添加到细胞悬液中,并在室温黑暗中孵育15 min。收集细胞并使用FC500流式细胞仪分析。
1.2.4 伤口愈合实验 将3×105个细胞接种在12孔板上。细胞达到80%融合后,用200 μL移液枪尖头做一细胞划痕,用磷酸盐缓冲液冲洗3次,去除脱落细胞,采用剂量为10 μmol/L的SA孵育24 h,分别于0 h和24 h观察并拍照。
1.2.5 细胞侵袭分析 使用Transwell(孔径8 μmol/L)检测细胞侵袭。采用剂量为10 μmol/L的SA处理细胞24 h。收集3×105个细胞并放入含有200 μL无血清培养液的上室中,上室预先用Matrigel涂覆。下室加入600 μL含有10%胎牛血清的培养液。24 h后,用4%多聚甲醛固定侵袭至下室的细胞,用1%结晶紫染色,显微镜下随机选择5个视野计数。
1.2.6 实时定量逆转录聚合酶链反应 使用TRIzol试剂分离细胞总RNA,并通过Nanodrop 2000分光光度计定量。使用PrimeScriptTMRT-PCR试剂盒从1 μg提取的总RNA样品中合成cDNA。使用SYBR Premix Ex TaqTMⅡ在ABI 7500型实时荧光定量PCR系统上进行PCR。使用2-△△Ct方法计算mRNA的相对表达。GAPDH用作CCAT1内部对照。具体的引物序列设计如下:CCAT1,正向,5′-CAGCGAAGGGGTTGAATGCA-3′,反向,5′-ACCTTAGCCCAGGAGCATAC-3′;GAPDH,正向引物,5′-GTGTTCACCTATTACTTGGTCC-3′,反向引物,5′-GTGACTGCTCTATTCTTACCAT-3′。
1.2.7 Western Blotting分析 用RIPA裂解缓冲液提取细胞蛋白,用BCA试剂盒测定蛋白浓度。将等量的蛋白质(50 μg)进行10%十二烷基硫酸钠聚丙烯酰胺凝胶电泳,然后转移至PVDF膜进行条带分离。随后,在含有Tween-20(TBS-T)的Tris缓冲盐水中用10%脱脂牛奶将膜封闭。将膜与p-PI3K(1∶1 000)、PI3K(1∶1 000)、p-AKT(1∶2 000)、AKT(1∶2 000)、PTEN(1∶2 000)、E-cadherin(1∶3 000)、MMP2(1∶3 000)、MMP9(1∶3 000)和GAPDH(1∶2 000)在4 ℃孵育过夜。第二天用0.1% PBS洗涤后,将膜与辣根过氧化物酶标记的山羊抗兔IgG抗体(1∶2 000)一起孵育,并通过ECL化学发光检测试剂盒进行显影。将目的蛋白水平标准化为GAPDH的蛋白水平。
1.3 统计学方法
采用SPSS 21.0统计软件进行数据分析,其中计数资料以百分率表示,采用χ2检验,计量资料以均数±标准差(±s)表示,采用t检验,以P<0.05为差异有统计学意义。
2 结果
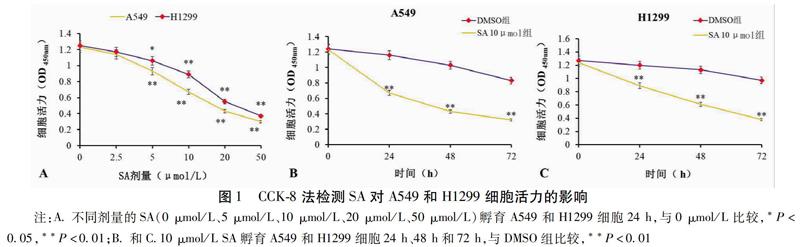
2.1 SA降低肺癌细胞活力
在第24 h,与0 μmol/L比较,5 μmol/L、10 μmol/L、20 μmol/L、50 μmol/L处理的A549和H1299细胞活力显著降低,2种细胞活性均呈剂量依赖性下降(P<0.05)。本研究中10 μmol/L的SA处理A549和H1299细胞24 h均显著降低了细胞活力(P<0.01),因此选择10 μmol/L的SA进行后续实验。用10 μmol/L SA处理2种细胞,在处理24 h、48 h和72 h时,与DMSO组比较,SA 10 μmol/L组的A549和H1299细胞活力均呈时间依赖性下降(P<0.05)。见图1。
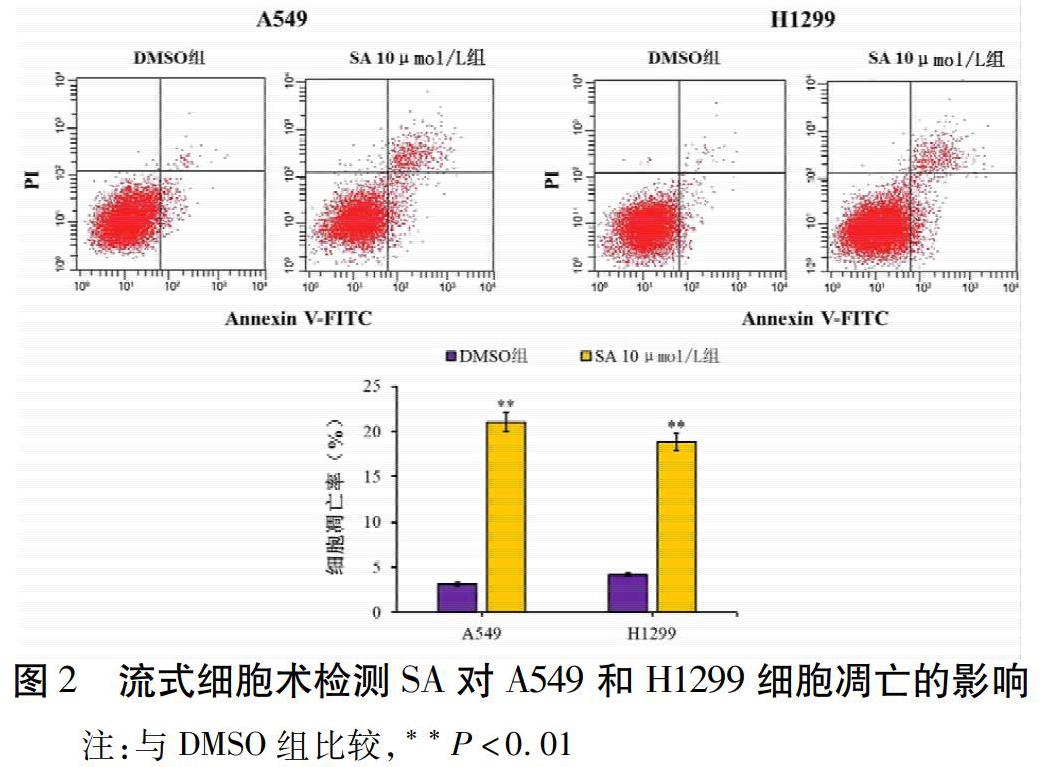
2.2 SA诱导肺癌细胞凋亡
通过流式细胞术检测细胞凋亡,与DMSO组比较,SA 10 μmol/L组的A549和H1299细胞凋亡率显著升高了5.71倍(3.14±0.15%相比21.06±1.13%,t=11.142,P<0.001)和3.48倍(4.21±0.21%相比18.85±0.94%,t=12.336,P<0.001)。见图2。
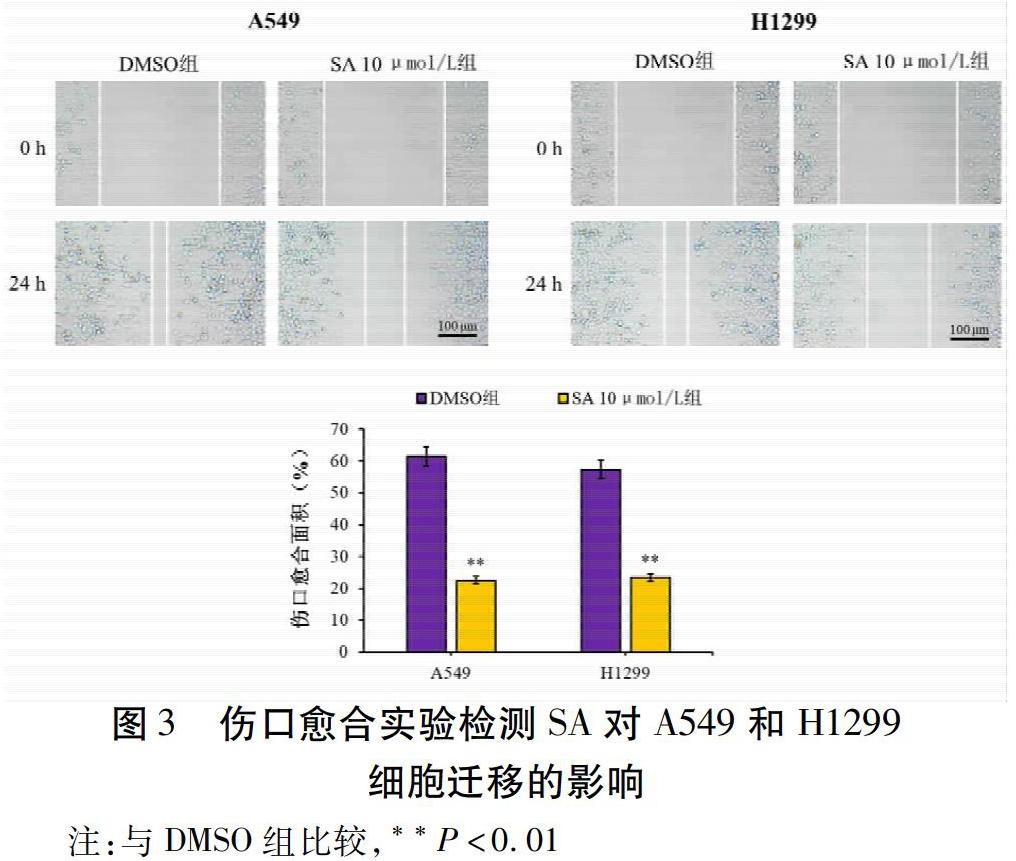
2.3 SA抑制肺癌细胞的迁移和侵袭
通过伤口愈合实验检测细胞迁移能力,与DMSO组比较,SA 10 μmol/L组A549和H1299细胞的伤口愈合面积显著降低了63.42%[(61.32±3.35)%相比(22.43±1.54)%,t=14.352,P<0.001]和59.15%[(57.35±2.65)%相比(23.43±1.06)%,t=14.325,P<0.001]。通过Transwell检测细胞侵袭能力,与DMSO组比较,SA 10 μmol/L组A549和H1299细胞的侵袭细胞数显著降低了53.98%[(387.21±22.07)%相比(178.18±7.80)%,t=9.542,P<0.001]和53.36%[(367.43±19.41)%相比(171.38±8.02)%,t=11.313,P<0.001]。通过Western Blotting检测细胞中E-cadherin、MMP-2和MMP-9的蛋白表达,与DMSO组比较,SA 10 μmol/L组A549细胞中E-cadherin的蛋白表达水平升高了3.15倍,而MMP-2和MMP-9的蛋白表达水平降低了71.43%和51.56%(P<0.001);SA 10 μmol/L组H1299细胞中E-cadherin的蛋白表达水平升高了1.14倍,而MMP-2和MMP-9的蛋白表達水平降低了64.71%和60.66%(P<0.001)。见图3~5。
2.4 SA抑制肺癌细胞中CCAT1的表达
通过qRT-PCR检测细胞中CCAT1的mRNA表达,与DMSO组比较,SA 10 μmol/L组A549和H1299细胞中CCAT1的mRNA表达水平分别降低了73.00%和77.00%(P<0.01)。见图6。
2.5 SA抑制肺癌细胞中PI3K/AKT信号通路
通过Western Blotting检测细胞中PI3K/AKT信号通路关键分子的蛋白表达,与DMSO组比较,SA 10 μmol/L组A549细胞中p-PI3K和p-AKT的蛋白表达水平显著降低了73.21%和70.69%(P<0.01),PTEN的蛋白表达水平显著升高了1.95倍(P<0.01),而PI3K和AKT蛋白表达水平无明显变化(P>0.05)。与DMSO组比较,SA 10 μmol/L组H1299细胞中p-PI3K和p-AKT的蛋白表达水平显著降低了64.18%和73.91%(P<0.01),PTEN的蛋白表达水平显著升高了1.45倍(P<0.01),而PI3K和AKT蛋白表达水平无明显变化(P>0.05)。见图7。
3 讨论
五味子酯甲(Schisantherin A,SA)是从中药五味子中提取的主要药效成分。据报道,SA可以抑制核因子κB和MAPK的活性以发挥其抗炎作用,被认为是一种核因子κB抑制剂[12]。许多研究也证实SA对神经和肝脏损伤有保护作用[5-6]。另外,在治疗癌症方面,Wang等[13]研究显示,SA通过素通过激活ROS/JNK和抑制Nrf2信号通路诱导人胃癌细胞凋亡,并抑制细胞迁移。然而,目前尚无文献报道SA在肺癌中的作用。因此,本研究考察了SA对肺癌细胞的作用及其机制。
本研究结果显示,SA对A549和H1299细胞的活力均有抑制作用,且呈浓度和时间依赖性。此外,SA诱导了A549和H1299细胞的凋亡。上述结果提示,SA可能是一种新型肺癌治疗药物。肿瘤转移涉及许多生物学过程,如细胞黏附、细胞外基质降解、肿瘤细胞迁移和侵袭等。本研究中,SA抑制了A549和H1299细胞的迁移和侵袭。此外,SA处理后,A549和H1299细胞中细胞黏附E-cadherin的表達增加,而基质金属蛋白酶MMP-9和MMP-2的表达水平则降低。这些发现表明SA可以显著抑制肺癌细胞的转移能力。
近年来,LncRNAs已成为肿瘤研究的热点,参与调节肿瘤细胞的增殖、侵袭、迁移、凋亡等生物学过程。LncRNAs的异常表达与癌症的发生发展密切相关。虽然LncRNA参与肿瘤发生发展的具体调控机制尚不清楚,但已证实多种LncRNA促进癌细胞的增殖、迁移、侵袭和转移,其中包括HOX基因的反义基因间RNA、转移相关肺腺癌转录本-1、SLC7A11-AS1、LINC01606等[14-17]。结肠癌相关转录本-1(Colon Cancer Associated Transcript-1,CCAT1)位于染色体8q24.21上,全长2 628个核苷酸,是一种与多种肿瘤相关的致癌基因,可被c-Myc激活,促进癌症的发展[18-20]。CCAT1在各种类型的肿瘤中异常表达并表现出致癌作用,例如胃癌、肝癌和肺癌[21-23]。更重要的是,在耐多西他赛的肺癌细胞中,CCAT1被发现起癌基因的作用并促进其化学耐药性[21]。在胃癌中,敲除Carlo-5基因降低了ERK和MAPK(P38)的磷酸化[24]。有研究证实CCAT1通过激活PI3K/AKT在FTC-133甲状腺癌细胞中发挥致癌作用[25]。本研究发现SA抑制了A549和H1299细胞中CCAT1的表达。因此,SA的对肺癌细胞转移的抑制作用可能是通过抑制CCAT1介导的。
磷脂酰肌醇3-激酶(Phosphatidylinositol 3-kinase,PI3K)是生长因子受体酪氨酸激酶(Receptor Tyrosine Kinase,RTK)下游的主要信号成分,它可以激活AKT,调节一系列调控细胞增殖、成活、生长和其他过程的靶蛋白[9]。PI3K/AKT信号通路是人类肿瘤中最频繁激活的信号通路之一,与肿瘤细胞对特定治疗药物的敏感性或耐药性有关,并参与控制上皮细胞向间充质细胞的转化[10]。PI3K/AKT信号通路具有防止细胞凋亡和促进恶性转化的作用。AKT的激活促进肿瘤转移和侵袭,拮抗细胞周期停滞和血管生成,并使mTOR蛋白激酶磷酸化。据报道,CCAT1作为一种癌症增强子RNA(eRNA),通过与TP63和SOX2以及与EGFR DNA分子相互作用来激活PI3K/AKT信号通路[26]。此外,PI3K/AKT信号通路的活性受RAS的正向调节和PTEN的负向调节[27]。本研究显示,SA抑制了A549和H1299细胞中PI3K和AKT的磷酸化,并上调了抑癌基因PTEN的表达。表明SA通过抑制PI3K/AKT信号通路来抑制肺癌细胞的生长和转移。
总之,本研究表明SA通过抑制CCAT1和PI3K/AKT信号通路抑制肺癌细胞的生长和转移。通过对SA进行结构和化学修饰来提高其溶解性并降低其对正常细胞的毒性,有望将其开发成一种新型抗癌药物。
参考文献
[1]Jonna S,Subramaniam DS.Molecular diagnostics and targeted therapies in non-small cell lung cancer(NSCLC):an update[J].Discov Med,2019,27(148):167-170.
[2]Osmani L,Askin F,Gabrielson E,et al.Current WHO guidelines and the critical role of immunohistochemical markers in the subclassification of non-small cell lung carcinoma(NSCLC):Moving from targeted therapy to immunotherapy[J].Semin Cancer Biol,2018,52(Pt 1):103-109.
[3]Proto C,Ferrara R,Signorelli D,et al.Choosing wisely first line immunotherapy in non-small cell lung cancer(NSCLC):what to add and what to leave out[J].Cancer Treat Rev,2019,75:39-51.
[4]Gong J,Wang X.Schisantherin A protects renal tubular epithelial cells from hypoxia/reoxygenation injury through the activation of PI3K/AKT signaling pathway[J].J Biochem Mol Toxicol,2018,32(7):e22160.
[5]Liu C,Sun W,Li N,et al.Schisantherin A Improves Learning and Memory of Mice with D-Galactose-Induced Learning and Memory Impairment Through Its Antioxidation and Regulation of p19/p53/p21/Cyclin D1/CDK4/RB Gene Expressions[J].J Med Food,2018,21(7):678-688.
[6]Zheng N,Liu F,Lu H,et al.Schisantherin A protects against liver ischemia-reperfusion injury via inhibition of mitogen-activated protein kinase pathway[J].Int Immunopharmacol,2017,47:28-37.
[7]Nasrollahzadeh-Khakiani M,Emadi-Baygi M,Schulz WA,et al.Long noncoding RNAs in gastric cancer carcinogenesis and metastasis[J].Brief Funct Genomics,2017,16(3):129-145.
[8]Alzahrani AS.PI3K/Akt/mTOR inhibitors in cancer:At the bench and bedside[J].Semin Cancer Biol,2019,59:125-132.
[9]Costa R,Han HS,Gradishar WJ.Targeting the PI3K/AKT/mTOR pathway in triple-negative breast cancer:a review[J].Breast Cancer Res Treat,2018,169(3):397-406.
[10]Hoxhaj G,Manning BD.The PI3K-AKT network at the interface of oncogenic signalling and cancer metabolism[J].Nat Rev Cancer,2020,20(2):74-88.
[11]O′Donnell JS,Massi D,Teng M,et al.PI3K-AKT-mTOR inhibition in cancer immunotherapy,redux[J].Semin Cancer Biol,2018,48:91-103.
[12]Liao S,Zhou K,Li D,et al.Schisantherin A suppresses interleukin-1β-induced inflammation in human chondrocytes via inhibition of NF-κB and MAPKs activation[J].Eur J Pharmacol,2016,780:65-70.
[13]Wang Z,Yu K,Hu Y,et al.Schisantherin A induces cell apoptosis through ROS/JNK signaling pathway in human gastric cancer cells[J].Biochem Pharmacol,2020,173:113673.
[14]Chiyomaru T,Fukuhara S,Saini S,et al.Long non-coding RNA HOTAIR is targeted and regulated by miR-141 in human cancer cells[J].J Biol Chem,2014,289(18):12550-12565.
[15]Hirata H,Hinoda Y,Shahryari V,et al.Long Noncoding RNA MALAT1 Promotes Aggressive Renal Cell Carcinoma through Ezh2 and Interacts with miR-205[J].Cancer Res,2015,75(7):1322-1331.
[16]Luo Y,Tan W,Jia W,et al.The long non-coding RNA LINC01606 contributes to the metastasis and invasion of human gastric cancer and is associated with Wnt/β-catenin signaling[J].Int J Biochem Cell Biol,2018,103:125-134.
[17]Luo Y,Wang C,Yong P,et al.Decreased expression of the long non-coding RNA SLC7A11-AS1 predicts poor prognosis and promotes tumor growth in gastric cancer[J].Oncotarget,2017,8(68):112530-112549.
[18]Coni P,Madeddu A,Kuqi L,et al.LncRNA colon cancer-associated transcript 1(CCAT1) in ovarian cancer[J].Eur Rev Med Pharmacol Sci,2018,22(6):1525-1527.
[19]Lai XJ,Cheng HF.LncRNA colon cancer-associated transcript 1(CCAT1) promotes proliferation and metastasis of ovarian cancer via miR-1290[J].Eur Rev Med Pharmacol Sci,2018,22(2):322-328.
[20]Yang C,Pan Y,Deng SP.Downregulation of lncRNA CCAT1 enhances 5-fluorouracil sensitivity in human colon cancer cells[J].BMC Mol Cell Biol,2019,20(1):9-19.
[21]Chen J,Zhang K,Song H,et al.Long noncoding RNA CCAT1 acts as an oncogene and promotes chemoresistance in docetaxel-resistant lung adenocarcinoma cells[J].Oncotarget,2016,7(38):62474-62489.
[22]Deng L,Yang SB,Xu FF,et al.Long noncoding RNA CCAT1 promotes hepatocellular carcinoma progression by functioning as let-7 sponge[J].J Exp Clin Cancer Res,2015,34(1):18-27.
[23]Yang F,Xue X,Bi J,et al.Long noncoding RNA CCAT1,which could be activated by c-Myc,promotes the progression of gastric carcinoma[J].J Cancer Res Clin Oncol,2013,139(3):437-445.
[24]Zhang Y,Ma M,Liu W,et al.Enhanced expression of long noncoding RNA CARLo-5 is associated with the development of gastric cancer[J].Int J Clin Exp Pathol,2014,7(12):8471-8479.
[25]Yang T,Zhai H,Yan R,et al.lncRNA CCAT1 promotes cell proliferation,migration,and invasion by down-regulation of miR-143 in FTC-133 thyroid carcinoma cell line[J].Braz J Med Biol Res,2018,51(6):e7046.
[26]Jiang Y,Jiang YY,Xie JJ,et al.Co-activation of super-enhancer-driven CCAT1 by TP63 and SOX2 promotes squamous cancer progression[J].Nat Commun,2018,9(1):3619-3621.
[27]Stambolic V,Suzuki A,de la Pompa JL,et al.Negative regulation of PKB/Akt-dependent cell survival by the tumor suppressor PTEN[J].Cell,1998,95(1):29-39.
(2020-11-19收稿 責任编辑:吴珊,徐颖)
猜你喜欢
中国典型病例大全(2022年11期)2022-05-13
中国药学药品知识仓库(2022年1期)2022-03-23
广东教育·职教版(2021年3期)2021-04-20
养生阅刊(2020年7期)2020-08-16
分析化学(2019年3期)2019-03-30
中国信息化周报(2019年8期)2019-03-27
饮食与健康·下旬刊(2019年10期)2019-03-09
学校教育研究(2018年27期)2018-05-14
中老年健康(2016年12期)2017-01-18
销售与管理(2006年9期)2006-09-17
