
白茶萎凋过程中差异基因的生物信息学分析与表达分析
2021-08-31 09:41陈静陈桂信叶乃兴黄先洲潘玉华杨晓滨
农业与技术 2021年16期
陈静陈桂信叶乃兴黄先洲潘玉华杨晓滨
(1.宁德职业技术学院生物技术系,福建 福安 355000;2.福建农林大学园艺学院,福建 福州 350002)
白茶萎凋工序是水分逐步散失过程,细胞渗透调节作用下,细胞膜渗透性发生明显改变,引起生化及内质成分的变化,各特征性次生代谢产物酚类物质[1-4]、咖啡碱[5-7]、茶氨酸[8,9]、萜烯类香气物质[10-12]代谢途径中的关键酶及相关基因共同决定白茶品质形成,已陆续在NCBI数据库登录相关基因的序列信息及相关功能注释。本研究从白茶萎凋芽叶-新鲜芽叶正向文库中分离获得了S-腺苷甲硫氨酸脱羧酶(SAMDC)、琥珀酸脱氢酶(SDH)、磷酸-2-脱氢-3-脱氧庚糖酸醛缩酶(DAHPS),并对基因序列进行了生物信息学分析,为以后研究白茶品质形成分子形成机理奠定了基础。
1 材料与方法
1.1 材料
供试品种为国家级审定茶树良种“福鼎大白茶”,种植于福建农林大学南区茶学教学基地种质资源圃,按传统白茶加工工艺进行萎凋,室内自然萎凋温度22~25℃,空气相对湿度70%~75%,分别称取0.1000g的鲜叶(萎凋处理0h)和萎凋处理4h、8h、16h、24h、32h、40h、48h、52h的叶片,共9个样品,设3次重复,经液氮速冻后置-80℃冰箱保存备用。
1.2 试验方法
1.2.1 白茶萎凋叶与鲜叶正向抑制差减杂交cDNA文库的构建
各取萎凋芽叶与新鲜芽叶样品总RNA为模板,参考俞滢[13]的方法构建文库。
1.2.2 白茶萎凋叶与鲜叶差异基因阳性克隆的筛选
反向Northern-blotting中探针的制备、点膜、预杂交、杂交、显色等步骤,参照俞滢[13]的方法和Roche公司的DIG High Prime Labeling and Detection Starter Kit I(Version 13)试剂盒的说明书进行,根据反向Northern-blotting的结果,挑取阳性克隆进行培养,用于测序。
经反向Northern Bloting显色验证出的阳性克隆所对应菌液送上海祥音公司测序。利用在线NCBI vecscreen剔除载体序列、删除污染序列和低于并结合300 bp的序列;利用DNAMAN软件对有效序列进行拼接,将测序结果生成fasta文件以供后期生物信息学分析;将测序结果与NCBI核酸数据库和拟南芥蛋白数据库结合进行Blast N比对分析,对基因功能进行注释和归类分析。
1.2.4 白茶品质形成的相关差异基因的生物信息学分析
利用NCBI数据库对阳性克隆的测序结果,见表1,所获得的核苷酸序列和氨基酸序列进行Blast X比对;阳性克隆基因的推测功能使用Protparam在线软件;预测阳性克隆基因所编码的蛋白质二级结构使用SOPMA在线软件,找出与功能密切相关的结构域;系统进化树在MEGA软件中形成以进行聚类分析。

表1 3个白茶品质形成相关基因信息表
1.2.5 白茶品质形成的相关基因在不同萎凋时期的表达量变化
利用Primer Premier 5和Beacon Desiner7软件设计荧光定量PCR引物,见表2。
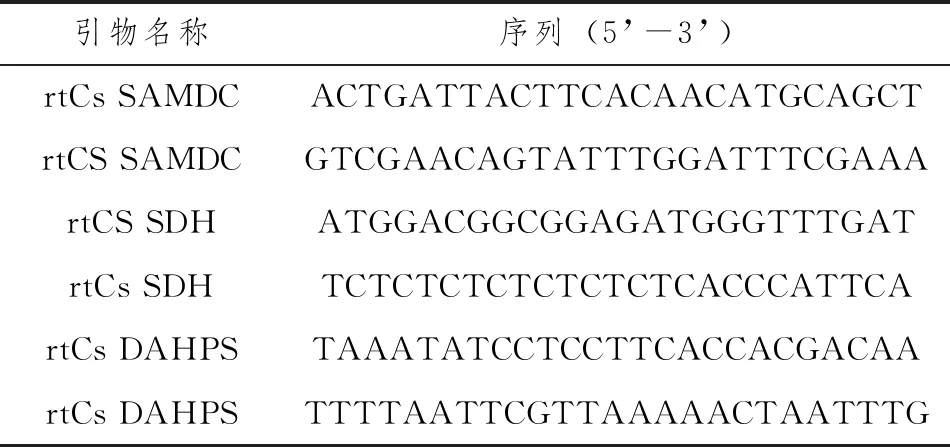
表2 荧光定量PCR引物序列
2 结果与分析
2.1 白茶Cs SAMDC基因的生物信息学分析与表达
2.1.1 白茶Cs SAMDC基因的生物信息学分析
S-腺苷甲硫氨酸脱羧酶是合成亚精胺的重要酶类。S-腺苷甲硫氨酸经过S-腺苷甲硫氨酸脱羧酶(SAMDC)催化,生成脱羧SAM,由Put与脱羧的SAM提供的氨丙基合成Spd和Spm,合成亚精胺合成酶和精胺合成酶,目前已在许多植物中克隆到了SAMDC基因,如拟南芥[14]、棉花[15]、番茄[16]、杜梨[17]等。研究表明,SAMDC基因是由一个多基因家族所编码[18],每一种植物中的SAMDC基因都不止一个,如拟南芥中有4个SAMDC基因[19]、水稻中有4个SAMDC基因[109]、番茄中有3个SAMDC基因[16]。
测序结果,见图3,白茶Cs SAMDC核苷酸序列长度为1355bp,该基因编码的蛋白由334个氨基酸组成,含有SAMDC_redism_C superfamily结构域和活性位点,见图1,相对分子量为38988.23,等电点为5.29,不稳定系数为48.55,属于不稳定脂溶蛋白,在蛋白质二级结构中,28.19%为α螺旋,23.44%为延伸链,7.42%为β转角,40.95%为无规则卷曲。
治愈:治疗三天,患儿没有腹泻症状,精神状态良好;有效:治疗三天,患儿没有呕吐脱水症状,大便次数减少;无效:治疗三天没有任何改善或更加严重。
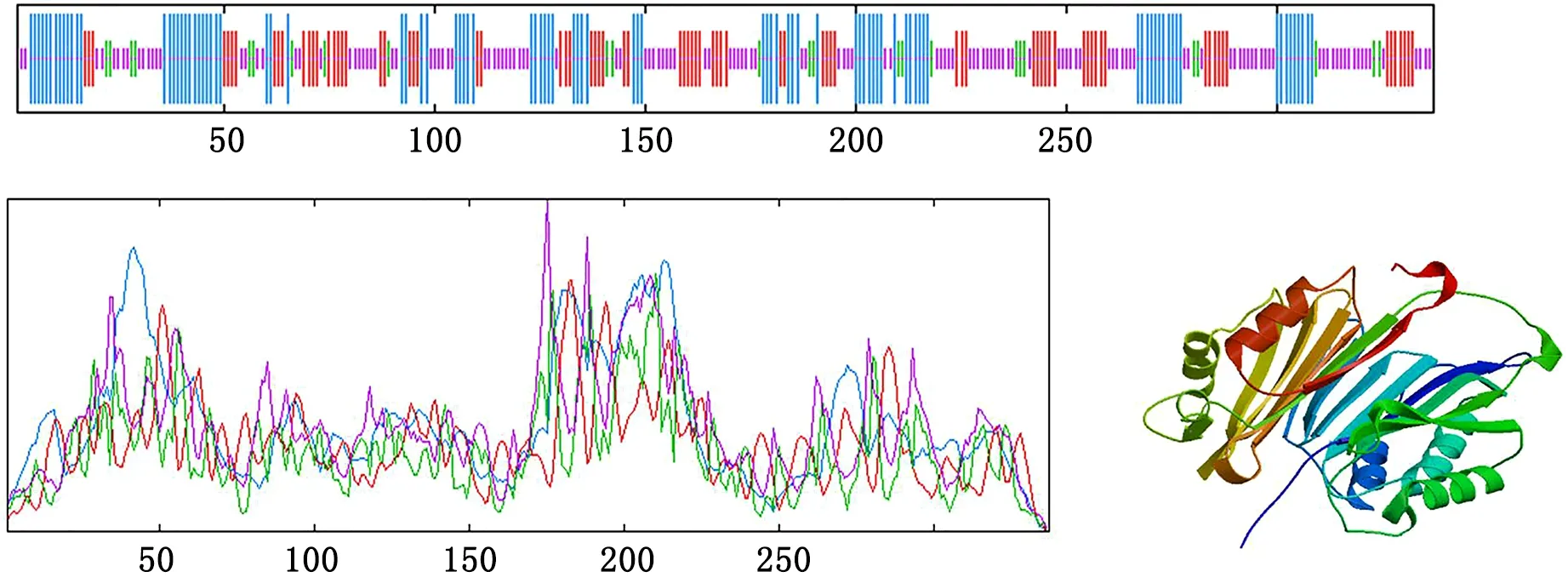
图1 Cs SAMDC氨基酸结构预测

图2 Cs SAMDC序列的NJ 系统进化树

图3 Cs SAMDC基因的核苷酸序列及推导的氨基酸序列
将Cs SAMDC基因的氨基酸序列,在NCBI数据库中进行BLAST比对,结果表明,该基因与茶树(Camellia sinensis)的同源性为87%、与香附子(Cynodon dactylon)、小麦(Triticum aestivum)、菜豆(Phaseolus vulgaris)、大豆(Glycine max)同源性均为100%。使用MEGA进行NJ系统进化树分析,结果见图2。
2.1.2 白茶Cs SAMDC基因的表达分析
以GAPDH为内参基因,采用荧光定量PCR对白茶萎凋过程中Cs SAMDC基因的相对表达量进行分析,见图4,结果表明,不同萎凋历时下,萎凋前期即萎凋历时0~16h,萎凋后期即萎凋历时24~52h,2个阶段Cs SAMDC呈现先上升后下降的趋势,在萎凋历时4h的相对表达量是最高的,是0 h的4倍。
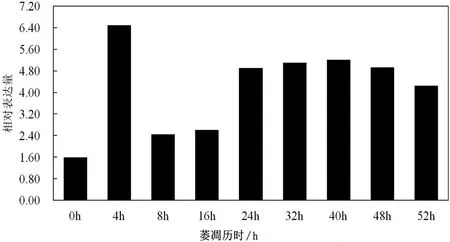
图4 白茶不同萎凋历时中Cs SAMDC基因的表达分析
2.2 白茶Cs SDH基因的生物信息学分析与表达
2.2.1 白茶Cs SDH基因的生物信息学分析
琥珀酸脱氢酶是参与三羧酸循环-氧化磷酸化的关键酶基因。
测序结果见图7,白茶Cs SDH基因的cDNA核苷酸序列长度为1224bp,包含17 bp的5/-UTR、57 bp的3/-UTR和1150bp的ORF,起始密码子为ATG、终止密码子为TGA;该基因编码的蛋白由388个氨基酸组成,见图5,相对分子量为43239.04,等电点为6.45,不稳定系数为32.15,属于不稳定脂溶蛋白,在蛋白质二级结构中,29.70 %为α螺旋,25.13%为延伸链,13.96%为β转角,31.22 %为无规则卷曲。
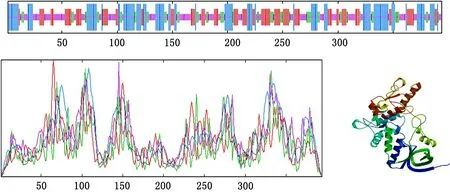
图5 Cs SDH氨基酸结构预测

图6 Cs SDH序列的NJ 系统进化树

图7 Cs SDH的核苷酸序列及推导的氨基酸序列
将Cs SDH基因的氨基酸序列,在NCBI数据库中进行BLAST比对,结果表明,该基因与荔枝(Litchi chinensis)、马尾松(Pinus massoniana)、葡萄(Vitis vinifera)、油棕(Elaeis guineensis)的同源性均为100%。使用MEGA进行NJ系统进化树分析结果表明,Cs SDH基因与玉米的SDH基因属于同一分支,见图6。
2.2.2 白茶Cs SDH基因的表达分析
以GAPDH为内参基因,采用荧光定量PCR对白茶萎凋过程中Cs SDH基因的相对表达量进行分析,结果表明,不同萎凋历时下,萎凋历时0~52h,Cs SDH含量呈现缓慢下降的趋势,变化差异不大,在萎凋历时0h的相对表达量是最高的,是52h的6倍。
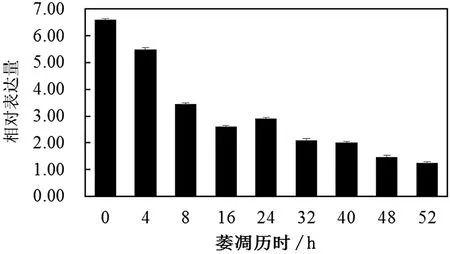
图8 白茶不同萎凋历时中CsSDH基因的表达分析
2.3 白茶Cs DAHPS基因的生物信息学分析与表达
2.3.1 白茶CsDAHPS基因的生物信息学分析
莽草酸途径是芳香族氨基酸的生物合成途径,同时也控制着芳香族次生代谢产物的产出,连接糖代谢和次生代谢的纽带。磷酸-2-脱氢-3-脱氧庚糖酸醛缩酶是莽草酸途径7个酶促反应中的第1个酶类,是参与次级代谢的主要蛋白。
测序结果见图11,白茶Cs DAHPS基因的cDNA全长为1935 bp,核苷酸序列长度为1682bp,包含65bp的5/-UTR、58bp的3/-UTR和1812bp的ORF,起始密码子为ATG、终止密码子为TGA;该基因编码的蛋白由538个氨基酸组成,含有异戊二烯结构域Isoprenoid_Biosyn_C1 superfamily,见图9,相对分子量为62021.74,等电点为8.82,不稳定系数为42.90,属于不稳定脂溶蛋白,在蛋白质二级结构中,35.36%为α螺旋,16.25%为延伸链,7.32%为β转角,41.07 %为无规则卷曲。

图9 Cs DAHPS氨基酸结构预测

图10 Cs DAHPS序列的NJ 系统进化树
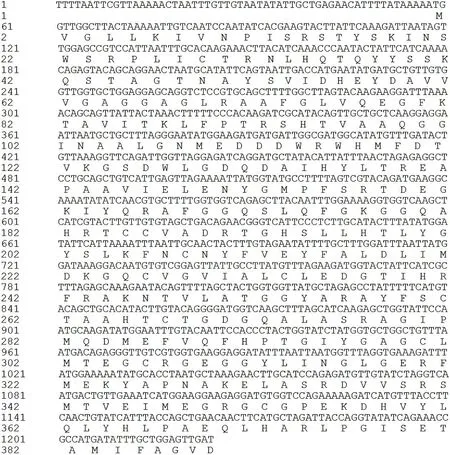
图11 Cs DAHPS的核苷酸序列及推导的氨基酸序列
将Cs DAHPS基因的氨基酸序列,在NCBI数据库中进行BLAST比对,结果表明,该基因与番茄(Solanum lycopersicum)、番薯紫竹(Ipomoea purpurea)、可可(Theobroma cacao)的同源性均为100%,见图10。
2.3.2 白茶Cs DAHPS基因的表达分析
以GAPDH为内参基因,采用荧光定量PCR对白茶萎凋过程中Cs DAHPS基因的相对表达量进行分析,结果表明,萎凋历时0~24h,Cs DAHPS含量呈现缓慢下降的趋势,变化差异不大,在萎凋历时32h的相对表达量是最高的,是52h的5倍。
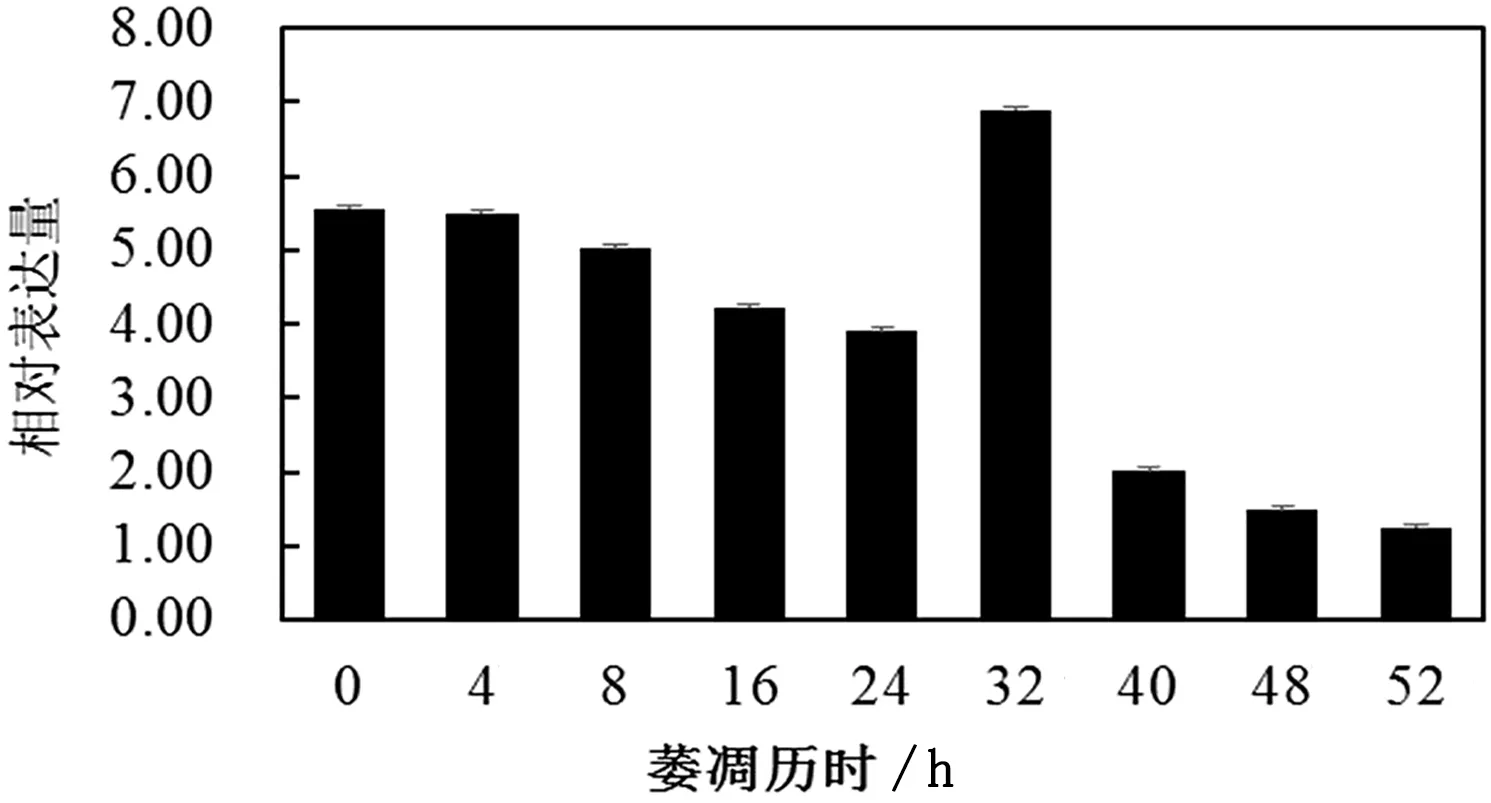
图12 白茶不同萎凋历时中CsDAHPS基因的表达分析
3 结论与讨论
3.1 白茶萎凋过程品质形成与逆境的关系
白茶萎凋过程中水分及酶活性不断变化,本身就是失水条件下的逆境,因此形成与水分散失条件下相关的基因,本研究分离的白茶Cs SAMDC合成亚精胺的重要酶类家族,BLAST比对结果表明,Cs SAMDC编码的蛋白在氨基酸序列上与茶树、菜豆等植物的SAMDC基因具有较高的同源性,两者均达到85%以上。从图2中Cs SAMDC的系统进化树可知,白茶Cs SAMDC属于SAMDC家族,与山茶属茶树形成一个分类群,可以初步确定Cs SAMDC是编码白茶的S-腺苷甲硫氨酸脱羧酶的同源基因。S-腺苷甲硫氨酸脱羧酶是合成亚精胺主要限速酶基因。白茶在萎凋历时4h,SAMDC酶表达量最高,说明体内可能产生大量的亚精胺等物质,通过渗透调节途径来增强能力,之后趋于平衡,可能是渗透调节达到一个平稳饱和阶段,有待进一步研究。
3.2 白茶萎凋过程品质形成与三羧酸循环的关系
白茶萎凋过程是各类酶系活跃的阶段,其中白茶Cs SDH是三羧酸循环关键限速酶基因,其表达量呈现缓慢下降的趋势,可初步判定萎凋失水的含量变化是由高至低,因此Cs SDH在萎凋过程中呈现表达逐渐下调趋势,这与孙跃进[20]的结果相一致,萎凋后期糖酵解途径加强,三羧酸循环减弱。
3.3 白茶萎凋过程品质形成与莽草酸循环的关系
莽草酸循环是芳香族氨基酸的生物合成途径,这些芳香族氨基酸除了用来合成蛋白质外,还作为前提生成种类繁多的具有芳香环结构的次级代谢产物,如木质素、维生素E、花青素、植物抗毒素、吲哚乙酸酯、紫草素、水杨酸、丹宁酸、肉桂酸、黄酮及黄酮类等[21-25]。王芳等[26]研究表明,室内自然萎凋过程中不同白茶品种的谷氨酸脱羧酶(GAD)性整体变化趋势基本一致,即萎凋前期的茶叶GAD活性随谷氨酸(GLU)含量的增加而增强,催化GLU转化为γ-氨基丁酸(GABA),但随着萎凋叶失水程度的增加,催化成GABA的能力下降,且GABA的降解速度大于生成速度,故其含量在萎凋中后期呈缓慢下降趋势。作为莽草酸途径的第1个酶,白茶Cs DAHPS 在萎凋历时0~24h呈现下调表达,萎凋历时40~52h呈现下调表达,这与王芳的结果基本一致。但在萎凋历时32h呈现最大值,这一结果可进一步研究。
猜你喜欢
天津市教科院学报(2021年5期)2021-11-10
生物学通报(2021年9期)2021-07-01
汉字汉语研究(2021年1期)2021-06-11
汉字汉语研究(2021年1期)2021-06-11
读者·原创版(2020年8期)2020-08-17
红楼梦学刊(2019年5期)2019-04-13
汉字汉语研究(2018年3期)2018-11-06
汉语世界(2017年4期)2017-03-07
淮阴工学院学报(2014年5期)2014-09-10
