
基于环境DNA宏条形码技术的北京地区鱼类多样性调查和外来鱼种入侵风险评估
2021-09-02 12:25范仲儒陈义永
生物安全学报 2021年3期
刘 波, 王 浩,, 秦 斌, 范仲儒, 熊 薇, 陈义永*
1北京市水文总站,北京 100089;2中国科学院生态环境研究中心,北京 100085
鱼类是水生生态系统中不可替代的重要组成部分,对于水生态环境健康与平衡、水资源保护和人类生活都具有重要价值(张姗,2020;Holmlund & Hammer,1999)。随着全球化进程的加快和人类活动的加剧,全球淡水鱼类多样性和种群数量急剧下降,淡水鱼类成为受威胁程度最严重的类群之一(Dudgeonetal.,2006;Guoetal.,2019)。根据2020年世界自然保护联盟(International Union for Conservation of Nature,IUCN)脊椎动物濒危物种评估结果,被列入红色名录的濒危鱼类高达2721种,而且近10年来濒危鱼类物种数目一直排在脊椎动物类群之首。目前,淡水鱼类面临的威胁主要集中在水体污染、过度捕捞、外来物种入侵、栖息地丧失与破碎化以及城市化进程等(Dudgeonetal.,2006;Vörösmartyetal.,2010)。现今,包括水产养殖、压舱水、船壳污损以及观赏动植物的释放等频繁的人类活动,导致越来越多的外来生物被引入水生生态系统中,而外来物种的大量引入又进一步加剧了人类活动对水生生态系统的结构、功能的影响(Dudgeonetal.,2006;Strayer,2010)。相比于其他生态系统,淡水生态系统更容易受入侵物种的影响,其中外来入侵鱼类被认为是造成水生鱼类多样性下降的主要原因(Haubrocketal.,2020)。据报道,意大利阿尔诺河鱼类群落在2个世纪内发生完全改变,土著鱼类已几乎完全被外来鱼类所替代(1800年:92%土著鱼种;2015年:94%外来鱼种)。北京地区河湖纵横,境内分布有425条河流、41个湖泊、88座水库,淡水鱼类资源比较丰富(潘术香,2004)。资料显示,北京地区共有93种鱼类,隶属于13目23科73属,物种数占全国淡水鱼类总数的7.03%,其中本地原生鱼类历史记录为83种(张春光等,2011)。但自20世纪中叶以来,北京原生鱼种数目骤减,外来鱼种数大幅增加,这导致北京地区鱼类多样性和群落结构发生改变(张春光等,2011)。2010年北京鱼类调查只发现43种本地原生鱼类,外来鱼种在北京许多水体中已成为优势种(张春光等,2011)。这表明北京地区水生态系统中鱼类资源面临巨大威胁,鱼类保护和恢复工作刻不容缓。因此,为保护北京鱼类多样性亟需开展鱼类调查和评估工作。
建立快速、高效的鱼类多样性监测方法是鱼类保护和管理的关键(Reesetal.,2014)。传统鱼类多样性调查方法主要利用拖网、电捕、地笼等捕捞方式,通过对渔获物进行形态学鉴定和计数称重,识别鱼类物种种类并统计丰度与生物量(刘焕章等,2016;张姗,2020)。传统方法生态损伤性大、工作量大、灵敏度低、对鉴定人员要求高,直接影响鱼类鉴定的准确度和效率,为加强鱼类保护和管理,需要建立集快速、高效、灵敏和高通量于一体的新型鱼类多样性监测方法。新兴的环境DNA宏条形码(environmental DNA metabarcoding, eDNA metabarcoding)技术可直接从环境样品(如水、土壤、空气)中提取DNA,利用针对目标类群设计的引物,通过PCR扩增和高通量测序(high-throughput sequencing,HTS),实现对环境样本中多个目标物种的识别鉴定(陈星宇等,2020;Piperetal.,2019)。基于环境DNA技术的鱼类监测方法采用非损伤性的取样方式,操作步骤简单高效、灵敏度高,且不要求检测人员掌握完备的形态分类学知识,因此该技术在鱼类多样性监测和评估方面具有极大的应用潜能(Reesetal.,2014;Thomsen & Willerslev,2015)。近年来,环境DNA技术广泛应用于物种组成和分布调查、外来入侵物种检测及濒危物种监测等(张姗,2020)。高通量测序技术的快速发展使得利用环境DNA方法开展大规模生物群落物种多样性的调查监测成为可能(Thomsen & Willerslev,2015)。外来物种在入侵初期群体规模较小,是对其进行控制和消灭的关键时期,但是入侵初期其种群密度极低,从复杂群落中检测出这些低丰度目标生物成为构建早期检/监测和预警体系的关键,环境DNA方法在水生外来入侵生物的早期监测与预警中具有广阔的应用前景(李晗溪等,2019)。例如,Evansetal.(2016)借助环境DNA方法从水库中检测出以前传统方法监测到的所有鱼类,并监测到一些以前从未检测到的鱼类物种。目前,国内利用环境DNA宏条形码技术进行鱼类多样性的调查研究相对欠缺(舒璐等,2020;Zhangetal.,2019)。
本研究利用环境DNA宏条形码技术对北京地区不同水体类型中鱼类多样性和群落结构进行监测和分析,并以监测结果为基础,汇总了目前北京地区水生态系统的本地鱼种和外来鱼种,评估了典型外来入侵鱼种的入侵风险。
1 材料与方法
1.1 采样点设置与水样采集
本研究监测工作于2020年6月10—17日进行,设置6个水库(RE)采样点、7个湖泊(LA)采样点和20个河流(RI)采样点,共计33个点位(S34~S66;图1)。利用取水器在每个样点采集1 L的表层环境水样,倒入干净的广口瓶中,并立即置于车载冰箱中低温保存。

图1 北京地区采样点分布图
1.2 环境DNA提取、PCR扩增与高通量测序
1.2.1 环境DNA富集 水样在采集当天运回实验室3 h内完成抽滤工作,利用0.45 μm的混合纤维滤膜对采集水样进行真空抽滤以富集环境DNA,每份样品过滤后的滤膜立即冷冻保存(-20 ℃)。
1.2.2 环境DNA提取 参照Zhangetal.(2020b)的方法,使用DNA提取试剂盒DNeasy Blood & Tissue Kit(Qiagen,69506)对滤膜上的DNA进行提取,利用NanoDrop 2000(Thermo Scientific,Waltham,MA,USA)检测DNA的浓度和质量后,将每份样品所得的环境DNA溶液迅速置于-80 ℃低温保存,直至下一步PCR扩增。
1.2.3 环境DNA扩增与建库测序 本研究选择淡水鱼类线粒体12S通用宏条形码扩增引物:Fish-F1/R1.1(杨江华和张效伟,2019)。此引物已证实对淡水鱼类具有扩增效率高、灵敏度高、群落覆盖度广的特点,且扩增PCR产物长度(约180 bp)适合高通量测序平台,可作为淡水鱼类环境DNA宏条形码监测方法的理想引物。正反向引物的序列分别是:Fish_F1-TCGTGCCAGCCACCGCGGTTA,Fish_R1.1-ATAGTGGGGTATCTAATCCCAG。50 μL PCR扩增体系如下:5 μL的10×buffer+,4 μL的dNTPs,正反向引物各3、0.4 μL的TaKaRa Taq酶,30.6 μL的ddH2O,4 μL的模板DNA(10 ng·μL-1)。PCR扩增程序为:95 ℃预变性5 min;35个循环包括:95 ℃变性30 s,58 ℃退火30 s,72 ℃延伸30 s;最后72 ℃延伸10 min。为了混样建库并进行高通量测序,在引物的5′端加入序列各异的8个核苷酸作为标签(barcode)以区分不同样品,共设计了19对带特异性标签的引物,因此本研究中33个样品只需要构建2个文库即可。每份样品重复PCR扩增3次,将同一样品的PCR产物混合后,利用1.5%的琼脂凝胶电泳进行条带检测,再利用柱式PCR产物纯化试剂盒(Sangon Biotech,Shanghai,China)纯化PCR产物,将纯化后的PCR产物等量混合,完成测序文库的构建并利用Illumina NovaSeq测序平台进行PE150双端测序。
1.3 数据分析
1.3.1 高通量测序数据处理 利用USEARCH软件对高通量测序后的原始序列进行拼接、滤错、序列剪齐、去重复,借助UNOISE算法的unoise3命令进行聚类,得到ZOTU(zero-radius OTU)和对应的序列信息,获取OTU table文件。利用Seed软件的BLASTn功能将所有OTU对应的序列与NCBI网站上下载的鱼类本地数据库进行比对,得到每个OTU的分类界元。其中,比对结果筛选标准为:E值<10-70,最小覆盖度(coverage)>80%,序列相似度(identity)>85%(Yangetal.,2019)。为了降低假阳性,最终只保留至少3条序列的淡水鱼类物种用于后续分析。
1.3.2 鱼类多样性分析 为分析比较不同采样点和水体类型间鱼类多样性水平,利用PRIMER 6.0软件计算Margalef物种丰富度(d)、香农多样性(H′)和Pielou均匀度(J)等多样性指数,对3种水体类型的鱼类物种多样性差异进行Wilcoxon秩和检验(p=0.05),利用每个样本中OTU相对丰度信息计算样本间Bray-Curtis距离,进行主坐标PCOA计算和作图,并结合ANOSIM分析检验不同区域群落之间的相似性。在进行PCOA分析前,物种数据采用平方根标准化以增加数据的正态性。此外,利用优势度公式Y=Ni×f/N(陈义永等,2018)识别3种水体类型的优势鱼种,当优势度值Y≥0.02时,该鱼种为该区域的优势鱼种。
1.3.3 本地和外来鱼种的分类汇总及入侵风险评估 参照张姗(2020)汇总的北京地区1984—2020年北京淡水鱼类和历史记载,将本研究监测得到鱼类物种与之比较,以鉴定区分本地种和非本地种,重点关注非本地种中的外来物种及入侵物种。入侵物种信息参照全球入侵生物数据库(GISD:http:∥www.iucngisd.org/gisd/)、水生外来生物数据库(AquaNIS:http:∥www.corpi.ku.lt/databases/index.php/aquanis/)、中国外来入侵物种数据库(http:∥www.chinaias.cn/wjPart/index.aspx)及中国外来/入侵鱼类相关文献(Chenetal.,2017;Linetal.,2015;Liuetal.,2017;Xiongetal.,2015)。本研究利用水生鱼类入侵风险评估指标体系(freshwater fish invasiveness screening kit,FISK V2;www.cefas.co.uk/nns/tools/;Coppetal.,2016)对外来鱼类在北京地区的入侵风险进行评估。FISK V2指标体系包括鱼类的生物地理/历史、生物/生态和气候变化等相关的55个指标,依据最终得分结果进行入侵风险等级划分。
2 结果与分析
2.1 北京地区鱼类物种多样性与群落组成
以本研究环境DNA技术监测结果为基础,通过对2个高通量测序文库的原始序列进行质量过滤和OTU聚类,共获得5530个原始OTU,其中3311个OTU成功比对到构建的淡水鱼类本地数据库中,最终物种注释得到52种淡水鱼类,隶属于7目22科43属。目级水平比较,鲤形目Cypriniformes种类最多,共计29种,约占鱼类总数的55.77%;其次是鲈形目Perciformes 12种,占23.08%。科级水平比较,鲤科Cyprinidae种类最多,共计18种,占34.62%;其次是鳅科Cobitidae 5种,占9.62%;沙塘鳢科Odontobutidae 4种,占7.69%;其余各科物种数目都较少(1~3种)。所有样点的相对生物量(物种序列丰度)最高的前5种鱼类分别是:鲫CarassiusauratusL.、麦穗鱼Pseudorasboraparva(Temminck & Schlegel)、泥鳅Misgurnusanguillicaudatus(Cantor)、青鳉Oryziaslatipes(Genetics)和饰妆铠弓鱼Chitalaornata(Gray)(表1)。此外,鳙Aristichthysnobilis(Rich)、鲫、草鱼Ctenopharyngodonidella(Val.)、鲤CyprinuscarpioL.、泥鳅、大鳞泥鳅Misgurnusmizolepis(Günther)、青鳉、大鳞副泥鳅ParamisgurnusdabryanusSauvage、麦穗鱼、饰妆铠弓鱼和细鳞裂腹鱼Schizothoraxchongi(Fang)等11种鱼在所有样点中均检测到,且多达26种鱼在超过一半的样点中被检测到。
鱼类数目最多的样点是北海(S39,36种)和玉渊潭西湖(S40,36种),圆明园福海(S38,17种)的鱼种数目最少。各样点香农多样性指数(H′)变化范围是1.49~2.22,最高的样点是高碑店湖(S43),最低的样点是后蔺沟(S48),说明不同样点的鱼类序列相对丰度(图2)。此外,基于物种OTU的PCOA聚类结果显示,不同样点鱼类群落结构呈现一定差异性(图3)。上述结果表明,北京地区不同样点间物种多样性和群落结构呈现一定的空间异质性。

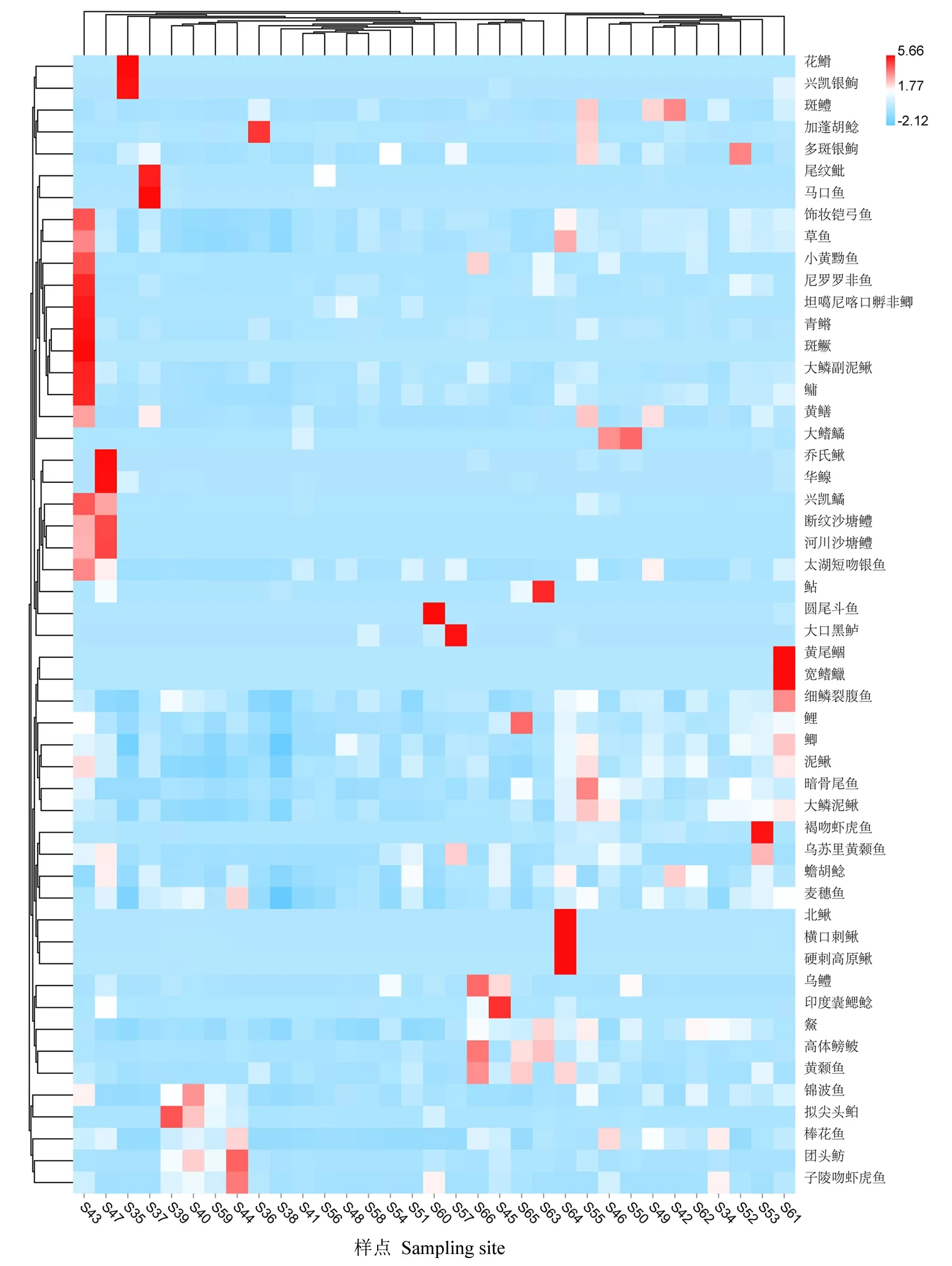
图2 不同采样点鱼类物种相对序列丰度的热图
为进一步探究不同水体类型中鱼类多样性和群落结构是否存在差异,本研究中所有样点划分为水库、湖泊和河流3种水体类型。根据3种水体类型的物种多样性分析箱式图显示,3种水体类型的鱼类物种多样性并没有显著差异(图4)。就Margalef物种丰富度指数(d)而言,虽然湖泊水体最高,其次为河流水体,水库水体最低,但是三者之间的差异并不显著(两两之间的Wilcoxon秩和检验p值均大于0.05)。PCOA(图3)和ANOSIM分析结果(R=0.048,p=0.033)也表明,3种水体类型之间的群落结构没有显著差异。此外,3种水体类型的优势种绝大部分是交叠的。鲫、饰妆铠弓鱼、草鱼、鲤、泥鳅、大鳞泥鳅、青鳉和麦穗鱼等8种鱼在3种水体类型均为优势种,而团头鲂MegalobramaamblycephalaYih是湖泊水体特有的优势种(Y=0.030),高体鳑鲏RhodeusocellatusKurumeus是河流水体特有的优势种(Y=0.031)(表1)。因此,北京地区的水库、湖泊和河流3种水体类型的鱼类多样性和群落结构无显著差异。

图3 北京地区不同采样点的鱼类多样性

图4 不同采样点鱼类群落组成的主坐标PCOA分析
2.2 本地鱼种和外来鱼种分类汇总
与张姗(2020)汇总的北京本地鱼类库(本地种83种,外来种13种)进行比较,本研究利用环境DNA分别检测到39种本地鱼种和13种外来鱼种。与北京地区历史记载的83种本地鱼种相比,本地鱼种明显下降。此外,本研究检测到的外来鱼种中入侵种的比例较高,占外来鱼种46.2%。说明近年来北京地区原生鱼类多样性下降明显,可能是由于大量外来入侵鱼类被引入北京地区水域,对北京本地鱼类多样性产生了严重影响。
本研究共检测到尼罗罗非鱼Oreochromisniloticus(L.)、大口黑鲈MicropterussalmoidesLacèpede、蟾胡鲶Clariasbatrachus(L.)、加蓬胡鲶ClariasgabonensisGünther、饰妆铠弓鱼和坦噶尼喀口孵非鲫Oreochromistanganicae6种外来入侵鱼种。饰妆铠弓鱼在所有样点中被检出;蟾胡鲶在所有水库和河流样点(26个)及6个湖泊样点中被检出;尼罗罗非鱼在26个样点中被检出,在河流水体中普遍存在;加蓬胡鲶在14个样点中被检出,水库和湖泊检出率较高;坦噶尼喀口孵非鲫和大口黑鲈分别在7和4个样点中被检出。
2.3 外来入侵鱼类风险评估
本研究参照FISK打分系统说明(Coppetal.,2016)及相关文献(Killietal.,2019;Interesovaetal.,2020),分别计算6种外来鱼类的BRA(basic risk assessment)指数和BRA+CCA(BRA plus climate change assessment)指数,分析外来鱼类入侵风险。根据北京水域特点和外来鱼类情况,本研究设置BRA指数阈值为18,即入侵低风险(得分<0)、中风险(1≤得分≤18)、高风险(得分≥19);设置BRA+CCA指数阈值设置为28,即入侵低风险(得分<0)、中风险(1≤得分≤28)、高风险(≥29)。2种指数计算结果见表2。根据BRA指数和BRA+CCA指数评价结果,本研究检测到的所有6种外来入侵鱼类在北京地区都具有高入侵风险。

表2 基于FISK V2指标体系的典型外来入侵鱼种的入侵风险评估
3 讨论
本研究利用环境DNA宏条形码技术调查分析了北京地区的鱼类多样性和群落组成现状,共检测到52种淡水鱼类(包括本地和外来鱼类),与张春光等(2011)、张姗(2020)调查结果差别不大。监测结果中,鲤形目鲤科的鲫鱼丰度最高且出现在所有样点中,其次是同属鲤科的麦穗鱼,这一结果与北京地区历史鱼类调查结果也一致(李雪健等,2018;邢迎春等,2007;张春光等,2011;张姗,2020)。本研究结果表明,环境DNA技术检出了与传统鱼类多样性调查方法相当的鱼种,并检测到传统采样方法难以发现的稀有物种,例如在海子水库(S61)中检测出黄尾鲴XenocyprisdavidiBleeker和宽鳍鱲Zaccoplatypus(Temminck et Schlegel)2种稀有物种,这2种鱼在近5年的传统方法调查中均未被发现。说明环境DNA宏条形码方法不仅可以实现快速、高效鱼类监测,还具有较高的灵敏性。由于水体中环境DNA存在机制较为复杂,通过环境DNA序列丰度评估物种、生物量等方面还有待进一步研究(Evansetal.,2016;Pontetal.,2018)。因此,建议将环境DNA宏条形码方法广泛应用于水生态监测,建立环境DNA水生生物多样性监测技术标准,实现对鱼类、贝类、大型水生植物、两栖类等生物监测。
本研究发现,水库、湖泊和河流3种水体类型的鱼类多样性和群落结构无显著差异,不同水体类型中鱼类物种同质化程度显著,这可能是引入大量外来鱼类导致的(张姗,2020; Oldenetal.,2004)。通过分析比较各样点间鱼类种类、生物量及多样性指数,发现山区自然水体样点的物种多样性及生物量高于城区水体样点。例如,山区海子水库东湖(S61)和八渡(S64)样点鱼类种类最多、生物量较高,而城区圆明园福海(S38)鱼类物种数目最少、生物量最低。整体而言,八渡、海子水库等山区河流、水库样点的鱼类物种多样性高、生物量大。这些处于上游山区、受人类活动影响较小、近自然状态的山区河湖,水质条件好,适合鱼类生存。而圆明园福海等城区河湖可能受城市化进程和水体污染等影响(张姗,2020)。由于北京市主要河流城市段的生境类型比较单一,渠化现象较为普遍,底质多以淤泥为主,鱼类群落空间格局遭到破坏,鱼类群落退化,具有典型的城市受干扰河流鱼类特征。此外,城区与山区采样点的差异还可能与增殖放流有关。例如,城区河湖鱼类优势种主要为放流鱼种,例如鳙鱼,属于滤食性的鱼种,主要用于净化和改善水质。上述结果表明,人类活动和城市化进程对北京河流鱼类多样性和群落空间分布产生显著影响(李雪健等,2018;张姗,2020)。
本研究共检测到39种北京地区的本地原生鱼种,远远低于历史记载本地原生鱼种(累计83种)。近年来北京地区鱼类多样性调查实际检测到的本地鱼种分别为65种(1984年)、43种(2011年)和32种(张姗,2020),表明北京地区的本地鱼类多样性呈逐年下降趋势。据调查,在北京本地鱼类调查中有记录的原生鱼类,如乌苏里拟鲿PseudobagrusussuriensisDybowski和尖头鱥Rhynchocyprisoxycephalus(Sauvage and Dabry)等(李雪健等,2018;杨文波等,2008),本次调查中所有样点均未检测到。另一方面可能是由于这些本地原生鱼种的野生种群数量极少甚至已灭绝消失。除了城市化和环境污染等人类活动干扰的影响,采样点的布设也可能是导致环境DNA检出原生鱼类多样性较少的一个原因。本研究主要依据北京市水生态监测的站点进行布点采样,大部分采样点位于城市中心或者郊区平原的水体,而北京地区北部和西南部的山地采样点较少,这些人为干扰较少、较为原生自然的山区水体中往往具有较高的本地鱼类多样性(李雪健等,2018;张姗,2020),因此可能存在部分本地物种由于采样点覆盖不全而未检测到的情况。
与历史调查数据相比,本研究发现,北京地区外来鱼种的比例大大增加,而且一些外来鱼种的检出率和序列丰度超过了本地物种。说明近些年来大量外来鱼类被引入北京地区,这可能是导致北京本地鱼类多样性下降的主要原因之一。其中绝大部分外来种是因水产养殖业需要而引入北京的,有些已成为野外繁殖种群,包括团头鲂、大口黑鲈和细鳞裂腹鱼等。此外,饰妆铠弓鱼和尾纹魮Labeobarbuscaudovittatus(Boulenger)等主要作为观赏鱼养殖和放生而进入北京河湖。值得关注的是,本次调查监测到的外来鱼种中有6种入侵鱼种,且具有高入侵风险。其中,烈性入侵物种蟾胡鲶,在几乎所有的样点(32个)中均检测到,而且生物量巨大,此物种原产于东南亚,现已入侵到全球众多水域,可捕食养殖鱼及蝌蚪等,并对本地鱼类有较大竞争优势(Baber & Babbitt,2003),已被世界自然保护联盟(IUCN)列为“世界百大外来入侵种”;原产于非洲的入侵鱼种尼罗罗非鱼,也在大部分样点(26个)中检出,此物种可吞食其他鱼类的受精卵,严重影响物种多样性;原产于北美的入侵鱼种大口黑鲈,于1983年第一次被引入中国(Maetal.,2003),现已成为我国重要的淡水养殖鱼类(李家乐,2007;张姗,2020),此物种会捕食较小的原生鱼类,造成当地鱼种数量降低甚至灭绝。
综上所述,北京地区的本地鱼类多样性呈逐年下降趋势,外来鱼种的比例大大增加,且存在多种高入侵风险的鱼类,相关监管部门应建立环境DNA宏条形码鱼类监测技术平台,开展全市五大水系鱼类多样性普查性监测,针对重点河流、湖泊、水库和水源地的外来鱼种和入侵鱼种开展定期监测、科学预警和防治,通过连续性监测掌握其种群动态和危害变化,降低外来鱼类对北京水生态系统鱼类多样性的危害。
猜你喜欢
湖北植保(2022年4期)2022-08-23
土壤(2021年1期)2021-03-23
科学养鱼(2020年8期)2020-12-20
流行色(2020年9期)2020-07-16
中国农村水利水电(2020年4期)2020-06-12
当代水产(2019年8期)2019-10-12
活力(2019年21期)2019-04-01
活力(2019年21期)2019-04-01
北京广播电视报(2017年35期)2018-02-28
科学种养(2017年1期)2017-02-16
