
诱变结合小孢子培养的氮高效大麦DH株系评价
2021-09-23 09:19高润红徐红卫郭桂梅宗营杰李颖波杜志钊陈志伟陆瑞菊王亦菲刘成洪
麦类作物学报 2021年5期
高润红,徐红卫,郭桂梅,宗营杰,李颖波,杜志钊,陈志伟,陆瑞菊,王亦菲,刘成洪
(1.上海市农业科学院生物技术研究所, 上海 201106; 2.上海市农业遗传育种重点实验室, 上海 201106;3.上海市农业科技服务中心,上海200335)
氮素是作物生长所必需的元素之一,近几十年农业生产力的提高与氮肥使用量大幅增加有关[1]。由于谷类作物的氮效率比较低,只有33%的肥料氮在谷物中得到利用[2]。过量的氮肥施用会导致资源的巨大浪费以及潜在的主要污染物[3],因此,提高作物氮效率是作物改良的主要 目标。
大麦作为世界上第四大谷类作物,主要用于动物饲料和啤酒酿造。到目前为止,大麦育种主要集中在提高籽粒产量和品质上,与此同时氮效率也得到了提高,而且氮效率的两个组成部分氮素吸收效率和氮素利用效率在基因型间也存在显著差异[4]。但是由于集约化的育种和栽培导致大麦的遗传多样性变窄,成为遗传改良的瓶颈[5],因此,创制和筛选氮高效突变体的种质资源成为氮高效育种的一种有效的方法。
单倍体和加倍单倍体是加速植物育种的一种特别有吸引力的生物技术方法。单倍体技术通过配子胚胎发生,使杂合子亲本一步培育出完整的纯合品系,已经对许多农作物产生了重要的影响[6]。加倍单倍体在单倍体诱导后的第一个世代就能达到100%的纯合率,不仅加速育种进程,而且从单倍体中获得目的基因型(包括突变体)所需要的群体规模较小,同时也可以用于发现隐性、显性以及有害性突变等[7]。单倍体诱导系统为突变体的诱导和选择提供了一个理想的靶位点[6]。小孢子培养技术是产生单倍体的主要手段,通过诱变并与小孢子培养相结合可以在第一代中筛选隐性突变体,避免嵌合体;通过加倍能够获得纯合稳定的突变体,缩短纯合突变体产生的周期[8],而且在一个培养体系里可同时实现突变体的诱变、筛选和纯合稳定[9]。诱变结合小孢子低氮胁迫培养获得低氮高效突变体材料已经在前期的工作中得到验证[10],获得的低氮高效突变体材料氮效率的提高均是由于氮素吸收效率提高所导致[11-12],还未筛选到氮素利用效率提高的突变体材料。
本实验室利用诱变并结合小孢子低氮胁迫培养创制了一批DH株系,经过连续多年的田间肥力水平鉴定,获得表现优异的突变体材料。本研究以花30不同诱变来源的8份DH株系为研究对象,在两个氮肥水平下,以产量为标准将其划分为不同的氮效率类型,同时对双高效类型和低氮高效类型的株系进行干物质和氮素积累、转运特征以及氮效率相关指标分析,以期为大麦氮高效筛选评价机制提供依据,并为氮高效品种的选育奠定材料基础。
1 材料与方法
1.1 供试材料
供试材料为大麦(HordeumvulgareL.)品种花30以及花30不同组织(小孢子、种子和离体穗)经过化学或者物理诱变并结合小孢子低氮胁迫培养获得的8份DH株系,分别为A1-184、A1-226、A9-41、A9-47、A4-17、A4-21、A5-24和A11-15。其中A1-184和A1-226是离体小孢子经过3 mg·L-1的甲基磺酸乙酯(ethyl methyl sulfone, EMS)诱变后低氮胁迫培养获得;A9-47和A9-41是离体小孢子经过5 mg·L-1的EMS诱变后低氮胁迫培养获得;A4-17和A4-21是干种子经过400 GY的钴60(60Co)辐照后田间种植,取游离小孢子低氮胁迫培养获得;A5-24是干种子经过500 GY的60Co辐照后田间种植,取游离小孢子低氮胁迫培养获得;A11-15是离体穗经过5 GY的60Co辐照后,取游离小孢子低氮胁迫培养获得。材料创制方法详情见陆瑞菊等[13]的 方法。
1.2 田间试验设计
试验于2018-2019年在上海市农业科学院重固试验基地进行。大田试验采取裂区设计,氮肥水平为主因素,株系为副因素,3次重复,小区行长3.3 m,行距0.33 m,每个材料种5行,每行180粒种子。基本苗为150万株·hm-2。施肥水平为45 kg·hm-2纯氮(低氮)和160 kg·hm-2纯氮(高氮)。低氮只施入基肥,高氮施入基肥和追肥,基肥与追肥分配比例是3∶7,以氮磷钾复合肥(各占15%)作为基肥在播种之前均匀施入,尿素(含N 46%)作为追肥,在分蘖期和拔节期分1∶1施入。其他措施按照一般田间管理方式进行。
1.3 样品采集与测定
于开花期与成熟期,每个小区随机选取5株生长表现一致的单株,分穗子和营养器官收获,105 ℃杀青30 min,75 ℃烘干至恒重,称干重并记录。粉碎后精确称取0.100 0 g,浓硫酸消煮后定容至100 mL,取5 mL利用连续流动分析仪进行氮素含量测定。
1.4 计算方法
干物质积累量、氮素积累量、花前干物质转运量(花后干物质积累量)对籽粒产量的贡献率和花前氮素转运量(花后氮素积累量)对籽粒氮素含量的贡献率的计算参考熊淑萍等[14]的方法;氮效率、氮素吸收效率以及氮素利用效率的计算参考Moll等[15]的方法。
1.5 数据分析
数据用Excel进行整理,并利用DPS 7.05软件进行显著性分析(LSD法)。
2 结果与分析
2.1 田间氮高效株系的筛选
为了筛选氮高效株系,以产量性状为指标,分别在高氮和低氮水平下进行比较分析。从表1可以看出,以亲本花30作为对照,在高氮水平下有2份DH株系(A9-47和A1-184)的产量显著高于花30;低氮水平下有3份DH株系(A5-24、A4-17和A9-47)的产量显著高于花30。在田间施氮量降低70%的情况下,产量也明显下降;与高氮水平相比,低氮水平下大多数株系包括亲本花30的产量均下降了60%以上,其中,A9-41和A1-226的降幅达到了70%以上,说明这两份株系对氮肥比较敏感;在这些株系中只有A5-24和A4-17的产量下降的幅度在60%以下,说明这两份材料的耐低氮性要比其他株系好。

表1 花30和不同DH株系在两个氮肥水平下的产量表现Table 1 Different yield performance of Hua 30 and the DH lines under two nitrogen fertilizer levels

Ⅰ:双高效型;Ⅱ:低氮高效型;Ⅲ:双低效型;Ⅳ:高氮高效型。H30:花30;HN:高氮;LN:低氮。
为了明确不同DH株系对氮肥的响应,以高氮和低氮水平下的平均产量为基准,将包括花30在内的9份株系分为4个不同的区。从图1可以看出,A9-47和A1-184为双高效类型,这2个DH株系在高氮和低氮水平下产量都高于供试株系的平均产量;A4-17和A5-24为低氮高效类型,这2个DH株系在低氮水平下产量高于供试株系的平均产量;A11-15、A4-21和花30归为双低效类型,这3个株系在高氮和低氮水平下产量均低于供试株系的平均产量;A1-226和A9-41归于高氮高效类型,这2个株系只有在高氮水平下产量才高于供试株系的平均产量。
2.2 不同氮肥水平下双高效和低氮高效类型株系与亲本花30开花期和成熟期的干物质和氮素积累以及转运特征
基于环境友好型考虑,分析了双高效和低氮高效类型中4份材料的干物质和氮素积累特征(表2)。高氮水平下,开花期和成熟期A9-47和A1-184的干物质积累量以及A9-47的氮素积累量均显著高于亲本花30。低氮水平下,开花期A9-47和A1-184的干物质积累量和氮素积累量均显著高于亲本花30,成熟期四份株系的干物质积累量均显著高于亲本花30,其中A5-24和A9-47的氮素积累量也显著高于亲本花30。说明氮高效株系具有较高的干物质和氮素积累能力。
对花前干物质转运量(花后干物质积累量)对籽粒产量的贡献以及花前氮素转运量(花后氮素积累量)对籽粒氮素含量的贡献进行分析。结果(表3)表明,无论高氮还是低氮水平,花后干物质积累量对籽粒产量占主要贡献,而高氮水平下花前氮素转运量对籽粒氮素的含量占主要贡献,低氮水平下花后氮素积累量占主要贡献。对于不同株系而言,高氮水平下,A5-24和A9-47的花前干物质转运量对籽粒产量的贡献率显著高于花30,而花30的花后干物质积累量对籽粒产量的贡献率最高,显著高于A5-24和A9-47,与其他株系之间差异不显著;A5-24和A1-184的花前氮素转运量对籽粒氮素的贡献率显著高于A4-17,和亲本花30和A9-47之间差异不显著,而A4-17的花后氮素积累量对籽粒氮素的贡献率显著高于A5-24,与亲本花30、A9-47和A1-184之间差异不显著。低氮水平下,A1-184的花前干物质转运量对籽粒产量的贡献率最大,其次是花30和A9-47,三者之间差异不显著,A1-184的花前干物质转运量对籽粒产量的贡献率显著高于A5-24和A4-17,而A4-17的花后干物质积累量对籽粒产量的贡献率显著高于A1-184,与其他株系之间差异不显著;A1-184的花前氮素转运量对籽粒氮素的贡献率最大,其次是花30,两者之间差异不显著,而A1-184的花前氮素转运量对籽粒氮素的贡献率显著高于A9-47、A5-24和A4-17,且花30的花前氮素转运量对籽粒氮素的贡献率显著高于A4-17,而A4-17的花后氮素积累量对籽粒氮素的贡献率显著高于花30和A1-184,且A5-24、A4-17和A9-47的花后氮素积累量对籽粒氮素的贡献率均显著高于A1-184。说明同一类型株系之间的花前干物质(氮素)转运量和花后干物质(氮素)积累量对籽粒产量(氮素)的贡献也可能存在显著的差异,推测同一类型株系之间也可能存在不同的氮素吸收利用机制。
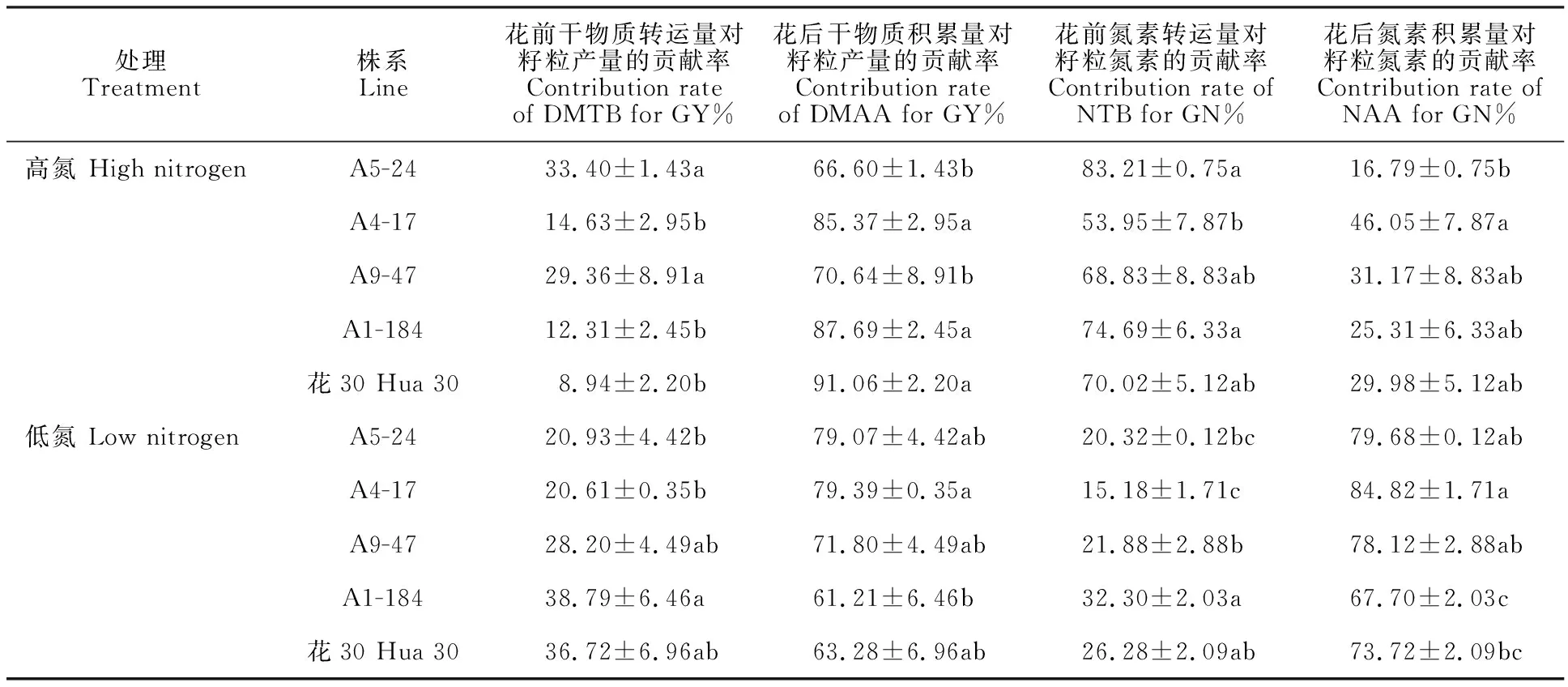
表3 花30与不同DH株系干物质和氮素转运特征Table 3 Characteristics of dry matter and nitrogen transportations between Hua 30 and DH lines
2.3 不同氮肥水平下双高效和低氮高效类型株系的氮素吸收利用效率分析
从表4可以看出,高氮水平下,A9-47和A1-184的氮效率与亲本花30相比,分别提高了 19.54%和19.68%,均显著高于亲本花30;A9-47的氮素吸收效率显著高于亲本花30,是花30的1.5倍;A1-184的氮素利用效率显著高于亲本花30,增幅14.53%。低氮水平下,A5-24和A9-47的氮效率和氮素吸收效率均显著高于亲本花30,增幅在20%以上,而A4-17的氮素利用效率略高于亲本花30,但差异不显著。说明在高氮水平下,A9-47氮效率的提高主要是由于氮素吸收效率的提高所导致,而A1-184氮效率的提高主要是由于氮素利用效率的提高所导致;在低氮水平下,A9-47和A5-24氮效率的提高主要是由于氮素吸收效率的提高所导致,A4-17具有相对较高的氮素利用效率,但氮效率与亲本花30差异不显著。值得注意的是,低氮水平能够提高氮效率和氮素利用效率,表明低氮处理下较高的氮素同化能力能够促使氮效率提高并促进产量增加。
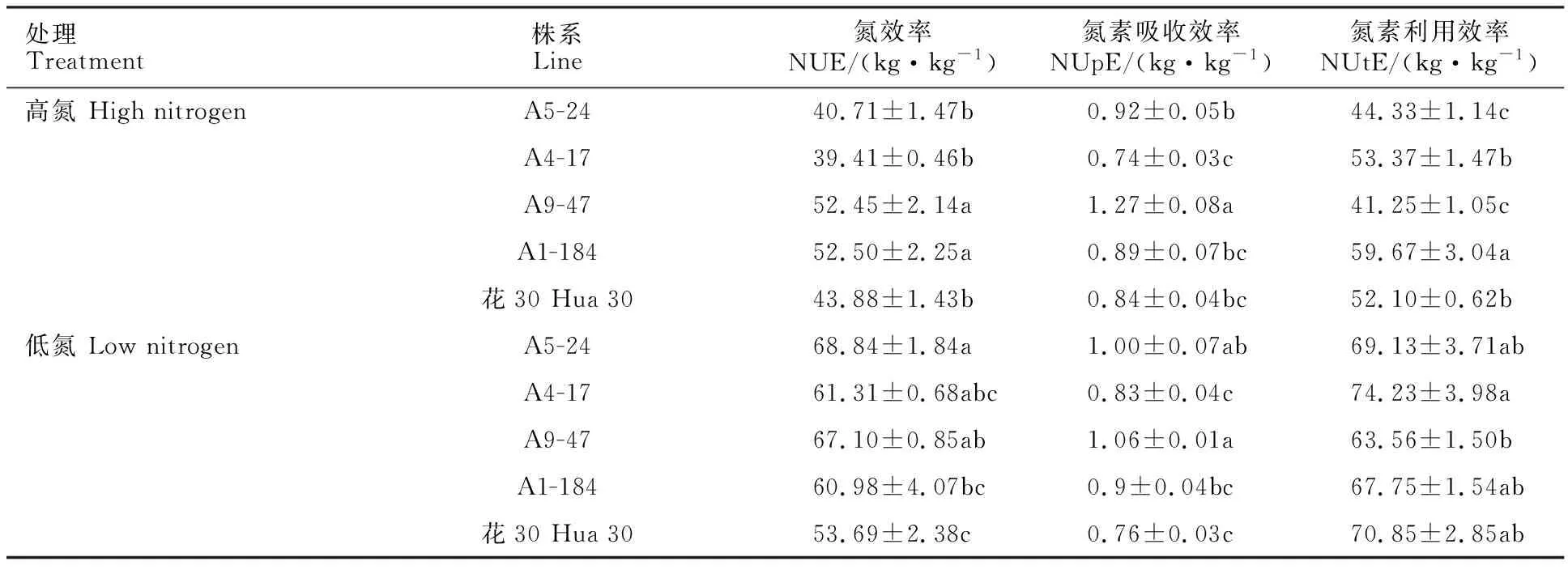
表4 花30与不同DH株系在两个氮肥水平下的氮素吸收利用情况Table 4 Difference of nitrogen use efficiency, nitrogen uptake efficiency and nitrogen utilization efficiency among Hua 30 and DH lines under two nitrogen fertilizer levels
3 讨 论
3.1 关于氮效率类型的划分
氮素在农业和食物系统中的行为与资源、环境、食物安全和人体健康等密切相关,是反映农业绿色发展的关键指标[16],氮高效种质资源的创制和新品种培育是发展绿色农业的一个重要途径。氮效率被定义为单位面积土壤供氮量所产生的作物籽粒产量,在同一个供氮水平下,多以籽粒产量作为氮效率评价的标准[17-18]。本研究在两个氮肥水平下根据籽粒平均产量将花30及8份DH株系划分为双高效、低氮高效、高氮高效和双低效四种类型。花30作为亲本被划分为双低效型,A1-184和A9-47被划分为双高效型,其在高氮和低氮下均能够获得较高的籽粒产量,这两个株系在实际生产中具有较大的应用潜力。A5-24和A4-17被划分为低氮高效型,这两个株系在低氮胁迫下能更好的吸收和利用氮素,可能存在氮高效吸收利用的分子机制。
目前,世界各地大多数品种测试系统都使用相对较高的氮素供应,育种者选择的都是在这种水平下表现较好的品种,但是由于基因型和供氮量的互作,高氮水平下选择的高产品种在低氮水平下表现并不一定优异[19],因此,为了可靠地确定哪些品种在低氮水平下产量较高,需要在低氮水平下进行育种和测试[20]。Gouis等[21]证实了低氮水平下籽粒产量存在遗传变异,基因型和氮素互作显著;徐 晴等[17]认为,低氮处理更容易显示出不同种质在氮素利用相关性状上的差异;黄 亿等[22]认为,氮高效基因型大麦在各个生育期的物质生产和氮素积累能力强,而且低氮处理下优势较为明显;Brancourt-Hulmel等[20]已经证明,小麦在低氮水平下进行直接选择会更有效。本研究中A5-24和A4-17在低氮水平下的优异表现也证明了这一点,说明进行低氮胁迫筛选有必要且有用。
3.2 关于开花前后干物质和氮素积累量对籽粒产量的影响
植物在营养生长期,根系和叶片起到储存器官的作用,吸收无机氮用于氨基酸和蛋白质的合成;在转运期,植物根系和叶片将营养器官中储存的氮素转移到生殖器官[23-24]。有研究表明,大多数氮素在开花期之前积累在营养器官中,而在花后吸收的氮素很少,籽粒中的氮素积累主要依赖于营养器官的氮素供应[25-26]。本研究中,在氮素充足的条件下,同一个株系开花期和成熟期的氮素积累量之间差异较小,说明籽粒中积累的氮素在开花前就已经积累在营养器官中,而且花前氮素转运量对籽粒氮素的贡献率大于花后氮素积累量,也证明了这一观点。而在低氮胁迫条件下,花前干物质转运量对籽粒产量的贡献率和花前氮素转运量对籽粒氮素贡献率都比较小,可能是由于缺氮导致光合作用受阻,抑制营养体的生长,使籽粒干重和氮素含量主要来源于花后干物质和氮素的积累。
作物干物质生产和氮素积累特性与产量和氮效率关系密切[22]。徐 晴等[17]认为,氮素高效小麦应具有较高的形成生物学产量和积累氮素的能力;李淑文等[27]表明,氮胁迫条件下小麦生育中后期较强的氮素吸收能力是氮高效品种在缺氮条件下籽粒产量和氮效率提高的重要生理基础;Worku等[28]研究表明,低氮条件下热带玉米杂交种的高产与后期较高的氮素吸收量、单位氮素积累量和氮素收获指数密切相关。叶利庭等[29]认为,齐穗后氮高效水稻具有较强的氮素吸收或转运能力才能满足植株对氮素的利用。在本研究中,对于双高效型株系A9-47和A1-184而言,不管是高氮水平还是低氮水平,开花期和成熟期的干物质和氮素积累量均高于或者显著高于亲本花30;低氮水平下,A5-24和A4-17在成熟期的干物质和氮素积累量均高于或者显著高于亲本花30,这种较高的干物质和氮素积累能力为籽粒产量的提高奠定生理基础,同时也可以作为氮高效种质材料筛选的基础指标。
3.3 关于氮素高效吸收(NUpE)与氮素高效利用(NUtE)材料的获得
氮效率可以被分为氮素吸收效率和氮素利用效率两部分,这两个部分哪个更重要尚无统一报道。在玉米中有报道认为,在高氮水平下,氮效率的变化可以用氮素吸收能力的变化来解释,而在低氮水平下,氮效率的变化主要是由于氮素利用效率的不同导致的[15,30];刘 鹏等[31]认为,高粱在高氮处理水平下,氮素吸收效率和氮素利用效率对氮效率贡献相同,而在低氮处理水平下,氮素利用效率对氮效率贡献大于氮素吸收效率;张亚丽等[18]认为,吸氮量和氮素利用效率对产量的贡献率受到水稻生育期的影响,对于生育期较短的水稻,无论供氮水平高低,均是氮素利用效率对产量起决定作用;而生育期较长的水稻,低氮水平下是氮生理利用效率起决定作用,高氮水平下是氮吸收量起决定作用;Worku等[28]研究表明,低氮水平下玉米的氮素吸收效率和氮素利用效率对氮效率有同等贡献;在前期工作中我们筛选到了氮高效大麦种质材料,不管是高氮还是低氮水平,氮效率的提高均是由氮素吸收效率提高导致的[11-12],而在本研究中不仅筛选到了氮素吸收效率提高的DH株系A9-47和A5-24,而且也筛选到了氮素利用效率提高的DH株系A1-184和A4-17。高氮水平下,A1-184的氮素吸收效率和亲本花30没有显著差异,但是氮素利用效率的显著提高导致了氮效率的提高;而低氮水平下,A4-17的氮素利用效率也略高于亲本花30,从而促使产量的 提高。
猜你喜欢
灌溉排水学报(2022年9期)2022-09-28
农业工程学报(2022年8期)2022-08-08
种子(2022年6期)2022-08-03
作物学报(2022年10期)2022-07-21
西南农业学报(2022年5期)2022-06-06
热带亚热带植物学报(2022年2期)2022-04-14
广东农业科学(2021年3期)2021-04-23
上海师范大学学报·自然科学版(2020年5期)2020-12-18
江苏农业科学(2020年14期)2020-08-28
三农资讯半月报(2020年13期)2020-07-31
