
耕地和林地土壤动物群落组成和多样性的差异
——中国区域的整合分析
2021-10-09 01:39郑大川李思雨
生态学报 2021年17期
郑大川,周 杰,李思雨,陶 军,*,顾 卫
1 北京师范大学环境演变与自然灾害教育部重点实验室, 北京 100875 2 北京师范大学地表过程与资源生态国家重点实验室, 北京 100875 3 北京师范大学地理科学学部, 北京 100875
全球变化是当今社会最突出的问题之一,而土地利用方式变化是全球变化的主要表现形式之一。20世纪以来,为了满足粮食生产需要,人类活动对土地的干扰日益加剧,在此长期过程中,已有大量的林地转变为耕地。林地转变为耕地造成了生境的破碎与植被的单一,对陆地生态系统生物多样性产生较大影响[1]。众多研究表明,耕地相比于林地不仅显著降低了地上生物多样性[1-2],也对地下生物多样性产生显著的影响[3-5]。地下生物多样性对地上生态系统变化的响应及其反馈作用已经成为陆地生态系统功能和服务以及可持续性研究的前沿领域和关键突破口。土壤动物作为生态系统分解者的重要组成部分,在凋落物的分解、土壤结构的改善、养分的释放和植物的生长等生态系统功能和服务上扮演极为重要的角色[6-7]。因此,研究不同土地利用方式下地下土壤动物密度、种群和多样性的响应差异,对于陆地生态系统多样性的稳定与可持续性具有重大意义。
目前,在我国关于耕地和林地两种土地利用方式下土壤动物多样性特征及其影响因素已开展了许多的研究。其中有些研究表明,耕地显著减低土壤动物的个体密度、类群数和多样性[8-10],但也有研究发现耕地土壤动物数量、类群数和多样性与林地无差异,甚至高于林地。影响土壤动物在耕地和林地间响应差异的因素,一方面可能归因于土壤环境因素[7,11],如土壤含水量、土壤有机碳和全氮等[5,12-13];另一方面也可能受到区域气候条件(温度和降水)的影响。有研究表明土壤动物数量和多样性受到年降雨量变化的影响[14-15];耕地和林地下土壤生物群落和多样性的变化主要归因于土壤水分和温度的变化。另外,中国区域不同类型的耕地和林地间土壤动物群落也存在不同响应差异。关于耕地土壤动物的研究包括水田和旱田两类[16];林地土壤动物的研究包括自然林、人工林和经济林[17-20]。因此,上述耕地和林地的利用类型如何影响耕地和林地间土壤动物数量和多样性的响应差异也需要进一步的研究。综上,对于耕地和林地方式下土壤动物数量和多样性的响应差异研究主要集中于小尺度的研究点上,而对于较大尺度(中国区域)的研究较为欠缺[21],这可能主要受到土壤动物采样和鉴定工作量的限制,而无法开展区域大尺度研究。而整合分析(Meta-Analysis)为该方面的解决找到另一条可能的途径[22]。因此,本研究基于当前研究现状,提出以下假设:(1)气候条件(温度和降水)会影响耕地与林地之间的土壤动物数量、组成和多样性的响应差异;(2)林地和耕地类型也会影响耕地与林地之间的土壤数量、组成和多样性的响应差异。
本研究采用Meta分析对中国地区耕地与林地间土壤动物群落差异进行定量的综述性研究,并通过分析气候条件、耕地和林地类型等因素,揭示中国区域耕地和林地间土壤动物数量、组成和多样性的响应特征。该研究在保护土壤动物多样性、维持地下生态系统稳定性方面,为人为改变土地利用方式的决策提供理论依据。
1 材料与方法
1.1 数据收集

图1 研究区地理分布Fig.1 Geographical distribution of the study areas
在Web of Science与中国知网文献数据库分别以“soil fauna”、“Arthropods”和“land-use”,与“土壤动物”、“节肢动物”和“土地利用”作为关键词进行检索获得文献。再根据以下标准对检索到的文献进行筛选:(1)研究对象为土壤动物群落,评价指标为个体数、个体密度、类群数或Shannon-Wiener多样性指数;(2)每个研究至少包含耕地和林地两种土地利用方式的对照试验;(3)每个处理至少3个重复;(4)文献中需明确告知研究区地理位置和采样时间;(5)分层取样的试验中需分别给出不同土层中各指标的数值;(6)分次取样的试验中需分别给出不同时间采样的各指标的数值。
经过筛选最终纳入中英文文献共37篇,包括188项独立试验,研究区分布如图1所示。选取土壤动物的个体密度、类群数和Shannon-Wiener多样性指数三个指标定量阐述中国土壤动物群落对耕地和林地响应的差异。另外还需提取土壤动物分类、年均温、年降水量、土壤动物尺寸、耕地类型和林地类型等信息,以便进行亚组分析。
1.2 研究方法
本研究在进行文献数据搜集时,如果数据以正文或表格形式给出,可直接读取;若文献中的数据是用图的形式表示,则用 GetData Graph Digitizer 2.24软件来提取。由于各个研究采样面积与采样次数不同,因此对数据进行标准化处理,以单位面积中平均个体数和类群数进行分析。 若文献中提供的数据为标准误,则统一转换为标准差,标准差可通过公式(1)得到:
(1)
式中,n是重复次数。统计学指标采用响应比(response ratios,RR)表示,并计算其 95%的置信区间(95% CI)。其计算公式为:
(2)

(3)
整合分析通过对每个独立研究的响应比进行加权,得出加权平均响应(RR++)。若土壤动物个体密度、类群数和Shannon-Wiener多样性指数的RR++的 95%的置信区间未跨过横坐标零点,则说明耕地相比林地差异显著(增加或降低)。反之则说明耕地土壤动物相比于林地差异不显著。土壤动物个体密度、类群数和Shannon-Wiener多样性指数变化的百分数(耕地土壤动物相比于林地土壤动物减少的百分数)可以通过(eRR++-1)×100%公式计算得到[23]。对耕地和林地土壤动物物种组成(21目)分别进行整合分析,以各个目的个体密度为指标,得加权平均响应,找出受耕地和林地差异影响的主要物种。亚组分析中以年均温、年降水量、耕地类型、林地类型和土壤动物尺寸等条件进行分组,进一步分析影响耕地和林地土壤动物差异的因素。
1.3 数据分析
采用 Meta Win 2.1 软件进行 Meta 分析[24]。Meta 分析合并独立研究的响应比得出加权平均响应比前,需明确试验各处理之间及各试验结果是否存在异质性(处理间或不同研究结果间的变异是否由随机误差引起)。因此,采用卡方检验(Chi-square test)进行异质性检验,如检验结果P>0.05,说明不同处理间或不同研究结果间具有同质性,可选用固定相应模型计算合并统计量,否则采用随机效应模型[25]。本研究经异质性检验表明存在明显异质性,故采用随机效应模型(P<0.001)。由于研究中水田的数据均来自湿润区(年降水量>800mm),故基于耕地类型的亚组分析只选取湿润区的研究,以消除气候条件造成的土壤水分差异[26]。
根据Meta分析结果,采用GraphPad Prism 8 软件进行森林图的绘制。
2 结果
2.1 土壤动物群落对耕地和林地响应的总体差异
Meta分析结果(图2)表明,与林地相比,中国的耕地显著降低了土壤动物的个体密度(55.74%)、类群数和(27.56%)Shannon-Wiener多样性指数(24.51%)。个体密度的结果与类群数和Shannon-Wiener 多样性指数的结果均存在显著差异。

图2 耕地和林地土壤动物差异Meta分析森林图Fig.2 Forest map of meta-analysis of differences in soil fauna between arable land and woodland小数为平均效应值;百分数为耕地土壤动物相比于林地土壤动物减少的百分比;括号内的数字为独立研究数量/文献数量
2.2 土壤动物个体密度在不同条件下的响应差异
年均温高于20℃时,林地与耕地间土壤动物的个体密度差异不显著;年均温低于20℃时,耕地土壤动物个体密度显著低于林地。年均温高于20℃的耕地与林地间土壤动物个体密度的响应比显著高于年均温低于20℃的响应比。不同年降雨量下,耕地土壤动物个体密度显著低于林地。在不同耕地类型、林地类型和土壤动物尺寸的亚组中,耕地的土壤动物个体密度显著低于林地土壤。耕地与自然林间土壤动物个体密度的响应差异(-79.96%)显著大于耕地与人工林(-45.46%)或经济林(-36.73%)的响应差异(图3)。
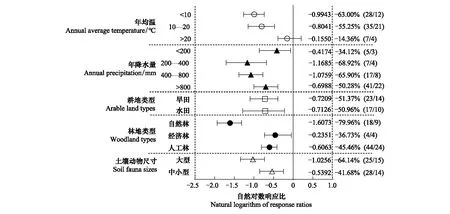
图3 土壤动物个体密度亚组分析森林图Fig.3 Forest map of individual density subgroup analysis of soil fauna
2.3 土壤动物类群数在不同条件下的响应差异
当年均温高于20℃时,耕地与林地间的土壤动物类群数差异不显著(图4)。年降水量少于200mm的地区耕地与林地间土壤动物类群数差异不显著;年降雨量高于200mm的地区耕地的土壤动物类群数显著低于林地土壤。在年降水量400—800mm地区,耕地与林地间土壤动物类群数的响应比最大,且与降水量<400mm地区的响应值差异显著。不同耕地类型、林地类型和土壤动物尺寸的亚组中,耕地的土壤动物类群数均显著低于林地土壤。土壤动物类群数在耕地与自然林间的响应比(-42.08%)大于耕地与经济林(-20.95%)和人工林(-20.93%)。耕地与林地间大型土壤动物类群数的响应比(-39.56%)显著大于小型土壤动物类群数的响应比(-17.93%)。
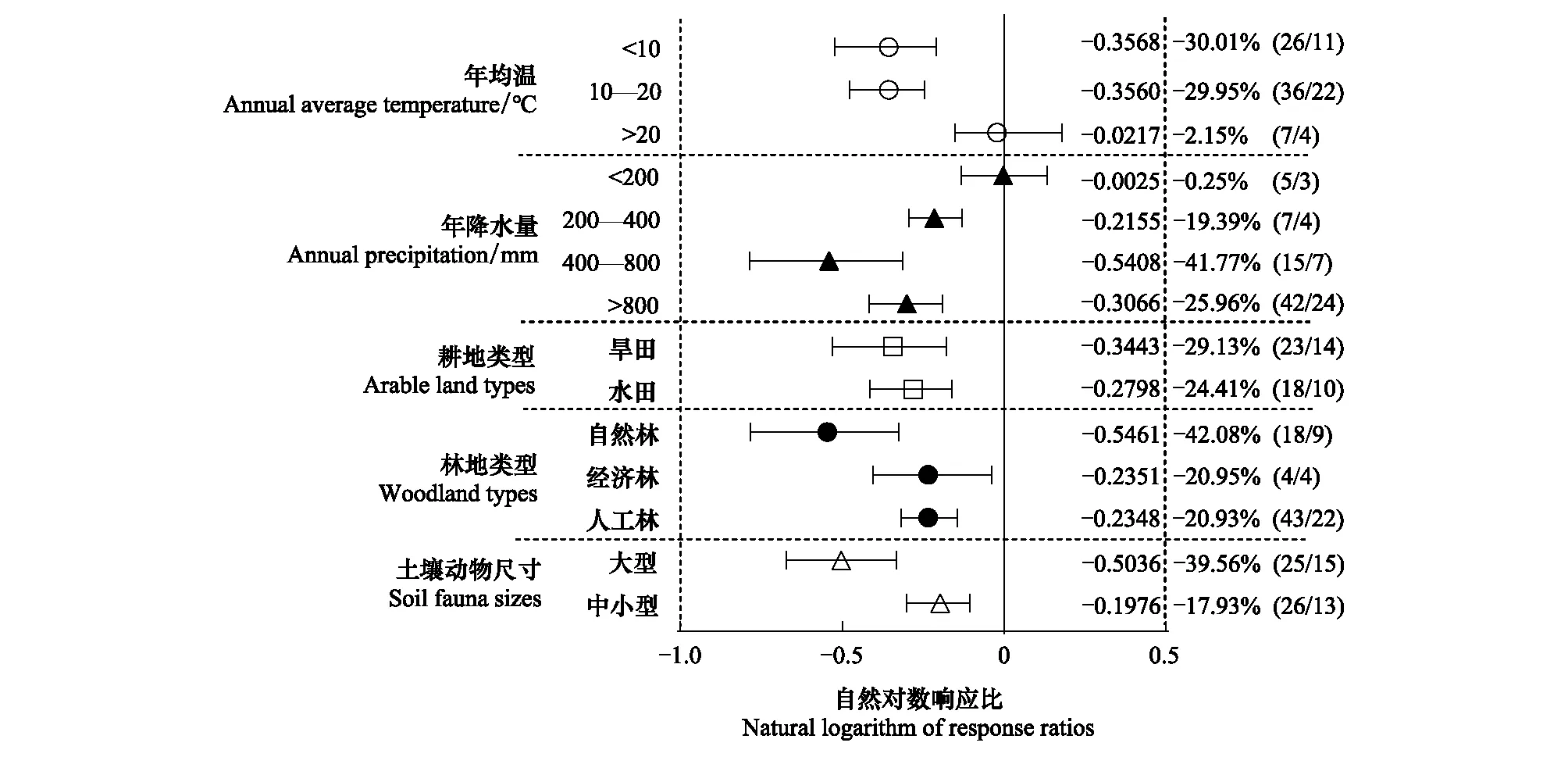
图4 土壤动物类群数亚组分析森林图Fig.4 Forest map of subgroup analysis of soil faunal groups
2.4 土壤动物Shannon-Wiener 多样性指数在不同条件下的响应差异
年均温低于20℃条件下耕地土壤动物Shannon-Wiener 多样性显著低于林地;而年均温高于20℃条件下耕地土壤动物Shannon-Wiener 多样性与林地无差异;年降水量200—400mm条件下耕地土壤动物Shannon-Wiener 多样性与林地无差异;年降水量400—800mm条件下耕地土壤动物Shannon-Wiener 多样性的响应比最大。旱田土壤动物Shannon-Wiener 多样性显著低于林地;而水田的土壤动物Shannon-Wiener 多样性与林地无显著差异。耕地土壤动物Shannon-Wiener 多样性显著低于自然林和人工林土壤,而耕地与经济林间无差异。耕地与自然林间土壤动物Shannon-Wiener 多样性的响应比(-48.20%)高于人工林(-17.40%)。耕地土壤大型和小型动物Shannon-Wiener 多样性均显著低于林地(图5)。

图5 土壤动物Shannon-Wiener指数亚组分析森林图Fig.5 Forest map of soil faunal Shannon-Wiener index subgroup analysis
2.5 土壤动物组成的响应差异
分别对耕地和林地21个主要物种组成的个体密度进行Meta分析(图6)。结果表明土壤动物不同种类在耕地和林地中表现出不同的差异。12个类群在耕地中显著低于林地,其中以膜翅目(Hymenoptera)降低最为明显(-75.79%);弹尾目(Collembola)、双翅目幼虫(Dipteralarvae)、正蚓目(Lumbricina)、颤蚓目(Tubificida)、等翅目(Blattaria)、同翅目(Homoptera)、鳞翅目幼虫(Lepidopteralarvae)和直翅目(Orthoptera)8个类群差异不显著;而革翅目(Dermaptera)在耕地的个体密度显著高于林地(102.97%)。

图6 土壤动物主要种个体密度森林图Fig.6 Forest map of individual density of main groups of soil fauna
根据耕地和林地的不同类型,分别对4个优势类群的个体密度进行亚组分析(图7)。结果表明,蜱螨目(Acarina)和膜翅目的个体密度效应值在不同土地类型中均表现出显著性的差异,分别在自然林亚组(-88.10%)和水田亚组(-85.65%)差异最大;鞘翅目(Coleoptera)在自然林(-56.77%)和旱田(-65.62%)的亚组中差异显著;弹尾目仅在自然林亚组(-75.68%)中存在显著差异。
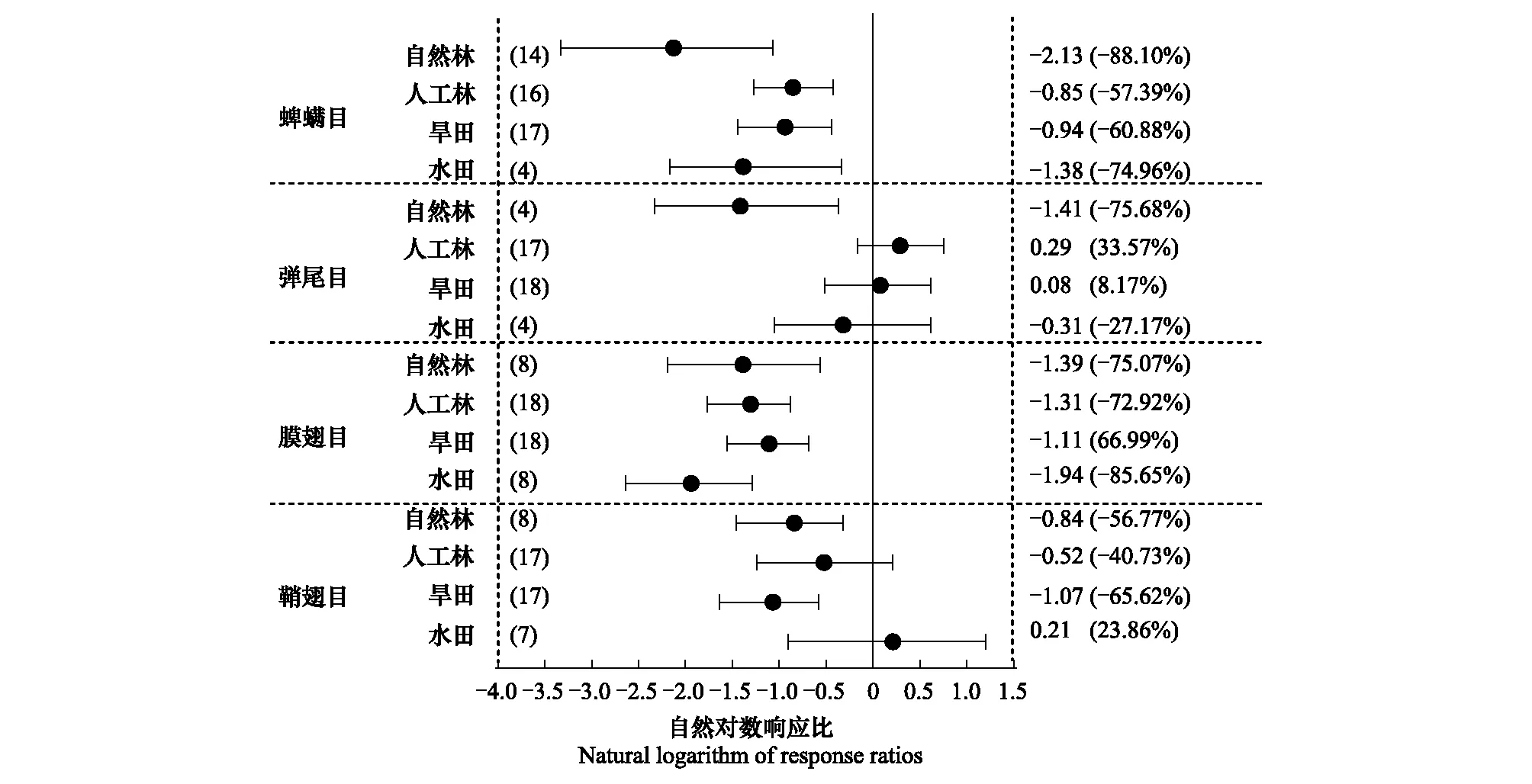
图7 土壤动物优势种个体密度亚组分析森林图Fig.7 Forest map of individual density subgroup analysis of dominant groups of soil fauna
3 讨论
3.1 温度和降水的影响
温度是调节陆地生态系统生物地球化学过程的重要环境因子,由于土壤环境相对稳定,土壤动物对温度变化的敏感性明显高于地上生物[27],而不同土地利用类型下土壤温度对气温的响应也存在差异。年均温高于20oC地区耕地与林地间土壤动物数量、类群数和多样性无差异,且它们间的响应绝对值显著小于年均温低于20oC地区。这一结果可能归因于林地土壤动物数量和多样性的降低,进而缩小了耕地与林地间差异。在温度较高的地区,林地表层凋落物快速分解[28],凋落物层的减少增大林地土壤温度变化幅度[29],进而可能对土壤动物产生不利影响;另一方面,林地快速凋落物分解不利于土壤有机质和营养物质的累积[30-31],降低土壤动物食物资源,影响了土壤动物群落。
土壤水分是影响土壤动物群落组成关键环境因子。由于多数土壤动物生活于土壤孔隙中,其活动依赖于水分的可获得性[32-33],降水可以对土壤动物产生直接影响[34],从而显著影响土壤动物群落组成和多样性带来显著影响;降水量在400—800mm地区耕地与林地间土壤动物数量、类群数和多样性的响应绝对值高于降水量大于800mm和降水量小于400mm地区。在降水量小于400mm地区,耕地与林地间土壤动物多样性没有差异。前人研究表明干旱会限制土壤微生物生物量[7,35],可能限制了土壤大型动物数量、类群数和多样性[26]。林地的大型土壤动物减少则使耕地和林地差异减小。在降水稀少的地区,土壤动物群落的主要限制因素是土壤水分而非土地利用方式,故土壤动物无法对耕地与林地的差异进行有效的响应。降水量大于800mm地区水和热(温度较高)加快林地凋落物分解,土壤有机质累积少,导致这一地区小型土壤动物数量和多样性较低[36],可能是导致耕地与林地土壤动物的差异减小的原因之一。另一方面,有研究表明较高的降水量使得土壤有较高的无机氮和较低的pH值,虽然地上生物多样性较高,但土壤生物多样性相对较低[37-38],进而可能缩小耕地和林地间的差异。此外,降水量大于800mm地区耕地多为水田,其土壤动物数量和多样性与林地差异较小(图5),也影响了土壤动物群落在耕地和林地间响应差异。年降水量400—800mm地区的研究主要集中在我国的东北地区,该地区的水分和温度(较低)导致林地凋落物分解慢[39],土壤有机质和营养物质丰富,其土壤生物多样性最高[26],使得耕地与林地间土壤动物群落的差异显著。
3.2 耕地与林地类型、动物尺寸的影响
耕地与林地类型不同程度的改变土壤理化性质,进而影响土壤动物数量、组成和多样性[6]。耕地中旱田土壤动物Shannon-Wiener多样性显著低于林地,但耕地中水田土壤动物Shannon-Wiener多样性与林地间差异并不显著。有研究表明水田相较于旱田受到人为干扰较小,生境得到保护,有利于土壤动物多样性增加[14],进而可能使得耕地中水田与林地间土壤动物多样性差异性减少。林地类型也不同程度地影响了土壤动物对耕地和林地的响应。耕地与自然林间土壤动物数量、类群数和多样性的响应绝对值最大,其次是耕地与人工林间,耕地与经济林间的响应绝对值最小(差异不显著)。研究表明人工林和经济林受人为干扰强度较大,如自然林<人工林<经济林。这与前人研究结论一致[40-41]。人为干扰对于经济林的作用与农田生态系统类似,强于人工林,长期和定期的耕作、施肥和除草等土地管理方式都会直接抑制土壤动物数量、组成和多样性[42-43]。此外,上述管理措施也可能通过改变土壤理化性质间接影响土壤多样性[44]。
另外,研究结果表明大型土壤动物数量对于耕地和林地的响应绝对值高于中小型土壤动物,这一结果说明大型土壤动物对土地利用方式的变化更为敏感。这一结果可能归因于大型土壤动物比中小型土壤动物更容易受到农业管理措施的影响[45],如翻耕、杀虫剂使用、机械压实等措施[46-47]。林地转变为耕地的过程中,土壤动物群落也向中小型土壤动物为主的方向转变,该结论与前人研究成果相符[45,48]。
3.3 耕地和林地土壤动物物种组成的差异
蜱螨类和弹尾类通常是我国各类土地利用方式下土壤生态系统中优势种[4]。亚组分析结果表明,耕地蜱螨类数量显著低于林地。这可能因为土壤螨类对耕地干扰活动(如翻耕)较敏感,进而导致其数量显著降低[49-50]。亚组分析结果也表明,耕地土壤中弹尾目数量与林地差异不显著。这是因为林地中人工林土壤中弹尾目的数量低,且耕地中旱田弹尾目数量较高,进而导致它们在耕地与林地间无差异。在长江中下游地区研究表明,弹尾类属于喜干类土壤动物,使其成为旱地的优势类群,且数量较多[51]。但在辽河平原地区,研究表明旱地土壤弹尾类数量较低[14]。这一矛盾结果可能归因于两个地区土壤类型和理化性状的差异。林地正蚓类数量低于耕地,这可能由于林地中蚯蚓一般属于深层类,其个体较大,占据更多生态位,但数量较少;而耕地土壤中蚯蚓都属于内生型蚯蚓,其个体小,但数量较多,所以导致耕地蚯蚓数量高于林地。某些物种对生活环境有较强的选择性,如优势类群中以蚁科(Formicidae)为主的膜翅目,农田的耕作活动影响蚁类筑巢而导致其数量明显减少[52-54]。农田耕作还会减少鞘翅目,特别是捕食性步甲科(Carabidae)的个体数量,并可以将其用来指示土壤环境的变化[55-57]。常见类群中的蜘蛛目(Araneae)、双翅目(Diptera)等也主要生活在植被较完整的林地、灌木环境中,耕地中少见。稀有类群中的等足目(Isopoda)和倍足目(Diplopoda)等,对干扰尤为敏感,也表现出了在耕地与林地数量上的显著差异,在某些特定生境条件适宜时,可能会成为某生境的常见类群,故可用于指示土地利用的变化[58-59]。
4 结论
基于整合分析表明,中国区域的耕地与林地间土壤动物数量、组成和多样性的响应差异也受到区域气候条件(温度和降水)的影响;耕地和林地类型也影响耕地与林地间土壤动物数量、组成和多样性的响应差异。耕地使得蜱螨类(优势种)的数量显著低于林地,而弹尾类和正蚓类的数量在耕地与林地间无差异。今后的研究可能需要将区域气候条件与土壤环境变化结合起来,阐明不同土地利用方式间土壤动物群落结构的响应特征,揭示区域地下生物群落组成和多样性的影响机制。
猜你喜欢
农业科技与信息(2020年14期)2020-12-18
中国中医基础医学杂志(2020年1期)2020-03-03
心电与循环(2020年1期)2020-02-27
绿色科技(2019年14期)2019-11-19
中国医学影像技术(2019年10期)2019-10-24
江苏农业科学(2019年5期)2019-09-02
陕西农业科学(2019年6期)2019-07-19
黑龙江粮食(2019年4期)2019-05-10
农民致富之友(2018年2期)2018-03-25
农民致富之友(2018年1期)2018-02-26
