
模拟增温对吕梁山生态脆弱区披针苔草光合特性的影响
2021-10-12 08:11于秀立赵红倩张慧敏葛浩彬张晋龙王永珍徐满厚
山西农业科学 2021年10期
于秀立,赵红倩,张慧敏,葛浩彬,张晋龙,王永珍,徐满厚
(太原师范学院地理科学学院,山西晋中030619)
气候变暖是全球气候变化的主要趋势之一,联合国政府间气候变化专门委员会(IPCC)2018年10月份最新报告指出,目前全球平均气温已经比工业革命前上升1℃左右[1];到21世纪末全球气温将升高0.3~4.8℃[2],未来几十年全球气温变暖的速度可能会加快[3],气温增加将更为突出[4]。温度控制着陆地生态系统中植物的生物化学反应速率,尤其是光合作用,进而影响其他生物学过程[5]。光合作用决定着植物营养物质的积累,是植物生长、发育和繁殖的基础[6];极易受到自然界环境变化的影响,尤其是温度的变化,并对不同生境表现不同的适应性[7]。前人研究结果发现,植物光合作用对温度升高既有表现出增强[8-14]、也有表现出减弱[13,15-18]或无影响[12,19-21]的情况,这些研究结果的差异可能与各自的研究方法、所选取的植物种类和生态型对温度的敏感性以及植物光合作用的最适温度不同有关。
吕梁山地处黄土高原,其自然环境恶劣,植被稀少,沟壑纵横,水土流失严重,是典型的生态环境恶劣地区[22];其森林覆盖率仅为18.5%,也是重要的生态功能区。披针苔草(Carex lanceolata)为吕梁山林下草本群落的优势种,是一种重要的牧草资源,对维护该区域的生态稳定具有重要的作用。针对该区域植被的研究主要集中在物种多样性[23-25]、群落多样性[22,26-27]及其与纬度、海拔和环境因子的关系[26-30]以及生物量[31]等方面;也有一些研究利用模拟增温探究物种多样性的变化等[24,30,32]。目前对干旱胁迫条件下毛苔草和不同水分亏缺灌溉条件下青绿苔草的光合特性已有报道[33-34],对毛苔草、青绿苔草、木里苔草和宽叶苔草在自然条件下光合特性也有研究[35-38],还有研究自然寒冻条件下崂峪苔草、青绿苔草以及青绿苔草变种光合速率的变化[39]。但是在该区域内模拟增温对植物光合特征的影响研究还比较少,尤其是有关披针苔草在模拟增温下光合特性的研究还未见报道。
本研究利用开顶式增温箱(OTC)进行模拟增温,以吕梁山林下草本群落优势种披针苔草为研究对象,研究气候变暖对植被的影响,揭示披针苔草光合特性对模拟增温的短期响应及其适应气候变化的生理机制,旨在为未来气候变化背景下该区域植被的生理机制研究提供科学参考,也为该区域植被恢复与重建提供理论依据。
1 材料和方法
1.1 研究区概况
吕梁山位于我国的第二大阶梯上,是山西省西部的一条总体走向为东北—西南的山脉,其南北走向长约400 km,东西走向宽80~120 km,该山脉主要包括北麓的管涔山、中麓的关帝山和南麓的五鹿山,总体上呈现北高南低的趋势,海拔地带性特征明显;纬度跨度较大,纬度地带性显著。该区域具有独特的气候与复杂的地形条件,给动植物资源提供了良好的生存环境,使该区域物种多样性丰富。本研究以吕梁山脉中段的关帝山、北段的管涔山为研究区。管涔山(地理坐标为东经112.60°~112.62°、北纬38.95°~39.05°)位于山西省忻州市宁武县东寨镇,研究区域的海拔在2 222~2 571 m;该区域的年平均气温为6.5℃左右,年平均降水量为475 mm左右,植被的垂直地带性分布比较明显[27]。关帝山(地理坐标为东经110.30°~111.30°、北纬37.33°~38.33°)位于山西省吕梁市交城县庞泉沟镇,研究地海拔为2 000~2 179 m,年平均气温3~4℃,年平均降水量830.8 mm,植被的垂直地带性分布差异显著[27]。
1.2 样地设置
试验在关帝山设置2个海拔梯度、管涔山设置3个海拔梯度,每个海拔梯度设计1个样地,共计5个样地,每个样地除增温处理外其他环境条件基本一致。其中,在关帝山选取的样地平均海拔分别为2 000、2 179 m;在管涔山选取的样地平均海拔分别为2 222、2 395、2 571 m。在各样地内选取植被分布均匀、地势较平坦、人为干扰少的林下草本层布设样地,样地大小为10 m×10 m,该样方内草本植物以披针苔草为主,并在周边用铁丝网加固,以保护样地。模拟增温试验采用开顶式增温箱增温,增温箱均为正六边形,顶面边长均为25 cm,各斜边于地面的夹角均为60°。在每个样地内随机布设低度增温(LW)样方、高度增温(HW)样方和对照(CK)样方各3个,每个海拔梯度共计9个样方,间距为1~3 m。其中,CK样方为植被自然生长,不进行增温处理,样方面积为1 m×1 m;LW样方的增温箱高度为40 cm,底面边长为50 cm,温度升高(1.7±0.3)℃;HW样方的增温箱高度为80 cm,底面边长为75 cm,温度升高(3.5±0.5)℃。增温装置于2016年8月全部布设完成。
1.3 光合作用指标的测定
于2019年7月下旬至8月初在各研究区进行野外试验测定,每个样方均随机选取生长状况一致的3组苔草进行编号处理,于13:00—15:00采用Yaxin-1102G便携式光合蒸腾仪测定苔草的光合参数,测定时采用开路测量的方法,设定人工光源,光强度为1 000 μmol/(m2·s)[36],叶面积设置为3 cm2。测定净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和细胞间二氧化碳浓度(Ci)等光合指标,并计算叶片水分利用效率(WUE)[40]和气孔限制值(Ls)[41],由于苔草较细长,一片叶子难以充满叶室,因此,用宽胶带将多个叶片尾部黏成一片,使叶室中的部分面积基本等于3 cm2,然后再利用仪器进行测量。

其中,Ca为大气中CO2浓度。
1.4 数据处理
利用Excel 2010对所有数据进行整理,使用SPSS 25.0软件对数据进行多因素方差分析、双变量相关性分析,相关图表采用Origin 2018完成。
2 结果与分析
2.1 不同海拔梯度下披针苔草光合特性的对比分析
海拔梯度的变化在一定程度上改变了植物的生长环境,进而影响着植物的光合特性[5]。本研究区内披针苔草的光合特性指标与海拔呈现出了一定的相关关系(表1),并且受海拔梯度变化的影响,披针苔草的净光合速率呈现了极显著差异(表2)。管涔山的海拔与披针苔草的光合特性相关性从大到小排序为:气孔导度>胞间二氧化碳浓度>水分利用效率>净光合速率>气孔限制值>蒸腾速率,气孔导度、叶片水分利用效率与海拔呈极显著正相关,胞间二氧化碳浓度与海拔呈极显著负相关,而净光合速率与海拔呈显著正相关,此外,蒸腾速率和气孔限制值与海拔呈正相关,但相关性未达到显著水平。关帝山的净光合速率、蒸腾速率、气孔导度、胞间二氧化碳浓度与海拔呈正相关,叶片水分利用效率和气孔限制值与海拔呈负相关,但均未达到显著水平(表1)。

表1 不同样地的海拔与披针苔草光合参数的相关性分析

表2 不同海拔和增温幅度及二者交互作用对披针苔草光合特性的影响
由图1可知,披针苔草的净光合速率、蒸腾速率和气孔导度随海拔的升高都呈现出双峰趋势,气孔限制值和水分利用效率随海拔升高呈现先减小后增加的单峰趋势,而胞间二氧化碳浓度随海拔的升高呈现先增加后降低的单峰趋势。在各光合指标中,净光合速率和蒸腾速率的峰值均出现在海拔最高的2 571 m处,叶片水分利用效率出现在海拔最低的2 000 m处。

2.2 模拟增温对披针苔草光合特性的影响
植物的光合生理特性对温度升高的响应不同,表现出了一定程度上的复杂性和不确定性[7]。披针苔草的光合特性变化幅度因海拔或者增温幅度的不同而存在着一定差异(表2)。不同海拔梯度对披针苔草的净光合速率、蒸腾速率、叶片水分利用效率产生了极显著影响(P<0.01),对气孔导度和胞间二氧化碳浓度产生了显著影响(P<0.05);不同的增温处理对披针苔草净光合速率的影响极显著(P<0.01),对其他参数影响不显著;海拔和增温的交互作用对披针苔草的净光合速率产生了极显著影响(P<0.01),对叶片水分利用效率影响显著(P<0.05),对其他光合参数的影响未达到显著水平。
2.2.1 净光合速率、蒸腾速率、胞间二氧化碳浓度与气孔导度的变化特征 从图2可以看出,净光合速率在HW-CK下呈明显的双峰曲线,中海拔处净光合速率波动较大且下降比较明显,最高峰值出现在2 179 m,相对增幅为0.12%,同时谷值出现在2 571 m,与对照相比降低了48.19%。净光合速率在LW-CK下呈单峰曲线,随海拔的增高呈现出明显的先上升后下降的趋势,峰值出现在2 222 m,相对增幅为17.11%,且谷值出现在2 000 m,相对降幅为80.11%。整体上,在高度增温处理下净光合速率随海拔的增高呈现下降趋势;在低度增温下净光合速率随海拔的增高呈现先增加后减小的趋势。
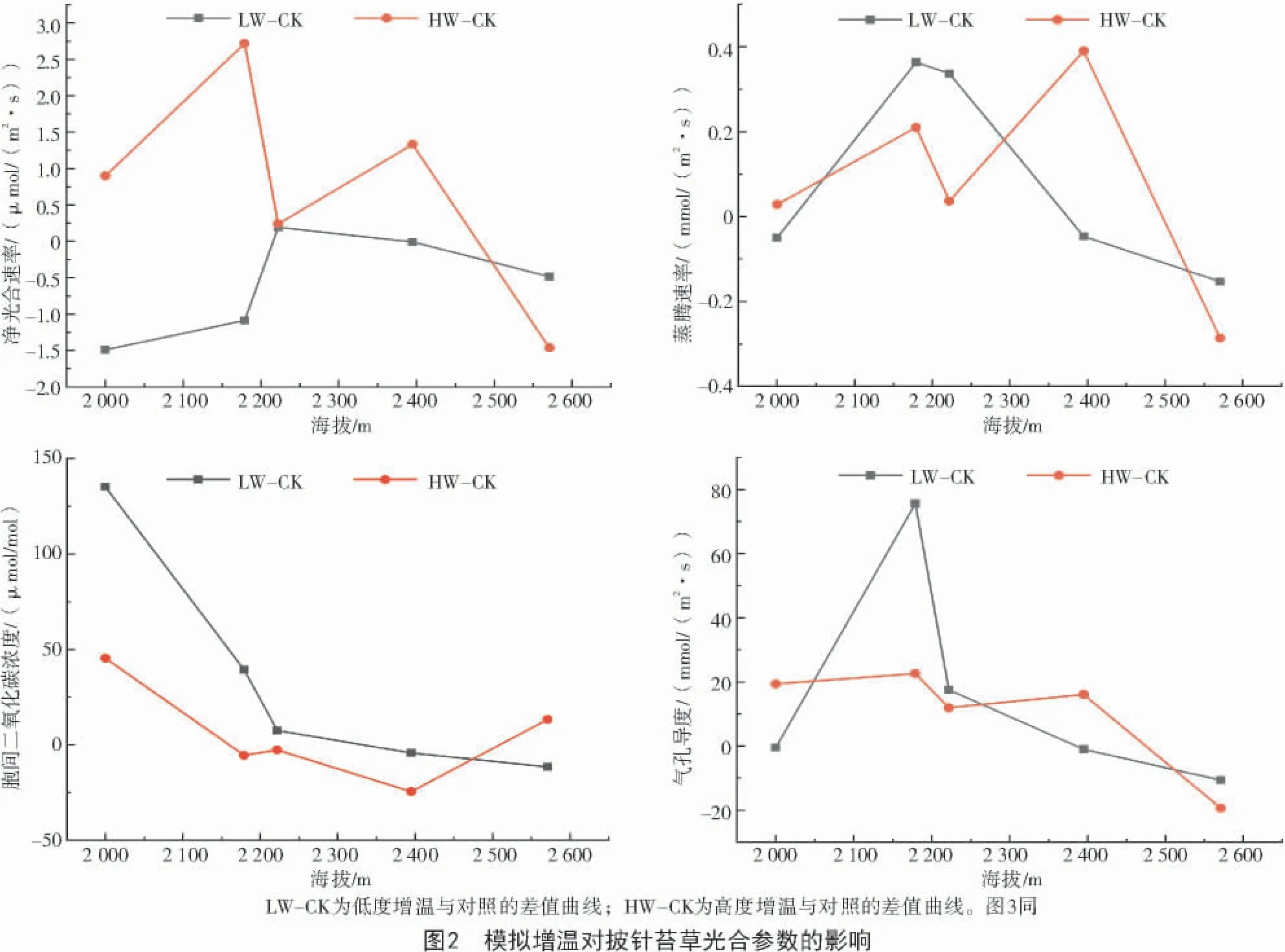
蒸腾速率在HW-CK下呈双峰曲线,最高峰值在2 395 m,相对增幅为99.15%,同时最低点出现在2 571 m,相对降幅为44.56%;在高度增温条件下,中低海拔处蒸腾速率呈现先增加后减小再增加的趋势,而在高海拔(2 571 m)处又呈现下降的趋势。蒸腾速率在LW-CK下呈单峰曲线,在低度增温条件下随海拔的增加呈现出明显的先上升后下降的趋势,且峰值出现在2 179 m,增幅为63.01%,谷值出现在2 571 m,且降幅为23.83%。整体上,蒸腾速率随海拔的增高呈现下降趋势。
胞间二氧化碳浓度的HW-CK曲线大致与LW-CK曲线走向一致,均随海拔升高呈现下降趋势。在高度增温处理下,披针苔草的胞间二氧化碳浓度HW-CK曲线的峰值出现在低海拔(2 000 m),增加了8.59%;谷值出现在2 395 m,降低了4.21%;低度增温处理下,胞间二氧化碳浓度LW-CK曲线的峰值位于低海拔(2 000 m),增加了25.56%,谷值在2 571 m,降低了2.24%。
气孔导度在HW-CK下呈逐渐下降的趋势;高度增温处理下气孔导度的HW-CK曲线峰值出现在低海拔(2 179 m)处,增加了36.71%,谷值在海拔2 571 m处,降低了43.17%。气孔导度的LW-CK呈现单峰曲线;低度增温处理下,随海拔的增高气孔导度呈现出先增加后降低的趋势;峰值出现在低海拔(2 179 m)处,增加了22.87%;谷值出现在高海拔(2 571 m)处,降低了23.76%。
2.2.2 水分利用效率与气孔限制值的变化特征 从图3可以看出,披针苔草的气孔限制值的HW-CK曲线表现为双峰,随海拔的增加呈现先增加后下降的趋势,峰值出现在2 395 m处,增加了54.87%,谷值出现在2 000 m处,降低了27.55%。气孔限制值在LW-CK下呈现单峰,随海拔的增加呈现上升趋势,峰值出现在2 571 m处,增加了9.88%,谷值于2 000 m降低了81.71%。整体上,气孔限制值随海拔的增高呈现增加趋势。

披针苔草的水分利用效率HW-CK曲线呈现单峰,随海拔升高而降低,峰值出现在2 000 m处,增加了61.56%,谷值出现在2 571 m处,降低了9.22%。在低度增温处理下,水分利用效率随海拔上升而呈现增加的趋势,峰值出现在2 571 m处,增加了9.21%,谷值出现在2 000 m处,降低了78.58%。水分利用效率HW-CK和LW-CK曲线随海拔升高而逐渐趋于0,说明增温对披针苔草水分利用效率的影响随海拔升高而逐渐减弱。
2.3 关帝山和管涔山披针苔草光合特性变化特征的对比分析
本研究发现,关帝山和管涔山在各自海拔梯度上增温后对披针苔草光合特性产生的影响也是有差异的。总的来说,关帝山的各光合参数的变化更为剧烈,管涔山则相对平缓。在未进行增温处理的情况下,管涔山披针苔草的光合特性与海拔的相关性比关帝山更显著(表1)。在低度增温处理下,关帝山披针苔草的净光合速率、蒸腾速率、气孔导度、水分利用效率、气孔限制值总体上呈现增加趋势,而胞间二氧化碳浓度总体上呈现降低的趋势;而在高度增温处理下,除胞间二氧化碳浓度和水分利用效率有所降低外,其余指标皆呈现增加趋势。在低度增温处理下,管涔山披针苔草的净光合速率、蒸腾速率、胞间二氧化碳浓度和气孔导度呈现下降趋势,气孔限制值和水分利用效率呈现增加趋势;在高度增温处理下,净光合速率、蒸腾速率、气孔导度和气孔限制值先增加后降低,胞间二氧化碳浓度先降低后增加,而水分利用效率有所降低(图2、3)。
3 讨论
3.1 海拔对披针苔草光合特性的影响
海拔是极其重要的地形因素之一,其对温度的影响最大。随着海拔升高,伴随着空气温度的下降、CO2分压降低以及光辐射强度增加等,进而对植物的生态和生理特征将带来巨大的影响[42]。温度影响植物的几乎所有生物学过程,特别是植物的光合作用,并且外界环境温度升高可能对其光合作用产生直接或者间接的显著影响[43]。徐亮[44]研究了不同海拔条件下春油菜光合生理特征,发现随海拔的升高春油菜的净光合速率呈现增加趋势。孙国超等[45]研究认为,随着海拔的上升黄果柑的净光合速率也增加。韦玉等[46]研究不同海拔高度矮嵩草的光合特性,结果表明,其净光合速率和光合作用能力随海拔梯度升高而逐渐增加。江厚龙等[47]研究发现,随海拔升高烤烟叶片的净光合速率呈现增加的趋势。就关帝山和管涔山而言,随海拔的升高,温度降低了,但披针苔草的光合速率和蒸腾速率也相应增加,可能是由于高海拔地区光辐射比较强所致,这与陈双林[48]的研究结论类似。然而,也有研究发现,随海拔的升高植物的光合作用能力降低。李惠梅等[49]研究了不同海拔麻花艽植物的光合特性,结果表明,高海拔地区的麻花艽植物受低温等条件的限制,其净光合速率低于低海拔地区。栗忠飞等[50]研究了不同海拔梯度下4种雨林幼苗的光合特性,结果表明,受低温的影响它们的净光合速率随海拔梯度的增加呈降低的趋势。就关帝山和管涔山整体海拔变化看,披针苔草的光合速率和蒸腾速率有所降低,可能是温度起主要作用引起的,温度降低进而影响光合作用相关酶的活性,这与罗旭[51]的研究结果类似。
3.2 模拟增温对披针苔草光合特性的影响
光合作用是植物生长发育和物质积累的基础[52],温度是影响植物光合作用能力强弱的重要影响因素之一[18]。叶旺敏等[14]研究了模拟增温对杉木幼树光合特性的影响,结果表明,增温条件下杉木幼树的净光合速率显著提高。本研究在高度增温时,披针苔草的净光合速率、蒸腾速率有所增加,可能是由于温度的增加使光合作用有关酶的活性提高所致,这与之前大部分研究结果一致[10,12-14]。闫玉龙等[53]研究了模拟增温条件下克氏针茅光合特性,结果表明,增温明显降低了其光合速率。引起植物光合速率降低的原因主要有2个:一是气孔因素;二是非气孔因素。究其原因,哪个因素起主要作用,主要是胞间二氧化碳浓度和气孔限制值的变化;胞间二氧化碳浓度降低而气孔限制值增加时,说明气孔因素占主导地位,反之则是非气孔因素占主导地位[54]。本研究在低度增温时,披针苔草的光合速率却降低了,究其原因可能是胞间二氧化碳浓度增加而气孔限制值降低的非气孔因素引起的;也可能是因为温度适当增加提高了披针苔草的光补偿点,呼吸作用变强,抵消了提高光合速率的效果[55]。
4 结论
本研究结果表明,关帝山和管涔山披针苔草的各项光合指标与海拔有一定的相关性;整体看,披针苔草的光合速率和蒸腾速率随海拔的升高呈现先增加后降低再增加的趋势。高度增温提高了披针苔草的净光合速率、蒸腾速率、气孔导度、胞间二氧化碳浓度和水分利用效率,降低了其气孔限制值;而低度增温降低了披针苔草的净光合速率、水分利用效率和气孔限制值,增加了其蒸腾速率、气孔导度和胞间二氧化碳浓度。因此,在高海拔地区,气候变暖有利于披针苔草的光合作用。本研究为该区域草本植物在气候变暖背景下的生理适应机制提供了参考依据。
猜你喜欢
灌溉排水学报(2022年9期)2022-09-28
农业工程学报(2022年8期)2022-08-08
作物学报(2022年10期)2022-07-21
农业工程学报(2022年4期)2022-04-24
农业与技术(2020年12期)2020-07-04
新课程·下旬(2017年7期)2017-08-14
物联网技术(2016年11期)2017-01-12
江苏农业科学(2015年8期)2015-09-10
天津农业科学(2015年8期)2015-08-06
湖北农业科学(2014年13期)2014-08-28
