
四川白水河自然保护区小型兽类物种多样性及群落结构
2021-10-14 01:59刘莹洵曾彦王旭明甘君巫嘉伟邓承敏张冬郭白濛廖锐
四川林业科技 2021年5期
刘莹洵,曾彦,王旭明,甘君,巫嘉伟,邓承敏,张冬,郭白濛,廖锐*
1.四川省林业科学研究院,四川成都 610081;
2.四川大学生命科学学院,四川成都 610011;
3.彭州市规划和自然资源局,四川彭州 611930;
4. 邻水县万峰山国有林场,四川邻水 635303;
5.西南山地,四川成都 610031;
6.四川农业大学,四川成都 611130
四川白水河自然保护区属于龙门山系核心地带,拥有丰富的生物多样性,在该保护区内曾开展了植物生物多样性及区系调查[1-3]和大型菌类调查[4],发现了抑虱跳小蜂属一新种[5],并对大熊猫[6-7]、林麝[8]等大型动物及红腹锦鸡等鸟类[9-10]进行过相关研究,但对于小型兽类的调查却较为缺乏,只有2001年原四川林业厅对白水河兽类资源调查中有所涉及[11],但并未开展过对小型兽类,尤其是非飞行性小型兽类的专项调查。小型兽类在生态系统中具有重要的作用,是食物链的重要组成部分,具有分布广、种类多、迁移性较小及其分布与栖息环境关系密切等特点,因此成为重要的生物多样性指示类群和研究动物地理学规律的代表性类群[12],也是森林生态系统中生产者和次级消费者能量传递中的重要组成[13]。通过对白水河自然保护区小型兽类的调查,其可以为保护区生物多样性的研究提供丰富的数据和资料,并通过对保护区垂直区系的研究分析,揭示各区系成分、物种丰富度、物种多样性随海拔变化的规律,以及不同群落的区系成分和物种多样性,为保护区的建设和生物多样性保护提供科学依据。
1 研究区概况
白水河自然保护区位于龙门山褶皱带的中南段,地处彭州市北部,地理位置介于北纬31°10′—31°29′、东经103°41′—103°57′之间,包括龙门山镇的中北部和小鱼洞镇的西部;东起蓥华山,西至光光山,南起安子坪则家沟源,北达太子城、南天门。保护区南北长约19 km,东西最宽约24 km;最低海拔1 481 m(锅框岩),最高海拔4 814 m(太子城),相对高差达3 333m;保护区总面积30150 hm2,全为国有林地。保护区于1998年成立的省级保护区,2004年批准为国家级自然保护区,是以保护大熊猫、金丝猴等国家重点保护动物及其栖息地为主的自然保护区。全国第4次大熊猫调查结果显示,白水河自然保护区有7只大熊猫。
2 调查与研究方法
2.1 野外调查方法
本次调查布设了1条样带:从最低点的罐子沟口开始,直到九峰山金顶区域。在选择的样带内,按照海拔每上升400 m布设1个调查区域,在调查区域内,按照植被类型,每个类型设置调查样方1个,每个调查样方下夹200夹次,布设50个陷阱。如果海拔上升不显著,则每前进2 km,布设1个调查区域。
2.2 分类鉴定方法
形态鉴定主要参考《中国兽类野外手册》[14],同时参考其他专著,如《中国动物志兽纲第六卷下册 仓鼠科》[15]。对于有疑问的标本,采取分子测序,得到cyt b序列后通过手工校正,并翻译为蛋白序列,以排除假基因,然后在GenBank中进行序列对比,下载相近序列,利用Mega6.0进行序列比对,确保序列均能翻译成氨基酸,以排除假基因,再利用下载的相关序列作为构建系统发育树的外群,构建系统发育树,确定物种种类。
查阅了《四川白水河自然保护区综合考察报告》[11],对一些物种的分布进行补充。
2.3 群落多样性分析
群落多样性计算方法参照《基础生态学(第2版)》[16]。不同群落内的物种组成按照鉴定结果统计,按照生境和海拔的不同,进行丰富度指数,辛普森多样性指数(Simpson,diversity index),香农-威纳指数(Shannon-Weiner index)和均匀度指数的计算,公式如下:
丰富度指数(Richnessindex)

式中,S为物种数,N为群落中所有物种个体数;
辛普森多样性指数(Simpson,diversity index):
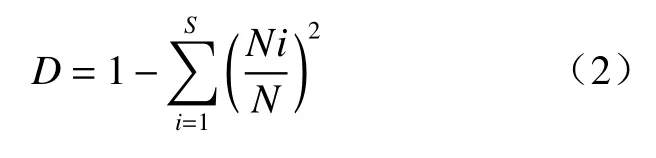
式中,S代表物种数,Ni表示种i的个体数,N表示群落中全部物种个体数;
香农-威纳指数(Shannon-Weiner index)计算群落多样性和均匀度指数:
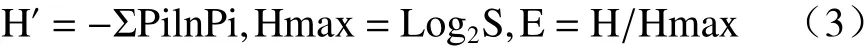
式中,S代表物种数,Pi属于种i的个体在全部个体中的比例。
3 结果
3.1 白水河自然保护区小型兽类多样性及区系分析
依据《中国动物地理》[17],白水河自然保护区有分布的44种小型兽类中,东洋界有33种,占总种数的75%,广布种2种,占4.55%,古北界9种,占20.5%,可见该区小型兽类以东洋界种类为主(见表1)。
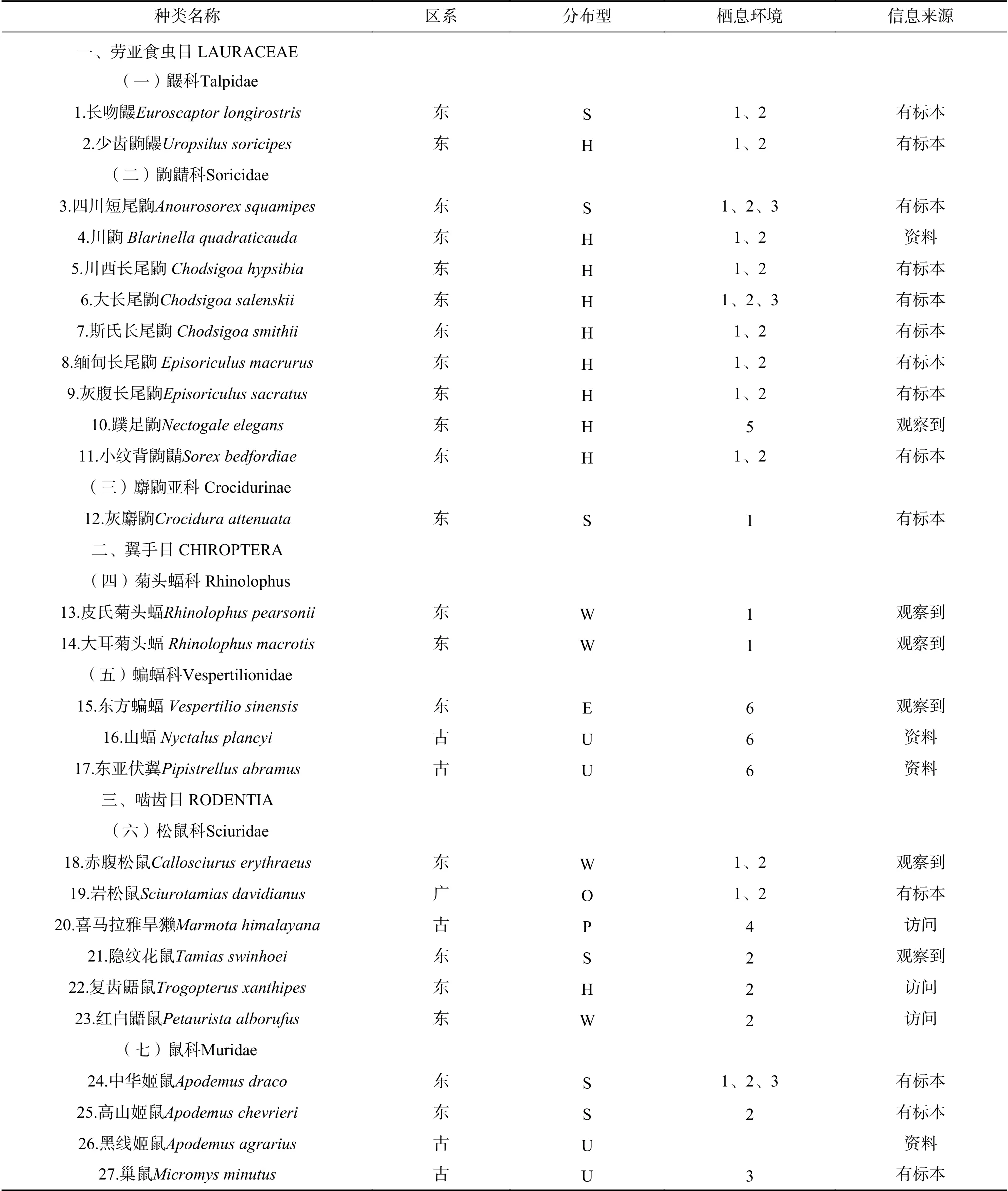
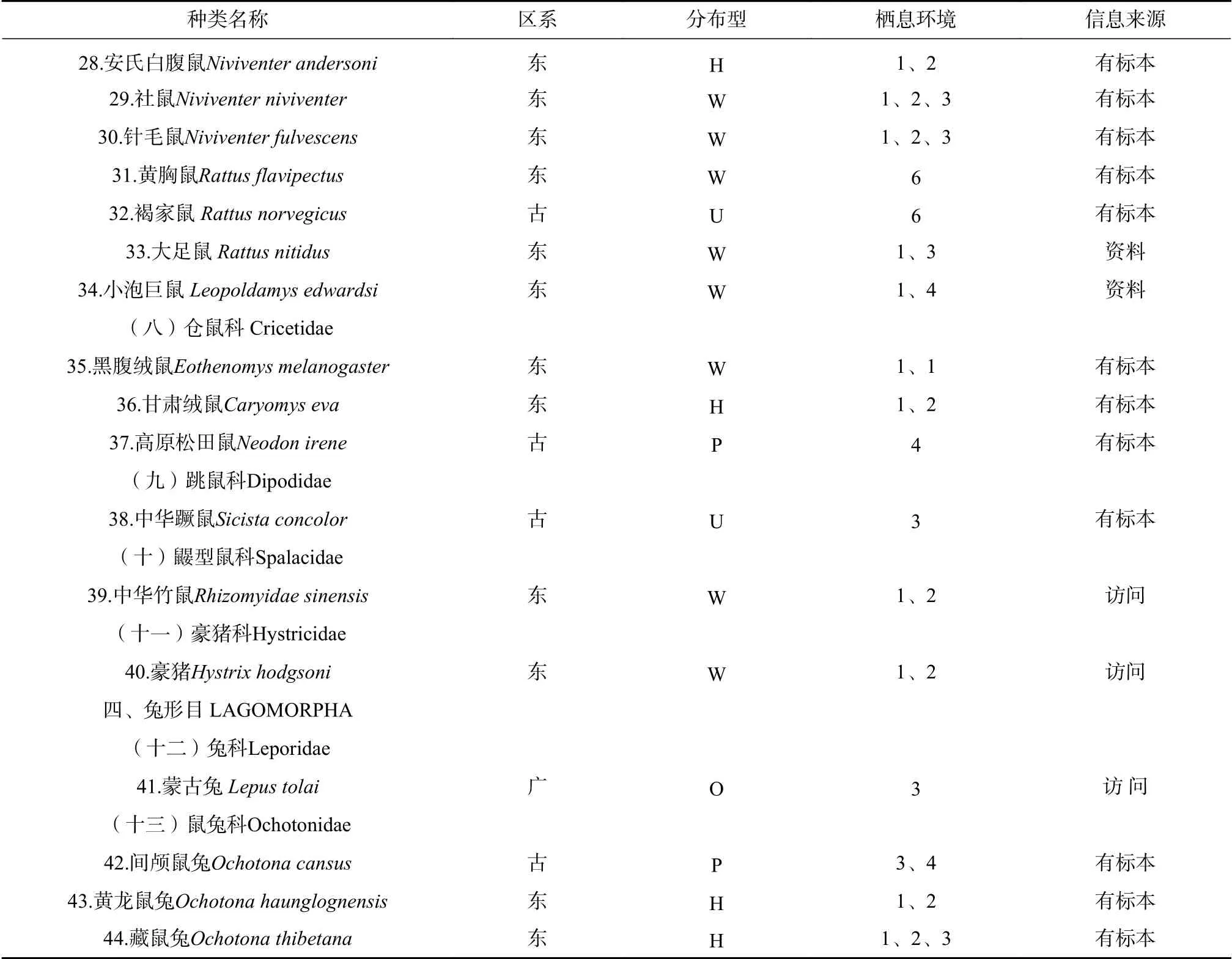
表1 白水河自然保护区小型兽类名录Tab.1 List of small mammal speciesin Baishuihe National Nature Reserve
从物种分布型来看,共有7种分布型。其中,东洋界有4种分布型,它们是喜马拉雅-横断山型、南中国型、东洋型和季风型。喜马拉雅-横断山型共计15种,它们大多是以横断山系为起源和演化中心的种类,包括了少齿鼩鼹(Uropsilus soricipes)、川鼩(Blarinella quadraticauda)、大长尾鼩(Chodsigoa salenskii)等;南中国型6种,包括长吻鼹(Euroscaptor longirostris)、灰麝鼩(Crocidura attenuata)、四川短尾鼩(Anourosorex squamipes)等;东洋型4种,黄胸鼠(Rattus flavipectus)、社鼠(Niviventer niviventer)、中华竹鼠(Rhizomyidae sinensis)、豪猪(Hystrix hodgsoni)。
古北界有4个分布型,其中高地型4种,包括喜马拉雅旱獭(Marmota himalayana)、高原松田鼠(Neodon irene)、中华蹶鼠(Sicista concolor)、间颅鼠兔(Ochotona cansus);古北型4种,巢鼠(Micromys minutus)、褐家鼠(Rattus norvegicus)、东亚伏翼(Pipistrellus abramus)、山蝠(Nyctalus plancyi);广布型2种,包括岩松鼠(Sciurotamias davidianus)、托氏兔(Lepus tolai)。
由此看来,白水河自然保护区小型兽类中,东洋界以喜马拉雅-横断山型种类最多,共15种。这一结果和白水河自然保护区所处的地理位置是紧密相连的。保护区地处四川盆地和青藏高原东缘,龙门山系东坡,也是中国生物多样性最丰富的横断山系的一部分。从动物地理上,白水河自然保护区处于东洋界,西南区,西南山地亚区。
3.2 白水河小型兽类群落多样性
通过表2可知,白水河自然保护区进行采样的
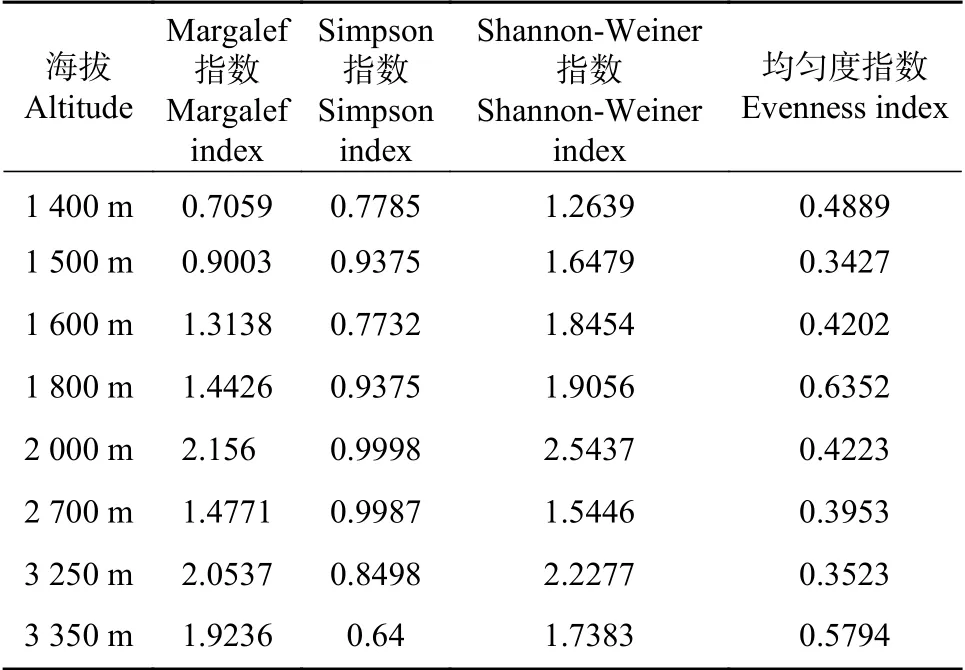
表2 不同海拔段物种多样性指数计算结果Tab.2 Calculation resultsof speciesdiversity index at different altitudes
8个海拔段中,1400 m海拔段Margalef丰富度指数最低,为0.7059;2 000 m海拔段Margalef丰富度指数最高,为2.1560;3250 m,3350 m海拔段仅次于2 000 m海拔段,分别为2.0537,1.9236;1500 m,1600 m,1800 m和2700 m海拔段Margalef丰富度指数较为接近,但结果偏低。
(续表1)
Simpson指数结果与Margalef丰富度指数结果略有不同,3350 m海拔段指数最低,为0.6400;2 000 m海拔段指数最高,为0.9998;2700 m海拔段次之,0.9987。1500 m指数为0.9375,与1800 m海拔段计算结果一致;接下来分别是3250 m,1400 m海拔段。
Shannon-Weiner指数结果和Margalef丰富度指数一致,1400 m海拔段Shannon-Weiner指数最低,为1.2639;2 000 m海拔段Shannon-Weiner指数最高,为2.5437。3250 m海拔段指数次之;1500 m,1600 m,1800 m,2700 m,3350 m海拔段指数计算结果较为接近,其中1600 m海拔段最高,2700 m海拔段最低。
但均匀度指标和前面的结果有较大不同,1800 m海拔段最高,为0.6352;1550 m海拔段最低,为0.3427。
通过采集样方物种种类和数量分析,我们可以发现1400 m,1500 m及1600 m海拔段的丰富度指数,辛普森多样性指数,香农-威纳指数均低于其他海拔段,原因在于这3个海拔段均为人工林,林下几无植被,生境单一,无法为物种提供良好的生存环境。2700 m海拔段指数较低因该工作面狭小,下夹数和陷阱数量,均少于其他海拔段,导致采集到的样品数量偏少。2 000 m海拔段丰富度指数,辛普森多样性指数,香农-威纳指数均为最高,其原因在于在该海拔段主要以阔叶林为主,生境多样性最为丰富,为物种生存提供了良好的环境。1800 m海拔段均匀度指数最高,其原因在于该海拔段物种数量和种类均为最少,但每个物种采集数量较为均衡,且数量接近,导致均匀度指数最高。
通过表3可知,白水河自然保护区进行采样的4种不同生境中,针阔混交林丰富度指数最低,1.3135;阔叶林丰富度指数最高,为2.1560;灌丛次之。
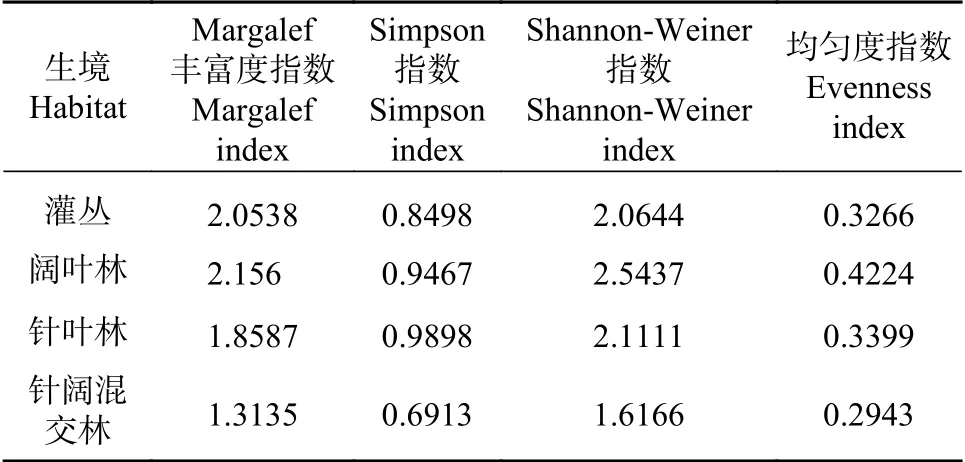
表3 不同生境物种多样性指数计算结果Tab.3 Calculation resultsof speciesdiversity index at different habitats
Simpson指数结果与丰富度指数结果一致,针阔混交林指数最低,为0.6913;针叶林指数最高,为0.9998;阔叶林指数接近针叶林,其次是灌丛。
Shannon-Weiner指数结果和丰富度指数一致,针混阔叶林香农-威纳指数最低,为1.6166;阔叶林香农-威纳指数最高,为2.5437。灌丛与针叶林指数相似,但略低于针叶林。
均匀度指标和前面的结果与丰富度指数,Shannon-Weiner指数结果一致,针阔混交林最低,0.2943;阔叶林最高,0.3523,2700 m次之。灌丛与针叶林指数相似,但同样略低于针叶林。
通过采集样方物种种类和数量分析,我们可以发现针阔混交林的丰富度指数,辛普森多样性指数,香农-威纳指数和均匀度指数均低于其他海拔段,原因在于该生境分布海拔最高,受海拔影响其生境中并无太多物种生活。阔叶林4种指数除辛普森多样性指数外均为最高,其原因在于在该生境采集到的样品种类最高,数量仅次于针叶林,且每个物种的采集数量均为均衡。
4 结论和讨论
保护区处于四川盆地向青藏高原东缘川西高山峡谷的过渡带,多种地理要素(地形地貌、气候、植物区系和植被区划等)在这里交汇,保护区自然地理要素复杂,山高谷深,气候湿润,植被类型垂直分布明显,动物组成复杂多样。保护区从海拔1400~4800 m依次覆盖着常绿阔叶林、常绿落叶混交林、落叶阔叶林、亚高山针叶林、高山亚高山灌丛、高山草甸和流石滩植被[18]。区内气候较为潮湿,植被类型丰富,有一定厚度的腐殖层,适合鼩鼱科和鼠科物种生存;海拔3200 m以上的亚高山针叶林和亚高山灌丛带中,鼠兔科物种占绝对优势,其原因在于鼠兔为植食性,主要生活在草甸、岩石、草原及斜坡环境中,亚高山针叶林和亚高山灌丛带环境更适合其生存。因此,不同的植被类型造成了保护区内物种分布的多样性。通过对采集的标本的数量分析可得,鼩鼱科和鼠科所占比例大,超过60%,超过了猬科、仓鼠科和鼠兔科数量的总和,表明在保护区内鼩鼱科和鼠科是小型兽类的优势科,这与其生存的环境存在着很大的关系。鼠科白腹鼠属和姬鼠属分别以社鼠和长尾姬鼠为优势种,所占比例远高于80%,表明鼠科物种多样性较为单一,物种在保护区类分布很不均匀,表现出优势物种占绝对优势,种间竞争大。
本次调查到几个值得关注的物种,包括大长尾鼩,斯氏长尾鼩,蹼足鼩以及鼠兔。其中,大长尾鼩于1907年由Kastschenko作为缺齿鼩鼱亚属成员而命名,Allen[19],Ellerman and Morrison-Scott[20]和Hoffmann[21]等学者均将其认为是1个单独种。该种所获标本甚少,本次调查采集到1号标本,可为后续的分类学研究提供重要的信息;斯氏长尾鼩是Thomas[22]根据Anderson 和Smith 在四川康定采集的标本命名的,并将其作为缺齿鼩鼱属的1个新种处理。除Ellerman and Morrison-Scott[19]将斯氏长尾鼩作为大长尾鼩的1个亚种处理,绝大多数学者[21,23-24]认为斯氏长尾鼩是单独种。该种所获标本也很稀少,本次调查采集到2号标本,可为后续的分类学研究提供重要的信息,且该种与大长尾鼩存在同域分布的现象,但有更为广泛的分布,因此采集标本对于这两个物种系统的研究有重要的意义;蹼足鼩属为单型属,种级分类地位稳定,现共计两个亚种:蹼足鼩指名亚种(N.e.elegans),锡金亚种(N.e. sikkimensis),He[25]通过分子生物学的研究,认为蹼足鼩属与东方水鼩属来自于同一祖先。但由于该物种数量稀少,采集样品极其困难,对其研究十分有限,蹼足鼩是鼩鼱科中最适应于水生生活的物种,对其生活习性的研究蹼足鼩族的起源和分化也具有重要的意义。分布于白水河自然保护区的鼠兔,我们发现了2种鼠兔,一种鼠兔有异耳屏,适于异耳鼠兔亚属物种,但形态上和已知的异耳鼠兔亚属的物种有差异,另外一种在外部形态上和间颅鼠兔接近,但分布海拔低,不符合间颅鼠兔的分布特点。为此进行了分子鉴定。经测定采集标本的cyt b序列,和GenBank的序列对比,下载相关序列构建系统发育树显示(见图1),异耳鼠兔亚属的物种和黄龙鼠兔最接近,但构成独立单系,和黄龙鼠兔有较大遗传距离,不排除为新种或者是黄龙鼠兔的新亚种。类似间颅鼠兔的标本确实和间颅鼠兔构成同一大支,但也是独立单系,并和间颅鼠兔有一定的遗传距离,因此,这批标本也不排除为新种或者新亚种的可能,有待进一步地研究。
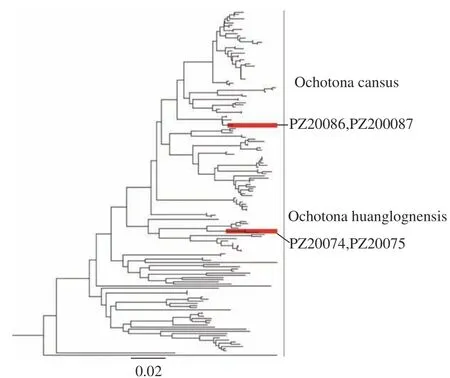
图1 关于鼠兔的系统发育树Fig.1 Phylogenetic treeof Ochotoma
2001年原四川省林业厅对白水河自然保护区进行的兽类调查中,记录到小型兽类共计4目13科63种,较此次调查4目13科44种的结果多了17种。在名录确定中,将不可能分布于该区域的物种去除,对于未采集到的物种或可能分布于该区域的物种,也未列入名录中。在翼手目的调查中,本次调查较2001年调查少了8种,因本次调查队翼手目的调查主要是通过观察和访问的方法,没有采集到标本,结果可能不甚准确。在对劳亚食虫目的调查中,本次调查较2001年调查少了4种,具体而言,依据《Handbook of the Mammals of the World–Volume 8:Insectivores》[26],北小麝鼩未分布于四川,中麝鼩未采集到,无法确定其是否存在分布,故将这两个种从名录中去除;通过采集的样品,补充斯氏长尾鼩,缅甸长尾鼩和灰腹长尾鼩3个物种;对于喜马拉雅水鼩、纹背鼩鼱、长吻鼹和长吻鼩鼹,我们并未采集到样品,对其是否分布于白水河保护区存在异议,故未列入名录中。在啮齿目的调查中,根据最新的分类结论将鼯鼠科并入松鼠科中,但并未发现灰鼯鼠和沟牙鼯鼠的分布;在鼠科中去除大耳姬鼠和小家鼠,川西白腹鼠因未采集到的样品对其分布存在疑问;在跳鼠科中去除四川林跳鼠;根据最新分类结论修订田鼠科为仓鼠科,并去除根田鼠和四川田鼠的分布,修订高原松田鼠的拉丁学名Pilymys irene为Neodon irene。在兔形目的研究中,通过分子鉴定补充间颅鼠兔和黄龙鼠兔,黄河鼠兔为达乌尔鼠兔的同物异名,其不在四川分布,故从名录中去除。传统分类学主要依靠对不同种属间形态和解剖方面的比较来决定亲缘关系,但不同种类之间的形态特征有时候并不明显,特别是食虫目的部分物种之间形态并未有明显的形态差异,或存在一定的个体变异,这导致依据形态进行的分类存在一定的错误。利用分子鉴定,构建系统发育树可以较为准确的反应物种的种类,对于分类学研究有着重要的意义。
猜你喜欢
北京航空航天大学学报(2022年8期)2022-08-31
林业与环境科学(2021年1期)2021-04-12
小学生学习指导(高年级)(2020年6期)2020-07-07
湖北农机化(2020年3期)2020-01-09
安徽农学通报(2018年8期)2018-05-30
中国诗歌(2016年6期)2016-11-25
浙江林业(2016年6期)2016-07-05
绿色中国(2015年12期)2016-01-05
阅读与作文(英语初中版)(2013年3期)2013-04-10
小学阅读指南·低年级版(2009年9期)2009-10-27
