
水稻与稻粒黑粉病菌互作分子机制研究进展
2021-11-06 02:59蒋钰琪舒新月郑爱萍王爱军
生物技术通报 2021年9期
蒋钰琪 舒新月 郑爱萍 王爱军
(四川农业大学农学院,成都 611130)
水稻不育系在高产优质杂交稻选育中发挥关键作用,是水稻高产的关键种质资源[1-2]。稻粒黑粉病由担子菌门真菌Tilletia horrida侵染引起,主要危害水稻不育系花器官[3-4],在世界杂交稻制种田中广泛分布。该菌厚垣孢子抗逆性强,在土壤和寄主种子中可长时间存活[5]。当环境条件适宜时,厚垣孢子萌发产生弹射能力较强的针状或香蕉状次生担孢子侵染寄主[6],随后在颖壳内产生大量的黑色粉状冬孢子影响水稻产量与质量[7-8]。以往认为该病发病率较低,对水稻生产影响有限,并未引起足够的重视[9-11]。目前,为了提高杂交稻制种产量,新选育的不育系柱头外露率增加,为稻粒黑粉病菌侵染提供了有利的宿主条件,导致该病害发生率逐年加重,已演变为威胁杂交稻高产优质的主要病害之一[12-13]。因此,研究稻粒黑粉病菌与寄主互作机制,进一步防控该病害的发生,对保障粮食安全生产具有重要的意义。
自Takahashi[14]于1896年报导稻粒黑粉病以来,国内外学者先后就其侵染细胞学开展了一系列研究[15-17],然而稻粒黑粉病菌与水稻互作机制研究较少,尤其是稻粒黑粉病抗性基因尚未见报道。本文综述了近年来稻粒黑粉病菌侵染过程、基因组学以及与水稻分子互作的最新进展,并提出了下一步研究的重点方向。以期为解析稻粒黑粉病菌致病机制、挖掘潜在的抗性基因;进一步通过分子育种手段培育抗病水稻不育系新材料奠定一定的理论基础,保障水稻产业的可持续发展。
1 稻粒黑粉病菌侵染过程
稻粒黑粉病菌主要侵染水稻不育系的柱头、雄蕊、浆片和雌蕊等花器官[3-4]。在开闭颖后,则主要侵染柱头的外露部分。当不育系受精后,才能形成具有黑色粉状物的病粒(图1)[9]。水稻开花初期发生侵染,盛期受害最重。花期分散、张颖角度大、柱头外露率高的不育系较容易发生侵染,且柱头活力强弱与病原菌侵染显著相关[18-20]。此外,高温多雨天气、过多施用氮肥、赶花粉、使用生长激素也利于病原菌侵染寄主[21]。病原菌形态特征方面,王爱军等[6]对稻粒黑粉病菌菌落、冬孢子及次生小孢子形态进行了观察,人工培养基上初期呈酵母状的白色菌落(图2-A),后菌落面积增大,中间隆起,颜色由乳白色变为中间颜色较深的奶酪色,表面产生辐射状褶皱(图2-B)。其冬孢子呈球形或椭圆形(图2-C),次生小孢子形态为线状和弯曲状(图2-D),单细胞核(图2-E)。

图1 水稻稻粒黑粉病症状Fig.1 Symptom of rice kernel smut
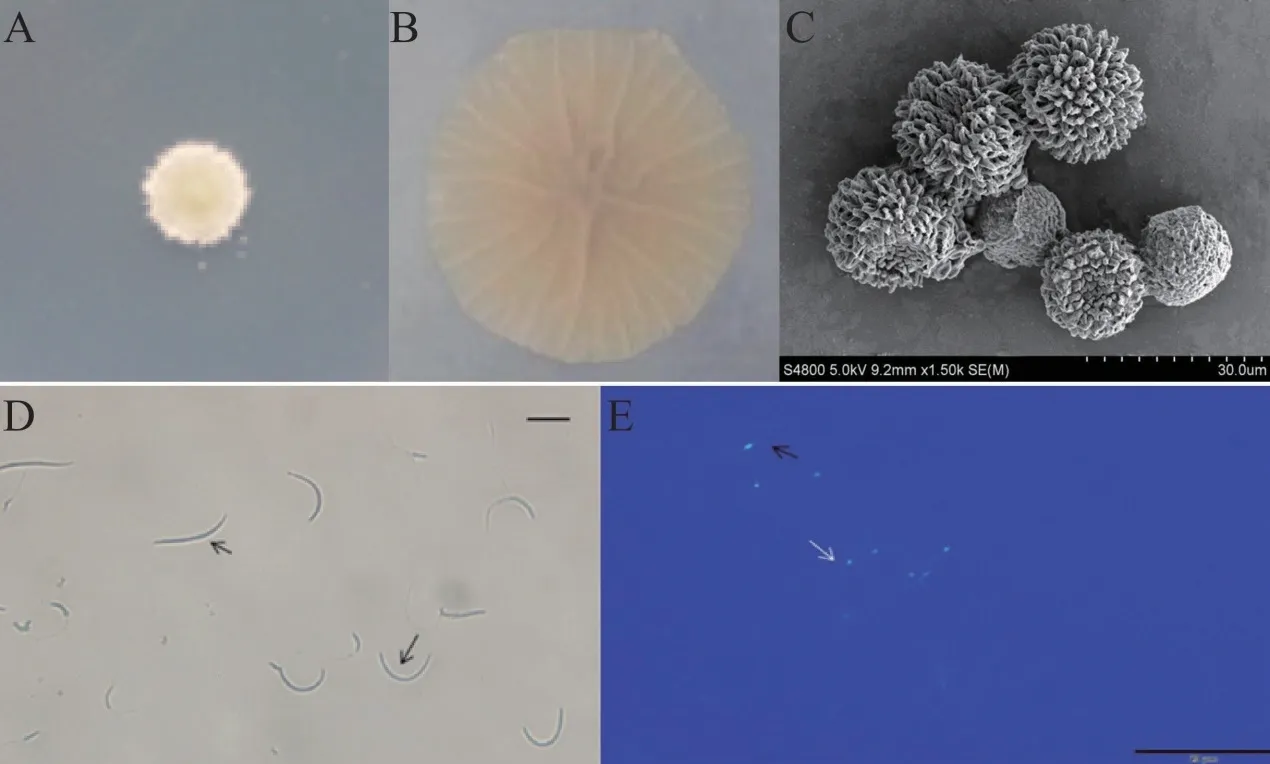
图2 水稻稻粒黑粉病菌形态特征Fig.2 Morphological characteristics of rice T. horrida
有关稻粒黑粉病菌侵染过程,国内学者做了些细胞学观察[15-17]。王中康等[22]对接种稻粒黑粉病菌组织切片进行了观察,结果表明病原菌担孢子从花柱进入子房,进而扩展到珠心组织,后在糊粉层细胞或细胞间形成近圆形的冬孢子。陶家凤等[16]分别对4个不育系珍汕97A、D90A、G46A和K17A接种稻粒黑粉病菌,观察到病菌菌丝直接侵染柱头,后扩展到子房的珠心组织,于接种8 h后在子房中发现了病原菌菌丝,第7天在种皮和糊粉层间形成了初生冬孢子,第9天冬孢子成熟,病菌在4个不育系材料中的浸染过程相似,且无需授粉就可以浸染,但只在胚乳能正常发育的子房中形成冬孢子[15]。此外,相关研究表明稻粒黑粉病菌侵染水稻花器官初期并不会破坏组织细胞,没有明显症状;近黄熟期才能出现明显的黑色粉状物。这为进行病害早期监控,及时提出防控措施带来了一定的困难。在后续的研究工作中,可进一步对优化稻粒黑粉病早期检测手段进行研究。
2 稻粒黑粉病菌的致病分子机制
作为活体营养型真菌,稻粒黑粉病菌在人工培养基上生长缓慢,平均生长速率为0.13 cm/d;且田间接种较为困难,在一定程度上阻碍了其致病机理的研究。稻粒黑粉病菌强毒力菌株JY-521的全基因组测序组装完成[23],为研究其效应蛋白提供了数据基础,并为通过病原菌效应蛋白挖掘寄主抗性蛋白奠定了重要的理论基础。
2.1 稻粒黑粉病菌全基因组测序完成
全基因组测序组装是从分子水平上阐明病原菌致病机制的前提。Wang等[24]使用第二代基因组测序技术对稻粒黑粉病菌菌株QB-1进了基因组测序组装,获得约20 Mb的基因组草图,预测获得9 038个基因,其中4 650个基因用NR库进行了注释。Wang等[23]进一步结合第二代和第三代基因组测序技术对稻粒黑粉病菌强毒力菌株JY-521进行了精细测序,组装出全长为23.2 Mb的基因组框架图,预测获得7 729个基因。此外,还对另外6个不同地理来源的稻粒黑粉病菌菌株进行了基因组重测序,并检测到了丰富的遗传变异。
进化分析表明,稻粒黑粉病菌与大麦坚黑粉菌(Ustilago hordei)具有较近的亲缘关系,但两者之间不存在明显的共线性。对碳水化合物酶、次生代谢产物及分泌蛋白编码基因进行了注释,结果表明稻粒黑粉病菌基因组中编码碳水化合物酶及次生代谢产物相关基因较少,可能反应了其独特的活体营养型寄生方式,这一结果与活体营养型真菌稻曲病菌(Ustilaginoidea virens)基因组注释结果相似[25]。在预测的597个分泌蛋白编码基因中发现,366个编码小片段分泌蛋白(<400氨基酸)的基因。此外,植物-病原互作蛋白数据库比对分析表明1 697个蛋白编码基因与病原菌致病性相关。基于比较和功能基因组学分析,稻粒黑粉病菌全基因组测序的完成为揭示其进化、致病分子机制研究及进一步有效防控该病害提供了重要信息。
2.2 稻粒黑粉病菌致病机制研究
与稻瘟病菌(Magnaporthe oryzae)和白叶枯病菌(Xanthomonas oryzae)等水稻病原菌相比,稻粒黑粉病菌田间接种及遗传转化较为困难,其致病机制研究进展缓慢。病原菌在侵染寄主早期,往往诱导其致病基因的上调表达。舒新月等[26]对稻粒黑粉病菌侵染感病水稻不育系材料9311A的5个时间点(8、12、24、48和72 h)转录组进行了分析,发现有500个基因在不同的侵染时间点被诱导差异表达,且在侵染8 h差异表达基因数最多。其中,有131 个编码小分泌蛋白基因在侵染8 h被诱导上调表达,这些基因预测为候选效应蛋白。差异基因GO(gene ontology)和KEGG(kyoto encyclopedia of genes and genomes)富集分析表明,脂类降解(lipid degradation)和自噬过程(autophagy processes)是稻粒黑粉病菌成功侵染寄主的关键生物学途径。为稻粒黑粉病菌致病机制研究提供了参考。
植物病原真菌分泌的效应蛋白在植物与病原菌互作过程中发挥重要的作用[27]。多数编码小分子分泌蛋白,通过诱导改变寄主细胞结构或功能,干扰寄主免疫,从而促进病原菌侵染[28-29]。Wang等[30]从预测的131个稻粒黑粉病菌效应蛋白中选择36个在烟草表皮细胞中进行了瞬时表达,发现 smut_2965和smut_5844可诱导烟草表皮细胞死亡,并激活烟草免疫反应。预测的信号肽是smut_5844引起烟草表皮细胞坏死反应能力是必须的,而预测的核糖核酸酶活性位点是smut_2965所必须的。这为稻粒黑粉病菌致病机制研究提供了新的视角。为进一步明确smut_2965和smut_5844在稻粒黑粉病菌致病过程中的作用,还需结合基因编辑技术敲除稻粒黑粉病菌目的基因,并验证其对病原菌毒力的影响。
上述研究为稻粒黑粉病菌致病机制的解析奠定了重要的基础。但是,由于稻粒黑粉病菌生长周期较长,研究其功能基因难度较大;稻粒黑粉病菌侵染初期不显症,季节、气候等因素对病原菌侵染影响较大,给田间菌株致病性检测带来了一定的困难;此外,稻粒黑粉病菌基因编辑体系尚不完善。这些原因导致了稻粒黑粉病菌致病机制研究进展缓慢。盘林秀等[31]对稻粒黑粉病菌不同菌株、不同生活史阶段和不同浓度 Basta 处理下的最佳内参基因进行了筛选鉴定,发现UBQ、GAPDH 和 EF1α为不同条件下的最稳定内参基因,为稻粒黑粉病菌致病基因在侵染时的表达模式研究奠定了重要基础。今后研究中,进一步完善稻粒黑粉病菌的接种方法和功能基因编辑体系至关重要,可为阐明稻粒黑粉病菌致病机理、制订有效的防控策略提供新的思路。
3 水稻抗稻粒黑粉病研究
应用和选育抗病水稻不育系是防控稻粒黑粉病最为经济、有效的手段,而关于稻粒黑粉病抗性水稻不育系资源及抗病基因报道较少。在稻粒黑粉病防控方面,Chen等[32]通过设计特异性ITS引物,可在病原菌侵染早期检测到稻谷中的黑粉病菌,便于及时做出相应的防控措施,克服了稻粒黑粉病菌侵染早期不显症的问题。邵见阳等[17]用4个稻粒黑粉病菌菌株分别接种了2个水稻不育系材料,发病结果表明4个菌株没有对某一个不育系表现亲和或不亲和性。该结果说明稻粒黑粉病菌没有明显的生理小种分化,为抗病水稻不育系田间接种鉴定奠定了重要的基础。王爱军等[33]对来源于我国福建、四川和湖北3省的78个水稻不育系材料进行了抗稻粒黑粉病评价,获得了4个表现中抗水平以上的水稻不育系材料4766A、江城3A、巨丰2A和天丰A,为稻粒黑粉病抗性研究提供了重要的抗源材料。
寄主植物与病原菌互作过程中,病原菌通过侵入、增殖和移动扩散完成对寄主的侵染;植物感知病原菌进而激活一系列的防御反应,减缓病原菌的侵染[34]。稻粒黑粉病菌强毒力菌株JY-521分别侵染抗病不育系江城3A和感病不育系9311A,在侵染8、12、24、48和72 h取样进行转录组分析。结果发现抗病水稻不育系中有更多的基因被稻粒黑粉病菌侵染诱导差异表达,表明稻粒黑粉病菌的侵染激活了抗病不育系的多个生物学途径[24,35]。在侵染早期(8 h),抗病不育系中类钙调蛋白(calmodulinlike proteins)编码基因OsCML7和OsCML14;参与活性氧(reactive oxygen species,ROS)爆发的谷胱甘肽S-转移酶(glutathione S-transferase)编码基因Os09g0467200、Os01g0369700和Os01g0949800;NADPH 氧化酶基因Osrboh9以及水杨酸(salicylic acid,SA)信号途径相关基因OsNPR1等多个基因被诱导上调表达。此外,病程相关蛋白(pathogenesisrelated proteins,PR)是植物受病原菌侵染诱导产生的一类蛋白,参与植物的诱导抗病性。转录组和荧光定量PCR分析表明,PR1a 和PR10b 也在稻粒黑粉病菌侵染抗病不育系早期被诱导上调表达。这些结果表明稻粒黑粉病菌侵染水稻初期就激活了水稻的基础防卫反应[35]。
稻粒黑粉病菌主要侵染水稻不育系的花器官。抗感不育系比较转录组分析发现,抗病不育系中水稻颖花开花调节基因OsCTR2、OsABF1和OsRR1被稻粒黑粉病菌侵染诱导差异表达,而在感病不育系中这3个基因不呈差异表达。先前研究表明OsRR1和OsCTR2 的过量表达株系表现晚花表型[36-37]。在侵染12 h时,抗病不育系中这两个基因被诱导上调表达。OsABF1是1个开花期的负调控因子,干涉表达株系表现延迟开花表型,与此对应抗病不育系中该基因被稻粒黑粉病侵染诱导下调表达。这些结果表明诱导晚花可能是稻粒黑粉病抗性的潜在机制[38]。为稻粒黑粉病抗性机制解析提供了理论基础。
此外,抗感水稻不育系转录组分析发现,有4 425个基因在抗病不育系中被稻粒黑粉病侵染诱导差异表达,而在感病不育系中不呈差异表达[35]。这些基因可能是参与稻粒黑粉病抗性调控的关键基因。在以后的稻粒黑粉病抗性基因克隆工作中,可将重点集中于这类基因。该结果为稻粒黑粉病抗性育种研究提供了潜在的基因资源。
4 总结与展望
植物病原菌致病包括从识别、侵染到产孢等多个阶段,每个阶段寄主与病原菌之间的互作涉及多个生物学途径及基因调控,是一个复杂的生物学过程。明确病原菌致病机理对防控植物病害具有重要意义。水稻稻粒黑粉病菌和稻曲病菌均为活体营养型真菌,且均侵染水稻花器官[6,39]。花丝被稻曲病菌侵染后伸长受到抑制,不能形成成熟的花粉,受精过程遇阻,淀粉不能完成积累[40]。因此,稻曲病菌在侵染水稻过程中可能通过模拟水稻胚珠受精过程,诱导灌浆相关基因(OsRISBZ1)、种子储藏蛋白基因(OsGlutln1、OsPromln2、Os03g58480)和淀粉代谢基因(OsSSIIIa、OsAGPS2b、Os02g32660)的表达,进而激活水稻灌浆信号途径,为病原菌生长提供营养物质[41-43]。相关研究也表明,完成受精的子房可以阻碍稻曲病菌菌丝形成稻曲球[40-41]。而与之相反,稻粒黑粉病菌只有在受精完成的不育系中才能形成黑色粉状的冬孢子[23],我们推测稻粒黑粉病菌致病机制与稻曲病菌模拟水稻胚珠受精过程不同,应该存在其独有的寄主-病原菌互作生物学过程。
致病功能基因研究可为稻粒黑粉病防治提供新的靶标,对促进该病的防控,提高杂交稻制种产量具有重大意义。随着稻粒黑粉病菌基因组测序组装的完成及侵染转录组数据的解析,为深入阐明其致病机制奠定了重要基础。基于比较基因组学分析,挖掘与其它病原真菌致病相关基因同源性基因,是研究稻粒黑粉病菌致病基因的有效方法。MAPK(mitogen-activated protein kinases)信号途径在植物病原真菌致病过程中具有重要的作用[44]。玉米黑粉病菌基因Ubc3是MAPK信号途径的1个关键基因,与病原菌菌丝的生长相关,还可通过调节致病性发育基因 Prf1影响病原菌致病性[45]。稻粒黑粉病菌smut_0057基因是Ubc3的同源基因[23],可能在稻粒黑粉病菌致病过程中发挥关键作用。HOG1信号途径是植物病原真菌中较为保守的MAPK 途径之一,其中的Hog1同源基因在真菌中具有较高的保守性,在维持病原菌细胞内外渗透压平衡,氧化胁迫应答、致病过程中起重要作用[46]。Hog1的同源基因Smut_0038可能是稻粒黑粉病菌致病关键基因[23]。进一步的工作可通过构建稻粒黑粉病菌突变株系阐明这些基因在水稻-稻粒黑粉病菌互作过程中的作用。
此外,基于基因组功能基因注释预测到的1 697个病原-寄主互作基因(pathogen-host interaction gene,PHI)中,部分基因是稻粒黑粉病菌特有的。这些基因可能参与了稻粒黑粉病菌的致病过程。进一步结合侵染转录组分析,预测了稻粒黑粉病菌潜在的效应蛋白,也为研究稻粒黑粉病菌新的致病功能基因提供了重要的依据。利用病原菌的致病基因挖掘寄主潜在的抗病基因是目前植物病理学领域的热点研究内容。未来工作一方面可进一步完善稻粒黑粉病菌基因编辑体系,应用基因编辑技术验证病原菌关键致病因子。另一方面,可充分利用已报道的致病相关基因,如smut_2965和smut_5844,挖掘寄主中与其相互识别的抗性靶基因。
尽管抗稻粒黑粉病水稻不育系资源的田间筛选方面有了一些进展,但其抗病相关位点及基因尚未见报道。生产上,应用抗病基因选育抗病品种是防控水稻病害最为有效的措施,然而抗稻粒黑粉病的遗传机制尚不清楚,阻碍了其抗性品种的选育。在进一步的工作中,可利用现有抗病材料构建遗传分离群体,通过高通量测序技术手段定位稻粒黑粉病抗病位点,进而克隆抗病基因。抗病基因研究最终意义是培育抗病品种。虽然基因组技术的不断发展为抗病基因的挖掘提供了技术支撑,但如何将抗病基因应用到实际生产中才是科研工作最为关键的。制种产量较高的水稻不育系通常具有高异交性,柱头外露率较大,大大提高了稻粒黑粉病的感病性[7,12]。结合分子生物学手段和常规杂交育种技术,已报道的稻粒黑粉病抗性水稻不育系4766A、江城3A和巨丰2A培育高异交抗病不育系[32],进一步提高杂交稻制种产量是我们未来工作的核心目标。
猜你喜欢
今日农业(2022年4期)2022-06-01
当代水产(2022年1期)2022-04-26
中国农业科学(2021年23期)2022-01-14
中国瓜菜(2020年8期)2020-09-26
作物研究(2020年2期)2020-04-04
发明与创新·大科技(2019年5期)2019-07-31
农民致富之友(2018年4期)2018-04-22
农民致富之友(2017年4期)2017-04-10
吉林农业(2014年8期)2014-08-20
农民致富之友·下半月(2013年11期)2013-01-13
