
核酸适体靶向的膜蛋白识别与功能调控研究进展
2021-11-15 12:47刘学娇张春娟刘巧玲
高等学校化学学报 2021年11期
刘学娇,杨 帆,刘 爽,张春娟,刘巧玲
(湖南大学生物学院,长沙410082)
膜蛋白是一类重要的生物活性分子,其功能异常是导致包括心血管疾病、肿瘤、代谢性疾病和神经变性疾病等重大疾病的主要机制[1].研究并调控细胞膜蛋白的结构和功能有助于阐明生命活动的基本规律,为开发针对相应疾病的药物以及构建高性能生物传感器提供理论依据.采用化学方法调控蛋白的结构及功能具有操作简单、效率高等优势,受到研究者的广泛关注,相关技术已被用于研究蛋白相互作用等领域[2~4].研究表明,以特定小分子作为诱导试剂,利用化学诱导法可以实现蛋白功能的精确调控.例如,Spencer等[5]发现利用雷帕霉素衍生物可诱导T细胞表面嵌合蛋白二聚,从而调控下游信号通路.然而,目前应用于蛋白功能调控的小分子化合物种类十分有限[6].此外,大部分化学诱导法的研究体系需要使用经过基因改造的嵌合蛋白,同时还需要保持嵌合蛋白的生物活性,技术难度较高.因此,发展操作简单,无需基因改造,可以在生物友好条件下快速、可逆、特异性调控蛋白功能的新方法在研究膜蛋白生物学功能及相关生物医学应用等方面具有重要意义.
核酸适体是通过指数富集配体系统进化(SELEX)的组合筛选从包含多达1015个随机序列的文库中分离获得的单链寡核苷酸[7~9].核酸适体通过自身折叠成特定结构与目标分子结合[10].因此,针对特定目标分子,利用SELEX筛选技术获得的核酸适体具有高亲和力和高特异性的特点.此外,核酸适体具有成本低、易于功能化修饰[11]以及靶标范围广泛等优势.采用合适的筛选技术,通过多轮筛选即可获得特异性结合金属离子、小分子、肽、蛋白质、细胞器、病毒甚至细胞的核酸适体[12,13],由此获得的核酸适体被广泛用于生物传感、生物成像及生物功能调控等领域[14].
由于核酸适体是一种具有识别能力的功能核酸,因此,基于碱基互补配对原理可以方便地将核酸适体与DNA纳米技术相结合,借助核酸适体特异性识别目标分子的特点以及DNA分子可程序化设计、可功能化修饰等优势,拓展核酸适体在膜蛋白识别与功能调控中的应用,为研究膜蛋白相互作用提供新途径.本文介绍了基于核酸适体靶向的DNA纳米技术在膜蛋白识别与功能调控中的研究进展.首先简要介绍核酸适体识别膜蛋白的作用特点以及利用DNA纳米技术调控膜蛋白相互作用的几种方式;然后阐述靶向膜蛋白的核酸适体对细胞功能的调控;最后对核酸适体靶向的膜蛋白识别及功能调控面临的挑战进行分析,对其应用前景进行展望.
1 核酸适体靶向的膜蛋白识别
1.1 基于Cell-SELEX技术的核酸适体筛选与膜蛋白识别
利用SELEX技术可以从溶液中筛选得到特异识别小分子、多肽和蛋白的核酸适体.而活细胞筛选(Cell-SELEX)技术的开发则将核酸适体的靶标范围拓展到活细胞表面的生物分子[15].Cell-SELEX技术是在指数富集的配体系统进化(SELEX)的基础上,以活细胞为研究对象,通过筛选获得与特定靶细胞结合的核酸适体的技术[图1(A)][16].该技术可在目标分子特征不明确的情况下为靶细胞筛选核酸适体.基于此,利用筛选得到的核酸适体捕获与核酸适体特异结合的靶标分子[17].Tan等[18]筛选到可特异识别胰腺癌细胞膜蛋白转铁蛋白受体(CD71)的核酸适体XQ-2d、可特异识别鼻咽癌细胞膜蛋白(CD109)的核酸适体S3[19]及可特异识别急性T淋巴细胞白血病细胞膜蛋白酪氨酸激酶-7(PTK7)的核酸适体Sgc8等[20].通过筛选得到的核酸适体与靶标膜蛋白之间具有亲和力强、特异性和稳定性好的特点.如,核酸适体Sgc8与其靶标蛋白PTK7之间的结合稳定性好,亲和力强(Kd=0.8 nmol/L、解离力为46 pN)[21,22];核酸适体XQ-2d与其靶标CD71之间的亲和力(Kd=50.5 nmol/L)甚至比CD71抗体与CD71之间的亲和力(Kd=635 nmol/L)更强[18];核酸适体NX1838与其靶标VEGF165之间的亲和力强(Kd=50 pmol/L)、稳定性好,可在含有培养基的人脐静脉内皮细胞表面稳定存在20 h[23].这些特异识别膜蛋白的核酸适体的发掘以及分析方法的建立为核酸适体靶向的膜蛋白功能分析和调控奠定了基础.
此外,借助核酸适体与膜蛋白的特异性识别作用,以核酸适体为向导,利用简单的化学反应可以将生物分子(如单链DNA)与膜蛋白结合,从而实现核酸适体介导的膜蛋白靶向识别与功能化修饰.Cui等[24]发展了核酸适体模板合成法(Aptamer-templated synthesis,ATS)用于细胞膜蛋白的特异性识别和功能化修饰.他们利用碱基互补的原理,将核酸适体与修饰有活性反应基团的单链DNA相结合.核酸适体与膜蛋白的特异性识别作用使修饰在单链DNA上的活性反应基团与膜蛋白的氨基酸残基相互靠近并发生反应,从而实现单链DNA对膜蛋白的识别[图1(B)].该研究为DNA分子选择性修饰蛋白质结合位点提供了简单有效的方法,结合DNA纳米技术,可实现对蛋白结构和功能的人工操控.
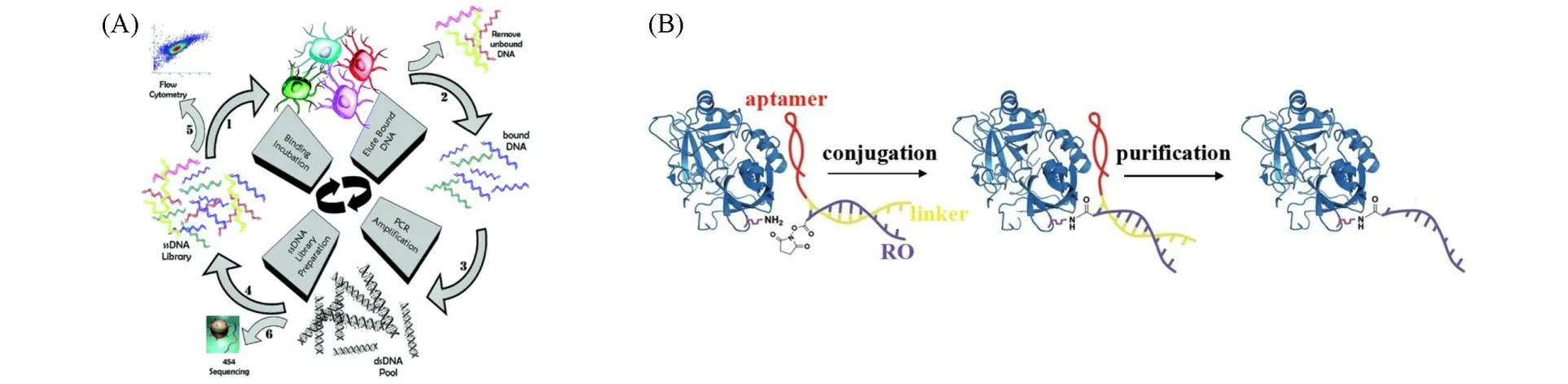
Fig.1 Scheme of aptamer screening utilizing cell⁃SELEX(A)[16]and membrane protein recognition and functional modification mediated by aptamer(B)[24]
1.2 基于DNA纳米技术的核酸适体靶向膜蛋白识别与信号放大
基于核酸适体的质谱流式细胞技术[25]、电化学技术[26~29]等由于其自身独特的优势已被用于膜蛋白分析研究.如,Mironov等[30]开发了基于核酸适体的质谱流式细胞技术.在传统流式细胞技术的基础上,引入生物素标记的特异性识别膜受体PTK7的核酸适体Sgc8、金属165Ho标记的中性亲和素、金属141Pr标记的PTK7抗体,当靶标细胞存在时,核酸适体Sgc8和抗体PTK7的金属元素分析呈现明显的相关性,可避免传统流式细胞技术中光谱重叠问题,从而实现靶标细胞的鉴定和检测.Gai等[31]开发了基于生物燃料细胞(BFC)的超灵敏自供电电化学细胞传感器.当带负电荷的CCRF-CEM细胞被负极核酸适体识别捕获时,负极产生的空间位阻以及静电斥力能有效阻止探针和阴极表面之间的电子转移,进而引起BFC输出功率的降低,从而灵敏地检测急性白血病CCRF-CEM细胞.近年来,DNA纳米技术领域发展迅猛,除了静态DNA纳米结构外,动态DNA纳米技术因其优越的可控性及功能性而成为研究热点[32].将DNA纳米技术与核酸适体相结合,发展基于DNA纳米技术的核酸适体靶向的膜蛋白识别与分析检测技术,有助于研究生命活动和疾病发展过程中的相关分子作用机制.
杂交链式反应(HCR)是一种基于DNA链取代反应的等温信号放大技术.在HCR体系中,靶分子引发两种DNA茎环交替开环,自组装得到包含大量重复单元的线性双链DNA纳米结构,具有恒温、免酶、放大效率高等优点[33].将HCR与核酸适体结合可以实现膜蛋白的特异性识别与信号放大[34,35].如,将末端修饰HCR触发链的核酸适体Sgc8作为识别单元,一旦核酸适体Sgc8与靶标PTK7结合,识别单元暴露出的HCR触发链即可引发膜蛋白表面HCR反应,实现膜蛋白PTK7的识别与信号放大[图2(A)][36].DNA链置换反应是利用DNA分子单链间的碱基互补配对原理,通过与引发链反应,释放出DNA单链产物.DNA链置换反应具有自引发性、灵敏性和准确性等特点[37].将DNA链置换反应与核酸适体相结合可以实现膜蛋白的动态识别与信号放大[38].如,Liang等[39]发现,通过细胞膜表面CY5荧光的有无实可现膜蛋白的动态识别.当细胞膜表面无膜蛋白Met存在时,溶液中双链DNA中CY5荧光被其互补链DNA中的淬灭基团淬灭;当细胞膜表面存在Met蛋白时,FAM荧光基团标记的单链DNA连接的核酸适体作为识别单元识别膜蛋白Met,同时溶液中双链DNA中CY5标记的DNA竞争性结合核酸适体,导致淬灭基团标记的DNA被释放,CY5荧光恢复,此时细胞膜显示FAM、CY5两种荧光,实现了细胞膜表面Met蛋白的动态识别[图2(B)].
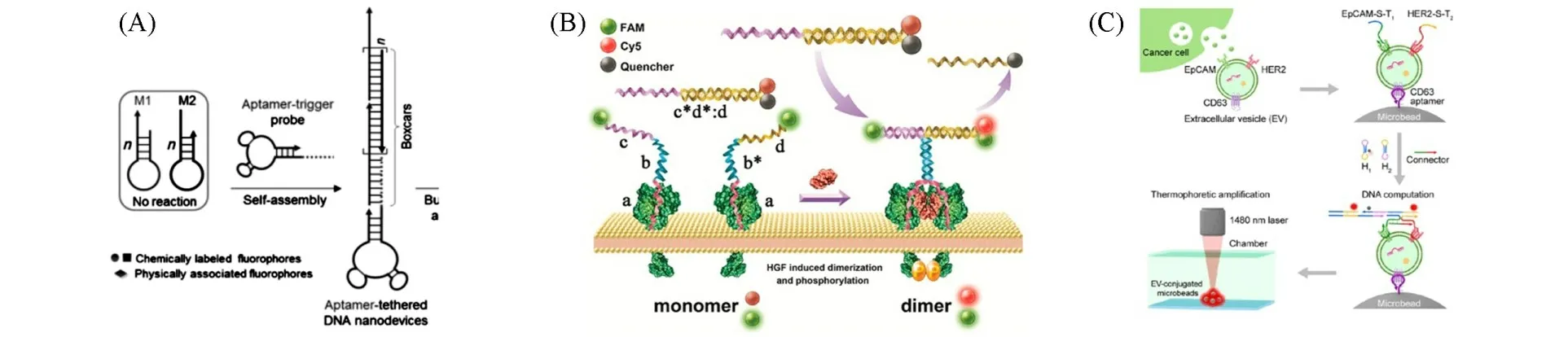
Fig.2 HCR reaction based on aptamer targeting used for membrane protein recognition and signal ampli⁃fication(A)[36],DNA strand displacement reaction based on aptamer targeting used for membrane protein recognition and signal amplification(B)[39]and DNA logical computation based on aptamer targeting used for membrane protein precise detection and signal amplification(C)[42]
DNA逻辑运算利用DNA分子碱基互补配对的性质,将所要处理的问题编码为特定的DNA分子链,当输入的DNA分子链与作为开关的特定DNA分子链进行碱基互补配对时,将产生输出DNA分子链,且输出DNA分子链的浓度与输入DNA分子链的浓度呈正相关,上述过程称为DNA逻辑运算[40].结合核酸适体,将核酸适体对膜蛋白的特异性识别作为输入信号,当多种核酸适体同时识别细胞表面对应膜蛋白时,触发核酸适体介导的DNA链杂交反应,整合放大输入信号作为输出信号,实现膜蛋白的精准识别与信号放大[41].如,Li等[42]将胞外囊泡中膜蛋白泛肿瘤蛋白EpCAM的核酸适体EpCAM-S-T1和乳腺癌标志物膜蛋白HER2的核酸适体HER2-S-T2相结合,当膜蛋白EpCAM和HER2同时存在时,整个反应系统执行“AND”的指令,在杂交链DNA(Connector)的驱动下触发HCR反应,实现膜蛋白的精准识别与信号放大[图2(C)].
2 核酸适体靶向的膜蛋白分析与功能调控
2.1 基于核酸适体靶向识别的膜蛋白分析
荧光技术通常被用于示踪和分析膜表面受体二聚或蛋白修饰[42].目前,大多数荧光技术依赖于蛋白受体的遗传改造,即体外拼接荧光蛋白与膜蛋白的基因,通过转染技术导入细胞,借助细胞自身蛋白表达系统表达融合蛋白.然而,这种策略虽然标记方便,但对膜蛋白结构和功能有一定的影响,例如会导致膜受体二聚化效率降低等[43].因此,研究者开发多种非遗传改造策略用于膜蛋白的结构、功能调控.其中,基于蛋白的非遗传改造策略虽然具有蛋白与膜受体间空间构象天然匹配等优势,但存在蛋白活性低、蛋白/多肽稳定性低等问题;基于小分子的非遗传改造策略虽然具有小分子灵活设计、合成的优势,但存在小分子种类有限、通用性低等问题;基于物理刺激的非遗传改造策略虽然具有远距离控制、时空调控等优势,但存在生物相容性低、副作用大等问题,上述问题限制了这些方法在膜蛋白功能调控中的应用[44,45].相比而言,核酸适体稳定性好、生物相容性好[46]、兼容性强,易与DNA纳米技术相结合[47],可作为新型分子工具高效识别膜蛋白并灵活调控膜蛋白的结构和功能,在膜蛋白分析与功能调控方面具有潜在应用价值.现阶段,核酸适体与膜蛋白的特异识别有助于开发新型分析检测技术,用于获取膜蛋白在细胞膜表面的相关信息,如,膜蛋白密度、膜蛋白分布状态等.Chen等[48]利用荧光染料标记的核酸适体实现了对膜蛋白的特异性标记,并依据膜表面荧光强度的变化分析细胞表面膜蛋白的密度及其分布情况.基于生物正交反应位点特异性标记、生理条件下反应高效等优点,研究人员开发了一种核酸适体诱导的蛋白特异性生物正交修饰技术,用于生物大分子(包括蛋白质、核酸、糖类等)结构、功能和相互作用研究[49].Chen等[50]利用化学基团Ac4ManNAz修饰的核酸适体作为识别单元,诱导Ac4ManNAz与细胞膜糖蛋白中的氨基发生生物正交反应,实现了对膜蛋白的特异性标记.Ambrosetti等[51]将特异识别膜蛋白的多种核酸适体进行组合,对其相应靶标膜蛋白(HER2、HER3、EGFR)进行识别.他们利用DNA纳米技术将HER2膜蛋白纳米簇中的组分信息转变为DNA序列信息,从而获得HER2膜蛋白纳米簇的组分构成及其相应的时空排布等信息.
2.2 核酸适体靶向的膜受体蛋白功能调控
细胞膜上的膜蛋白簇作为细胞信号转导的重要调控因子,在细胞对外界刺激的响应中发挥着重要作用.免疫细胞中由不同种类固有免疫受体形成的信号簇是宿主-病原菌相互作用的重要调控机制,可作为多组分装置用于病原体的检测以及免疫响应调控[52].在T细胞识别抗原过程中,外源信号触发单个T细胞受体聚集成簇促使T细胞活化,增强免疫应答[53].受此启发,研究人员以核酸适体为T细胞受体识别单元,利用DNA纳米技术诱导T细胞表面受体形成蛋白簇.McNamara等[54]通过将多价核酸适体与小鼠T细胞表面的T细胞共刺激受体蛋白4-1BB结合,体外诱导T细胞表面4-1BB蛋白成簇.Li等[55]在核酸适体末端引入可互补的DNA链,利用DNA链之间发生的DNA链杂交反应诱导膜受体蛋白Met聚集.基于此,Chen等[56]利用特异识别Met的核酸适体和特异识别CD71的核酸适体,采用DNA链置换反应诱导膜受体蛋白Met与CD71结合,从而调控膜受体蛋白Met在细胞膜上的分布状态[图3(A)].
膜受体蛋白作为细胞与外界沟通的桥梁,是细胞生命活动的门控开关.目前,针对膜蛋白相互作用的研究方法,Snider等[57]对利用膜酵母双杂系统在生物体内研究膜蛋白相互作用的相关工作进行了综述;Ma等[58]对利用邻近标记方法研究细胞间膜蛋白相互作用的相关工作进行了综述;Fan等[59]对利用不同类型识别单元以及DNA纳米技术构筑细胞表面受体以及调控细胞功能的相关工作进行了综述.本文则从核酸适体的特异性识别性能出发,聚焦利用DNA分子工具调控膜蛋白结构和功能的相关研究,对该领域的研究工作进行综述.如,利用核酸适体特异识别目标分子的特性,研究者将核酸适体作为膜蛋白配体类似物,用于调控膜受体蛋白功能,从而激活/抑制下游信号通路.Ueki等[60,61]将核酸适体作为肝细胞生长因子HGF的类似物,利用核酸适体对Met的特异性识别,诱导Met二聚,激活下游Erk或Akt信号,抑制细胞凋亡,促进细胞增殖、分化、迁移.为了进一步提高核酸适体在体内的稳定性,该课题组[62]将DNA链杂交反应与核酸适体TD0结合,组装成功能类似于碱性成纤维细胞生长因子(bFGF)的DNA纳米组装体,利用核酸适体TD0与纤维生长因子受体(FGFR)的特异性识别,介导FGFR二聚,触发FGFR磷酸化,选择性激活PKC信号通路、Ras/Raf/MEK/ERK信号通路、JAK/STAT信号通路,诱导多能干细胞的自我更新及多能性恢复[图3(B)].
在细胞膜受体蛋白中,膜受体蛋白Met功能异常通常与细胞增殖、侵袭和转移有关,已成为肿瘤治疗的重要靶点[63].以Met作为研究对象,研究人员采用核酸适体靶向膜蛋白识别策略操控膜受体蛋白Met的聚集状态进而调控其蛋白功能.如,Wang等[64]利用特异性识别膜蛋白Met的核酸适体Apt-Met和特异性识别膜蛋白TfR的核酸适体Apt-TfR,借助DNA杂交反应,诱导膜蛋白Met与TfR二聚,阻碍下游信号分子Erk和Apt磷酸化,抑制细胞增殖和迁移[图3(C)].
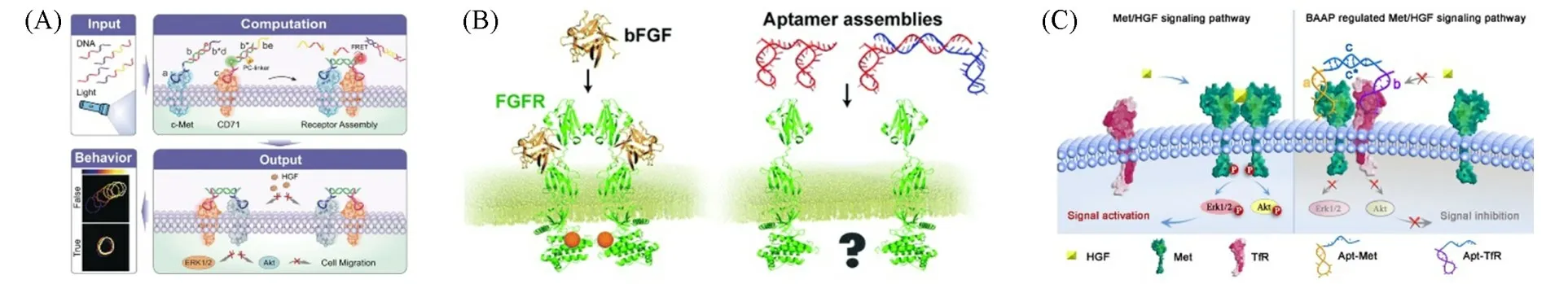
Fig.3 Analysis and regulation of membrane protein based on aptamer targeting(A)[56],regulation of dimerization of membrane receptor proteins based on aptamer targeting(B)[62]and regulation of cell proliferation and migration based on aptamer targeting(C)[64]
虽然上述研究证实核酸适体可作为配体类似物用于激活膜受体蛋白,但复杂的细胞乃至活体环境往往不利于核酸适体高效、精准识别和调控膜蛋白功能.对此,研究者开发了基于核酸适体靶向的光操控膜蛋白活性的新策略,从而实现了膜蛋白的高效识别与功能调控.Chen等[65]在核酸适体与DNA纳米技术联用的基础上,利用DNA链修饰的光敏基团(PC Linker)对紫外光的响应,触发膜受体蛋白Met表面DNA链杂交反应,诱导Met二聚,激活c-Met信号通路,促进细胞增殖、迁移.虽然上述研究工作验证了基于核酸适体靶向的光控策略可以有效调控细胞功能,但是紫外光穿透深度有限,无法对深层组织中的细胞发挥作用.针对此问题,Wang等[66]开发了利用近红外光调控膜蛋白活性及细胞功能的新策略,他们首先在金纳米棒表面修饰单链DNA,利用近红外光照射产生的光热效应释放金纳米棒表面修饰的单链DNA,借助单链DNA与特异性结合RTK的核酸适体之间的杂交反应,触发膜表面受体蛋白RTK二聚,激活RTK介导的信号通路,促进细胞极化和迁移,实现活体水平上细胞行为的精准调控.此外,利用核酸适体对靶蛋白的识别性能,研究者已成功实现表皮生长因子EGFR、EGFR和整合素等膜受体蛋白介导的细胞功能调控.Ramaswamy等[67]报道了二价核酸适体调控血管内皮生长因子-2(VEGFR2)二聚的策略.VEGFR2结合二价核酸适体形成二聚体,发生自磷酸化而活化,激活下游Akt信号通路,增强内皮细胞内NO合酶活性,促进内皮细胞毛细血管的生成.Baig等[68]报道了基于核酸适体的DNA纳米装置调控膜蛋白EGFR和integrin(α6β4)活性的策略.利用核酸适体aptamer-1对膜蛋白integrin(α6β4)特异性识别的特点,设计可同时靶向膜蛋白EGFR和integrin(α6β4)的DNA纳米装置,激活膜蛋白EGFR及integrin(α6β4)活性,上调磷酸化的integrin-β4、EGFR、Akt和Erk1/2蛋白表达水平,引起细胞收缩,同时启动生化反应通路,改变细胞分裂、内吞及胞吐过程,进而影响细胞的增殖和凋亡.这些研究显示了核酸适体与DNA纳米技术联用策略在细胞功能调控中潜在的应用价值.
3 总结与展望
细胞膜蛋白是一类重要的生物分子,在细胞与外界环境的物质交换、能量转换和信息传递等生命过程中发挥着重要作用.作为具有识别能力的功能核酸,核酸适体与DNA纳米技术的有机结合为精准、高效调控膜蛋白的结构和功能提供了新策略,有助于研究生命活动和疾病发展过程中的分子机制.核酸适体靶向膜蛋白识别与功能调控在药物开发和疾病治疗等方面具有广泛的应用前景.然而,拓展核酸适体靶向的膜蛋白功能调控的生物医学应用仍面临核酸适体在体内易被核酶降解、稳定性差及筛选周期长等问题.另外,目前可特异识别膜蛋白的核酸适体数量仍十分有限,一些具有重要研究意义的蛋白仍然缺乏相应的核酸适体用于开展研究.发展提高核酸在体内稳定性的新方法、开发高效获取疾病相关膜蛋白的核酸适体的新技术以及将核酸适体靶向调控疾病相关膜蛋白功能的策略拓展到临床应用有望成为未来重要的发展趋势.
猜你喜欢
中国慈善家(2022年3期)2022-06-14
保健医苑(2022年5期)2022-06-10
现代苏州(2022年9期)2022-05-26
快乐语文(2021年34期)2022-01-18
中国临床医学影像杂志(2021年6期)2021-08-14
肝博士(2020年5期)2021-01-18
中国(俄文)(2020年8期)2020-11-23
哈尔滨医药(2016年3期)2016-12-01
医学研究杂志(2015年7期)2015-06-22
癌变·畸变·突变(2015年4期)2015-02-27
