
土壤水分和植株密度互作对垂穗披碱草地下营养繁殖及生物量的影响
2021-11-22 12:29王玲玲库努都孜阿依吐鲁洪孟广飞郭正刚
草业学报 2021年11期
王玲玲,库努都孜阿依·吐鲁洪,孟广飞,郭正刚
(兰州大学草地农业生态系统国家重点实验室,兰州大学农业农村部草牧业创新重点实验室,兰州大学草地农业教育部工程研究中心,兰州大学草地农业科技学院,甘肃 兰州730020)
垂穗披碱草(Elymus nutans)是青藏高原地区天然草地的主要优势禾本科植物[1],根系发达,生长期长[2],地下繁殖能力和可塑性较强,不仅是高寒地区天然草地的优良牧草组分[3],而且是青藏高原地区退化草地修复和栽培草地建植的主要草种[4]。大面积的垂穗披碱草栽培草地不仅为青藏高原地区畜牧业生产提供物质资料[5],而且逐渐成为该地区草牧业经济发展的基石[4]。然而栽培草地具有自然衰退的特征[6],主要是种群自我更新能力弱所致[1]。虽然垂穗披碱草能够实现种子繁殖[4],但青藏高原地区海拔高、温度低、生长季短,在这种严酷的环境下,垂穗披碱草栽培草地更倾向于营养繁殖[7],实现其通过自我更新而维持持续生产的目标[8]。植物地下营养繁殖是植物营养繁殖的核心,被视为栽培草地种群自我更新和维持持续生产的驱动力[9]。植物地下营养繁殖主要通过克隆繁殖和芽库繁殖适应严酷环境而维持其自我更新[10−11],其中克隆繁殖能力经常用克隆小株数和传播距离测度[12],而芽库繁殖能力则用地下芽数量或密度测度[13]。因此,研究垂穗披碱草克隆繁殖和芽库繁殖,是青藏高原地区垂穗披碱草栽培草地维持稳产和持续利用的需求。
植物克隆繁殖和芽库繁殖能力不仅受水分和光照等环境因子的影响[14],而且受植株密度的约束[15]。植株密度的变化能引起个体间的相互作用[16]。栽培草地密度往往具有自疏特性[17],出现个体变大而密度降低趋势,表现出因植株密度降低而栽培草地呈现退化的现象,然而栽培草地又具有产量最后恒值法则[18],即植物个体密度变化引起的种群生物量波动,往往会通过种群的自我调节而逐渐弥补。土壤水分是植物生长的重要限制因子[19]。植株密度与土壤水分的关系既能互惠,又能拮抗。植株密度过大,会加快植物蒸腾作用;植株密度过小,地表蒸发过大,会影响土壤水分利用效率[20]。因此,只有植株密度和土壤水分耦合时,才能既维持植物稳产,又提高水分利用效率。虽然已有研究证实了土壤水分和植株密度对垂穗披碱草单株地上生物量有显著的互作效应[18],但土壤水分和植株密度互作对垂穗披碱草地下营养繁殖特征是否产生影响,尚需要科学试验提供证据。因此,本研究采用盆栽试验,分析土壤水分和植株密度互作对垂穗披碱草克隆繁殖和芽库繁殖能力以及生物量的影响,不仅为揭示土壤水分和植株密度互作影响垂穗披碱草单株地上生物量的机制提供了重要信息,而且为青藏高原地区天然草地合理管理和垂穗披碱草栽培草地生产力的持续维持提供了理论依据。
1 材料与方法
1. 1 试验设计
采用盆栽试验,设计为双因素处理,两个因素分别是土壤水分和植株密度。选用口径长24 cm,宽10 cm,底径长20 cm,宽10 cm,高12 cm 的聚乙烯塑料方形花盆,土壤水分设计为3 个水平,分别为土壤饱和含水量的30%,50% 和80%,分别用W1,W2,W3表示,植株密度设计为4,8,12 株·盆−13 个水平,分别用R1,R2,R3表示,共计9 个处理,即W1R1,W1R2,W1R3,W2R1,W2R2,W2R3,W3R1,W3R2,W3R3,每个处理6 个重复,共计54 个花盆。
垂穗披碱草种子来自青海省畜牧科学院,试验地点为兰州大学榆中校区的智能温室,白天室内温度保持在25 ℃,夜间保持在15 ℃,光照强度>600 μmol·m−2·s−1。试验采用的土壤pH 为7. 4,有机质含量为0. 66%,全氮含量为0. 11%,速效磷含量为57. 66 mg·kg−1,速效钾含量为240 mg·kg−1[5]。试验于2019 年5 月16 日开始,首先,向另外的4 个盆中装入2. 16 kg 的土,测定土壤的饱和含水量;其次,向54 个盆中装入2. 16 kg 的土壤,根据土壤饱和含水量的60% 向每个花盆缓慢浇水,使花盆内土壤水分相对均匀分布;第三,每个花盆完成浇水后,于翌日(2019 年5 月17 日)播种,每个花盆播种籽粒饱满的种子20 粒,此后每隔2 d 通过称重法监测土壤水分,让土壤水分继续维持于土壤饱和含水量的60%,同时观测种子发芽情况;第四,根据垂穗披碱草种子发芽检测标准,11 d 认为其完成发芽[21],即2019 年5 月29 日开始按照试验设计调整每个花盆的土壤水分,让其达到试验设计的水平;第五,各个花盆土壤水分达到试验设计水平后,即2019 年5 月30 日开始采用手工间苗法,将各个花盆的植株密度调整到试验设计水平。此后至试验结束,每隔2 d 继续采用称重法控制土壤水分到试验设计水平。试验期间,每隔7 d 随机挪动花盆位置,让每个花盆的植物生长环境相对均匀一致。
1. 2 指标测定
根据垂穗披碱草生长习性[22],2019 年10 月1 日取样。首先,针对花盆内的每株植株,用手轻轻地拨开植株根茎处的表层土壤,让该植株的根茎充分暴露出来,然后沿着每条根茎的方向,确定其是否有克隆小株,最后记录每株植物的所有克隆小株数;其次,每株植物有多个克隆小株,采用直尺测定各个克隆小株之间的直线距离,将其中最大的距离,记录为这个植株的传播距离;第三,测定完克隆小株和传播距离后,将花盆中每个植物的地上和地下部分整体取样(每个植物包括其所有克隆小株),用清水轻轻冲洗植株地下部分附着的土壤,冲洗干净后,采用电子显微镜,鉴定每株植物地下部分附着的地下芽,然后根据地下芽附着的位置,将地下芽分为分蘖芽和根茎芽,统计每株植物上的分蘖芽和根茎芽数量(实际鉴定地下芽时,因垂穗披碱草生长于温室,尚未发现根茎芽);最后,用剪刀将每株植物的地上和地下部分分离,其中地下芽属于地下部分,分别装入信封袋中,做好标记,在65 ℃条件下烘干48 h 至恒重,待冷却至室温时再称重。
1. 3 数据处理
首先,检验每个指标数据的正态性和同质性,若数据不服从正态分布,则采用log 进行数据转化,然后分析。采用一般线性模型对每个指标做双因素方差分析,其中将土壤水分和植株密度设置为固定因子,克隆小株数、传播距离、地下芽数量和生物量设置为响应变量,当方差分析结果显著时(P<0. 05),采用Duncan 法进行多重比较分析,以确定各处理间的差异;若土壤水分和植株密度交互作用显著时,采用MATLAB 2016b 构建二元二次模型,确定交互作用显著变量对土壤水分和植株密度的响应过程。最后,采用皮尔森相关性分析确定克隆小株数、传播距离、地下芽数量和生物量之间的关系。用SPSS 22. 0 进行双因素方差分析和皮尔森相关性分析。
2 结果与分析
2. 1 克隆小株数和传播距离
植株密度、土壤水分以及二者互作对垂穗披碱草克隆小株数和传播距离均有极显著影响(P<0. 01)。随土壤水分增加,克隆小株数呈先增加后减小的趋势,传播距离呈增加趋势。随植株密度增加,克隆小株数和传播距离均呈减小趋势(表1)。
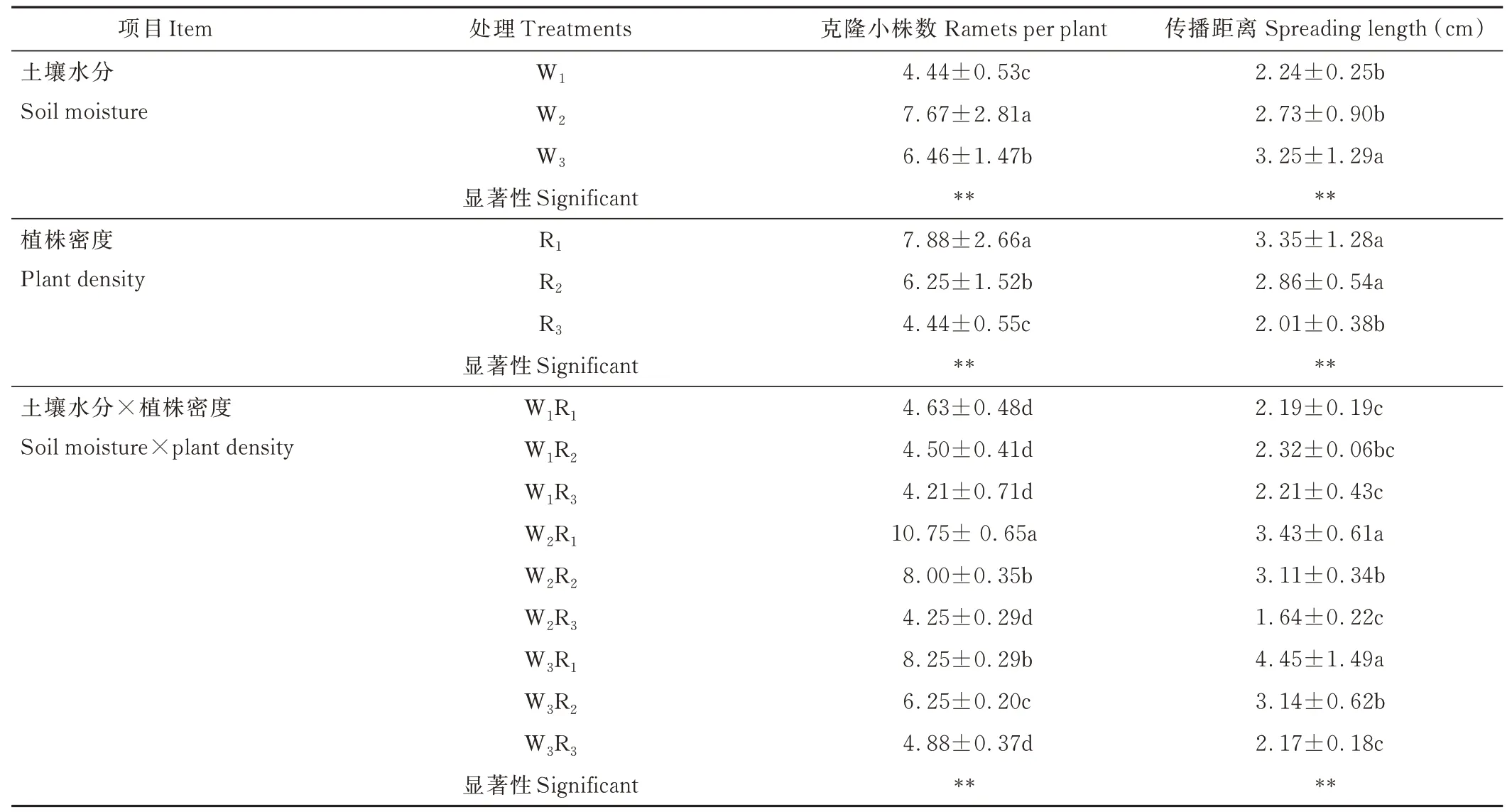
表1 土壤水分和植株密度对垂穗披碱草克隆小株数和传播距离的影响Table 1 Effects of soil moisture and plant density on ramets per plant and spreading length
克隆小株数F(x2)与土壤水分(x1)、植株密度(x2)的关系为F(x1,x2)=−5. 76+0. 5294x1−0. 06079x2−0. 004027x12−0. 0225x22−0. 01158x1x2,F 检验时,P=0. 0001,R2=0. 970,说明土壤水分(x1)、植株密度(x2)与克隆小株数具有显著回归关系。x1的一次项系数为正数,x2的一次项系数为负数,且土壤水分(x1)系数绝对值大于植株密度(x2)系数绝对值,表明在一定范围内土壤水分增加克隆小株数的效应要大于植株密度减少克隆小株数的效应;x1、x2的二次项系数均为负数,表明该面为一个开口向下的抛物面(图1A),说明垂穗披碱草克隆小株数在土壤水分和植株密度互作时理论上存在最佳组合,为W2R1。
传播距离F(x1,x2)与土壤水分(x1)、植株密度(x2)的关系为F(x1,x2)=−0. 2551+0. 07884x1+0. 5946x2−0. 0001x12−0. 04375x22−0. 01091x1x2,F 检验时,P=0. 007,R2=0. 702,说明土壤水分、植株密度与传播距离具有显著回归关系。x1、x2的一次项系数均为正数,且土壤水分(x1)系数绝对值小于植株密度(x2)系数绝对值,表明在一定范围内土壤水分增加传播距离的效应要小于植株密度增加传播距离的效应。x1、x2的二次项系数均为负数,表明该面为一个开口向下的抛物面(图1B),说明垂穗披碱草传播距离在土壤水分和植株密度互作时理论上存在最佳组合,为W3R1。

图1 土壤水分和植物密度互作下垂穗披碱草克隆小株数和传播距离曲面图Fig. 1 3D response surface of ramets per plant and spreading length of E. nutans between soil moisture and plant density
2. 2 地下芽数量
土壤水分、植株密度及其二者互作均显著影响垂穗披碱草地下芽数量(P<0. 05)。随土壤水分增加,地下芽数量呈增加趋势,但随植株密度增加,地下芽数量呈减小趋势(表2)。

表2 土壤水分和植株密度互作对垂穗披碱草地下芽数量的影响Table 2 Effects of soil moisture and plant density on underground buds per plant
地下芽数量F(x1,x2)与土壤水分(x1)、植株密度(x2)的关系为F(x1,x2)=6. 747−0. 0403x1−1. 97x2+0. 00074x12+0. 2479x22−0. 0038x1x2,F 检验时,P=0. 028,R2=0. 573,说明土壤水分、植株密度与地下芽数量具有显著回归关系。x1、x2的一次项系数均为负值,且土壤水分(x1)系数绝对值小于植株密度(x2)系数绝对值,表明在一定范围内土壤水分减少地下芽数量的效应要小于植株密度减少地下芽数量的效应,x1、x2的二次项系数均为正数,表明该面为一个开口向上的抛物面(图2),说明垂穗披碱草地下芽数量在土壤水分和植株密度互作时存在最小值,为W2R3,且理论上的最佳组合为W3R1。

图2 土壤水分和植株密度互作下垂穗披碱草地下芽数量曲面图Fig. 2 3D response surface of underground buds per plant of E. nutans between soil moisture and plant density
2. 3 生物量
土壤水分、植株密度及二者互作对垂穗披碱草地上生物量、地下生物量及总生物量均有显著影响(P<0. 05)。随土壤水分增加,地上生物量、地下生物量及总生物量均呈增加趋势;随植株密度增加,地上生物量、地下生物量及总生物量均呈减小趋势(表3)。

表3 土壤水分和植株密度互作对垂穗披碱草生物量的影响Table 3 Effects of soil moisture and plant density on biomass per plant of E. nutans(g·plant-1)
地上生物量F(x1,x2)与土壤水分(x1)、植株密度(x2)的关系为F(x1,x2)=0. 3316−0. 02579x1−0. 01215x2+0. 00004x12+0. 004583x22−0. 002819x1x2,F 检验时,P=0. 00001,R2=0. 912,说明土壤水分、植株密度与地上生物量具有显著回归关系。x1、x2的一次项系数均为负值,且土壤水分(x1)系数绝对值大于植株密度(x2)系数绝对值,表明在一定范围内,土壤水分减产地上生物量的效应要大于植株密度减产的效应,x1、x2的二次项系数均为正数,表明该面为一个开口向上的抛物面(图3),说明垂穗披碱草地上生物量在土壤水分和植株密度互作时存在最小值,为W1R3,理论上最佳组合为W3R1。

图3 土壤水分和植株密度互作下垂穗披碱草地上生物量曲面图Fig. 3 3D response surface of aboveground biomass of E. nutans between soil moisture and plant density
地下生物量F(x1,x2)与土壤水分(x1)、植株密度(x2)的关系为F(x1,x2)=−0. 6377+0. 05087x1−0. 08432x2−0. 00033x12+0. 0068x22−0. 0011x1x2,F 检验时,P=0. 043,R2=0. 585,说明土壤水分、植株密度与地下生物量具有显著回归关系。x1的一次项系数为正值,x2的一次项系数为负值,且土壤水分(x1)系数绝对值小于植株密度(x2)系数绝对值,表明在一定范围内,土壤水分增产地下生物量的效应要小于植株密度减产的效应,x1的二次项系数为负数,x2的二次项系数为正数,表明垂穗披碱草地下生物量受土壤水分和植株密度互作影响时较为复杂(图4)。互作时存在最小值,为W1R1,理论上的最佳组合为W2R1。

图4 土壤水分和植株密度互作下垂穗披碱草地下生物量曲面图Fig. 4 3D response surface of belowground biomass of E. nutans between soil moisture and plant density
总生物量F(x1,x2)与土壤水分(x1)、植株密度(x2)的关系为F(x1,x2)=−0. 39+0. 06889x1−0. 03875x2−0. 00022x12+0. 008437x22−0. 003938x1x2,F 检验时,P=0. 00001,R2=0. 861,说明土壤水分、植株密度与总生物量具有显著回归关系。x1的一次项系数为正值,x2的一次项系数为负值,且土壤水分(x1)系数绝对值大于植株密度(x2)系数绝对值,表明在一定范围内,土壤含水量增产总生物量的效应要大于植株密度减产的效应,x1的二次项系数为负值,x2的二次项系数为正数,说明垂穗披碱草总生物量受土壤水分和植株密度互作影响时较为复杂(图5)。互作时存在最小值,为W1R3,理论上的最佳组合为W3R1。

图5 土壤水分和植株密度互作下垂穗披碱草总生物量曲面图Fig. 5 3D response surface of total biomass of E. nutans between soil moisture and plant density
2. 4 克隆繁殖和芽库繁殖与生物量间的关系
皮尔森相关分析结果表明,地上生物量、地下生物量与地下芽数量、克隆小株数和传播距离均呈显著正相关(表4),说明地下营养繁殖能力与垂穗披碱草栽培草地生产力具有密切关系,一定程度上说明垂穗披碱草地下营养繁殖能力提高,不仅有助于增加垂穗披碱草的产量,还有助于增加垂穗披碱草的生产力。

表4 各因素的相关系数Table 4 The correlation coefficient of parameters
3 讨论
垂穗披碱草作为优良禾本科牧草[1],其种群自我更新能力事关高寒地区栽培草地的持续生产[12]。本研究采用盆栽试验从克隆繁殖和芽库繁殖两个方面,研究了不同土壤水分和植株密度互作下垂穗披碱草的克隆小株数,传播距离,地下芽数量,生物量的变化特征,以及克隆繁殖和芽库繁殖与生物量之间的关系,解析垂穗披碱草栽培草地适应青藏高原地区水分环境变化和种植密度变化的可能途径,为垂穗披碱草栽培草地稳定持续生产提供理论依据。研究结果表明,垂穗披碱草克隆小株数随土壤水分增加呈先增加后减小的趋势,这与苔草属植物[23]和禾本科植物玉米(Zea mays)[24]的克隆小株数响应土壤水分的过程趋同,说明土壤水分较低或过高,均不利于植物克隆小株数的形成,只有土壤水分适宜时才能促进植物形成更多克隆小株数,从而通过增加垂穗披碱草克隆繁殖能力而很好地适应环境。垂穗披碱草传播距离随土壤水分增加而增加,这不同于美丽箬竹(Indocalamus decorus)克隆分株与土壤水分的关系[25],但类似于淫羊藿(Epimedium brevicornum)传播距离与土壤水分的关系[14],这主要与植物自身的生物学习性有关。若植物属于湿生植物,其传播距离受土壤水分的影响较小,但植物属于中生或旱生植物时,其传播距离受土壤水分影响较大,以帮助植物在更大空间内获取更多资源[23,26−27]。研究结果表明,垂穗披碱草地下芽数量随土壤水分增加而增加,这与Aarssen[28]提到的地下芽数量或密度响应土壤水分的变化趋势一致,主要是土壤水分能够调控地下芽的萌发。植物克隆小株数和地下芽是一个互作系统[29],当土壤水分适宜时,地下芽则会萌发为克隆小株,此时克隆小株数增加,而地下芽数量或密度维持在较低水平,而当土壤水分过大时,此时地下芽萌发为克隆小株的速度受抑,导致克隆小株数数量降低,而地下芽数量或密度维持在较高水平,因此垂穗披碱草克隆小株和地下芽数量对土壤含水量的响应本质是植物自我调节的机制之一。垂穗披碱草地上和地下生物量随土壤水分增加呈增加趋势,这与藜属植物在智利地区响应水分条件的结果趋同[30],当土壤水分能够满足植物生长需要时,则会增加植物地上和地下生物量[31],然而本研究亦发现垂穗披碱草地上和地下生物量增加时所需的土壤水分存在一定的差异,植物地上生物量增加所需的土壤水分要大于植物地下生物量增加所需的土壤水分标准,主要是植物根系首先获取土壤水分,满足自身生长需求,然后才通过自我调节机制向地上部分输送水分。当土壤水分条件满足植物生长需要的时候,垂穗披碱草通过增加克隆小株数而增加地上生物量。水分能改变禾本科植物根茎形态特征,土壤水分增加会增加垂穗披碱草根茎体积和数量[5],从而增加垂穗披碱草地下生物量,而地下生物量的增加,会促进垂穗披碱草更好地利用土壤养分资源和水分资源[25],反哺地上生物量[32]。地上地下生物量随土壤水分增加,促进了垂穗披碱草总生物量的增加。
密度制约是自然植物种群普遍存在的特征,通过改变种群内每个植株可获得资源总量,即植物种群内部个体特征在很大程度上依赖于密度变化状况。本研究发现,垂穗披碱草克隆小株数随植株密度增加呈减小趋势,这与Makihiko 等[33]采用方格模型(lattice model)模拟的结果一致。植株密度增加,每株植物的克隆小株数之间的竞争亦会增加,这会反过来抑制克隆小株数的增加[34]。因此植物密度和克隆小株数的变化过程实质是一个权衡效应过程,主要目的是维持栽培草地的产量相对稳定。垂穗披碱草传播距离随植株密度增加而减小,这与Wong 等[35]采用随机个体模型(stochastic individual-based model)模拟的结果一致,植株密度较高时,资源匮乏,较短的传播距离有助于抵抗这种情况下的外界干扰。垂穗披碱草地下芽数量随植株密度增加呈减小趋势,地下芽是地上部分生长的补偿[36],密度较低时植株会产生更多克隆小株,并且地下芽数量或密度维持在一个较高水平;而密度较高时,资源相对受限,植物增加克隆小株及地下芽数量或密度的能力受抑[37],从而导致地上生物量、地下生物量、总生物量均呈减小趋势,这亦与老芒麦(Elymus sibiricus)生物量对密度的响应结果一致[38]。密度较低的生境内,植物个体能够充分利用环境资源,个体之间的竞争较弱或不存在;但当种群密度较大时,个体间维持正常生长,必然会形成资源竞争格局[16],且随植株密度增加,个体间在竞争光照和养分资源方面的程度变强,光合作用减弱,从而地上生物量降低[39],至密度较高时,就会引起植物个体积累碳水化合物的能力下降,个体生长受阻[16],地下生物量亦减少。个体繁殖能力是栽培草地生产力的基础,然而个体繁殖能力往往与植株密度有关,植物密度维持相对稳定时,则意味着栽培草地能够持续生产[40]。个体繁殖能力因植物密度自疏而发生改变,这种改变若处于一定的密度阈值之内,则会维持栽培草地的持续生产,然而当密度过小时,则因栽培草地不能充分利用自然资源[16−17],而导致垂穗披碱草总生物量减小。
垂穗披碱草克隆小株数、传播距离、地下芽数量与其地上和地下生物量呈显著正相关关系,说明土壤水分和植株密度对垂穗披碱草生物量的增加具有积极效应,但同步存在明显的互作效应,并不是土壤水分越大,植株密度越大,其克隆小株数、传播距离、地下芽数量、地上生物量、地下生物量以及总生物量越大,而是具有最佳的理论组合,本研究设置的9 个处理中,克隆小株数、传播距离、地下芽数量、地上生物量、地下生物量、总生物量对应的最佳组合分别为W2R1、W3R1、W3R1、W3R1、W2R1、W3R1。因此,垂穗披碱草栽培草地的管理,并不是种植密度越大越好,亦不是土壤水分含量越高越好,而是需要根据垂穗披碱草栽培草地的管理目标,采用合理的种植密度,维持合理的土壤水分水平,这为青藏高原地区垂穗披碱草栽培草地的稳定持续生产提供了理论基础。
猜你喜欢
草业学报(2022年11期)2022-11-23
美文(2022年18期)2022-10-20
现代畜牧科技(2021年7期)2021-07-28
西藏农业科技(2021年4期)2021-04-18
临床检验杂志(电子版)(2020年1期)2020-04-03
新农民(2019年9期)2019-02-19
高原山地气象研究(2016年2期)2016-11-10
学苑创造·B版(2015年12期)2016-06-23
塔里木大学学报(2014年3期)2014-03-11
自然资源遥感(2014年2期)2014-02-27
